Digestion Performance and Microbial Metabolic Mechanism in Thermophilic and Mesophilic Anaerobic Digesters Exposed to Elevated Loadings of Organic Fraction of Municipal Solid Waste

Abstract
:1. Introduction
2. Results and Discussion
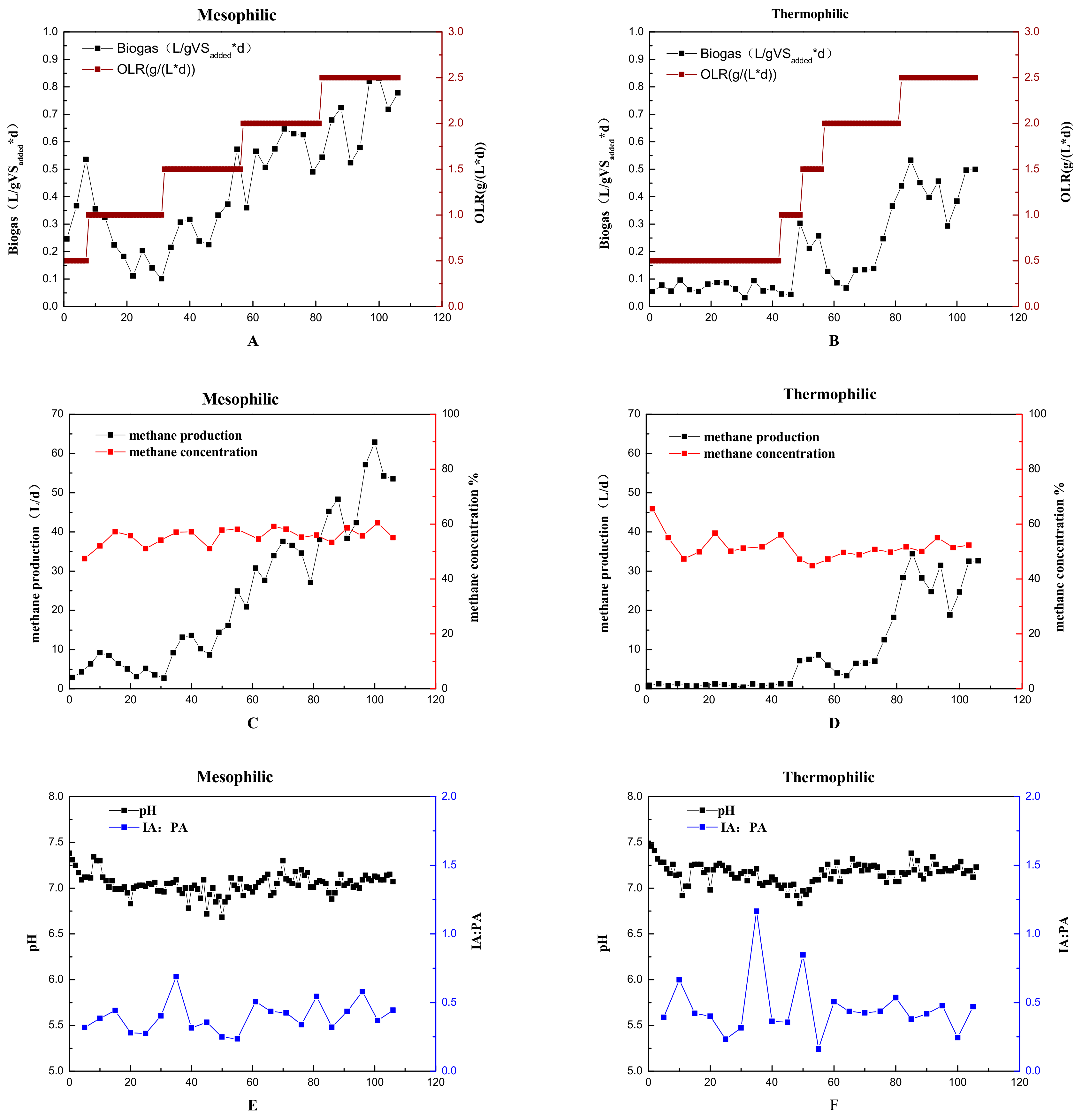
2.1. Performance of the Reactors
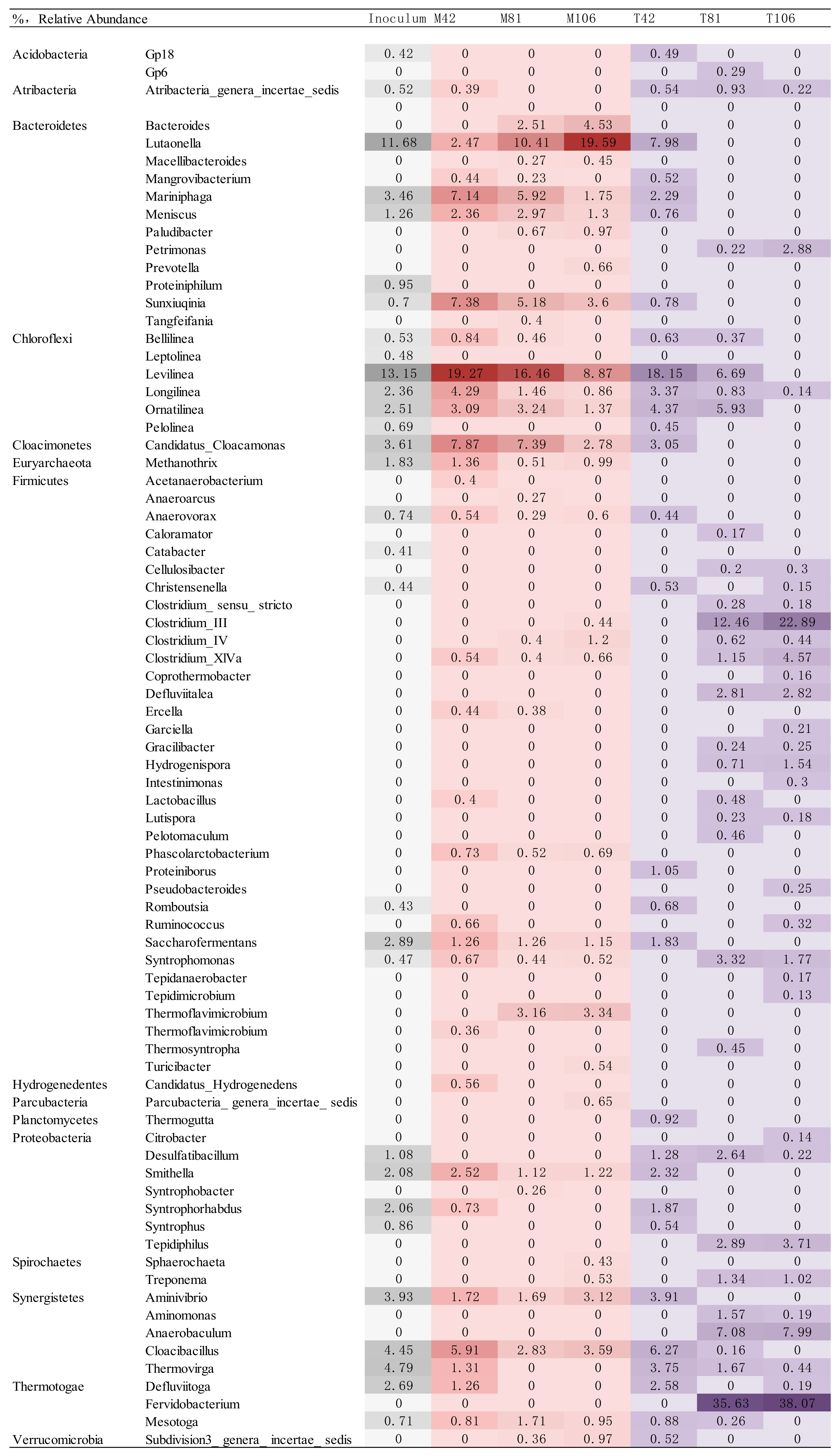
2.2. Bacterial Communities Revealed by Pyrosequencing
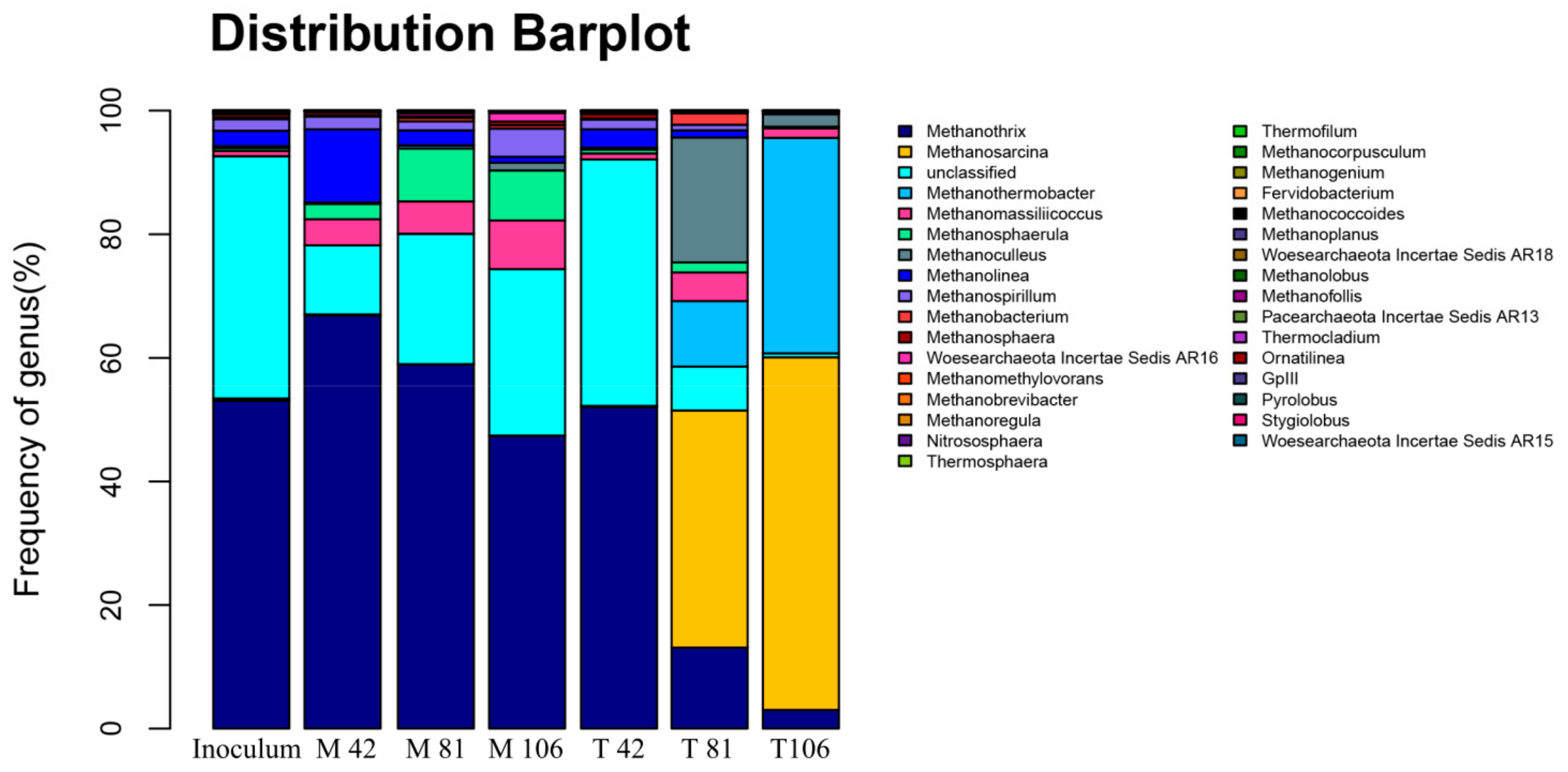
2.3. Dynamics of Methanogen Communities
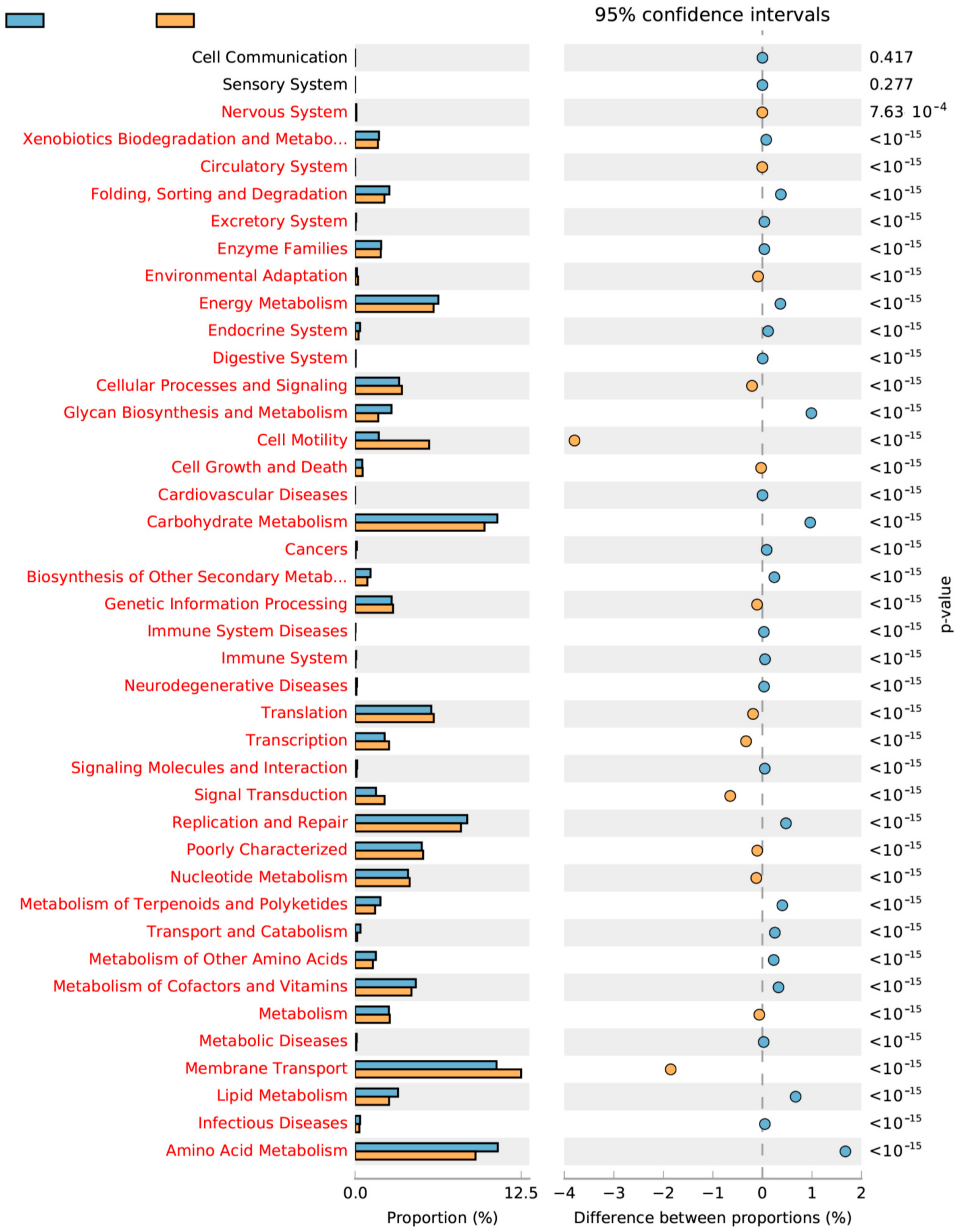
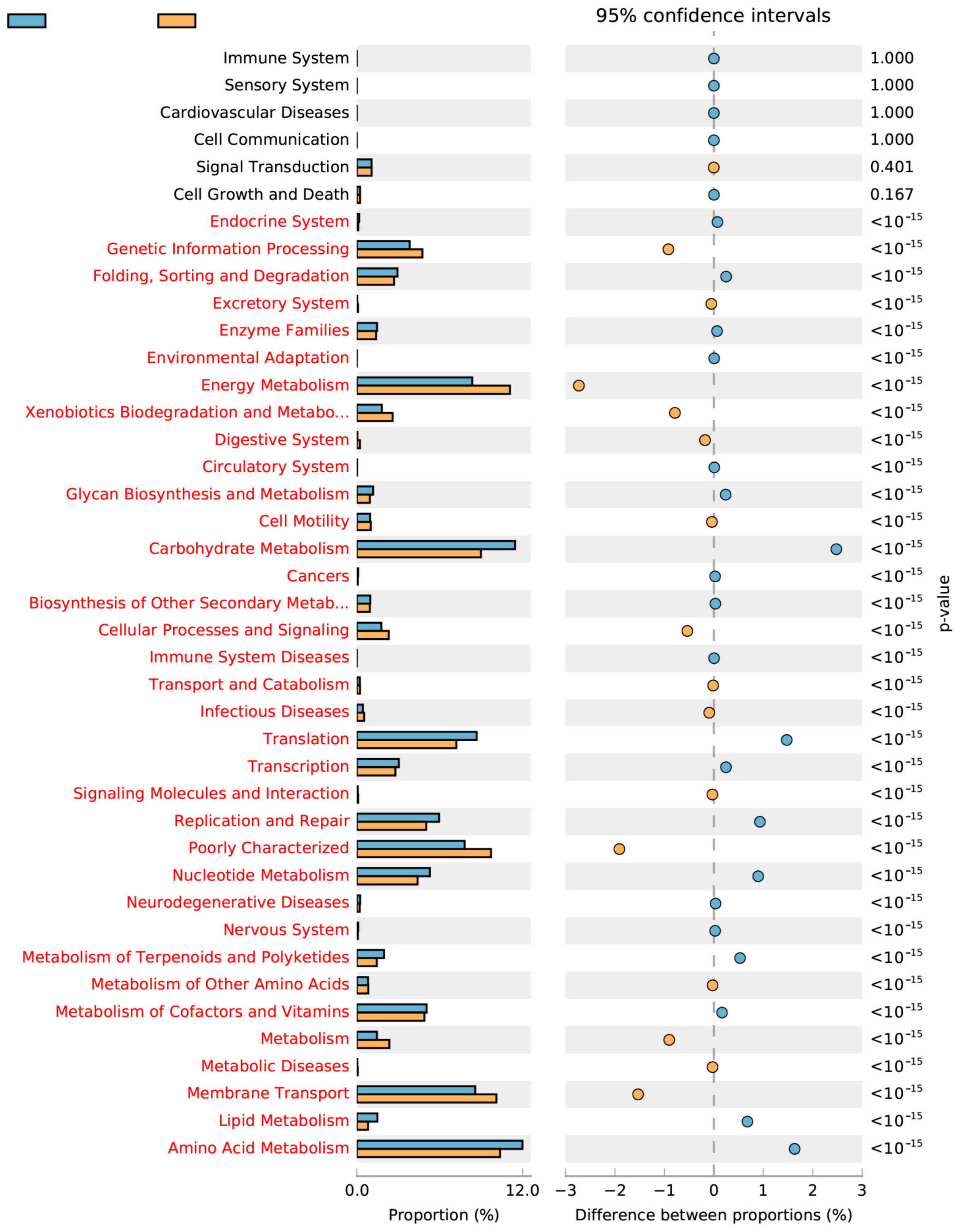
2.4. Linkage between Metabolism and Reactor Performances
3. Materials and Methods
3.1. Anaerobic Digesters
3.2. Chemical Analysis
3.3. DNA Extraction and Amplification
3.4. High-Throughput Pyrosequencing Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Murphy, J.D.; McKeogh, E. Technical, economic and environmental analysis of energy production from municipal solid waste. Renew. Energy 2004, 29, 1043–1057. [Google Scholar] [CrossRef]
- Griffin, M.E.; McMahon, K.D.; Mackie, R.I.; Raskin, L. Methanogenic population dynamics during start-up of anaerobic digesters treating municipal solid waste and biosolids. Biotechnol. Bioeng. 1998, 57, 342–355. [Google Scholar] [CrossRef]
- Song, Y.C.; Kwon, S.J.; Woo, J.H. Mesophilic and thermophilic temperature co-phase anaerobic digestion compared with single-stage mesophilic- and thermophilic digestion of sewage sludge. Water Res. 2004, 38, 1653–1662. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Sun, Y.; Li, L.; Kong, X.; Yuan, Z. Improving methane production from anaerobic digestion of Pennisetum Hybrid by alkaline pretreatment. Bioresour. Technol. 2018, 255, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Panepinto, D.; Genon, G. Analysis of the extrusion as a pretreatment for the anaerobic digestion process. Ind. Crop. Prod. 2016, 83, 206–212. [Google Scholar] [CrossRef]
- Ahring, B.K. Perspectives for anaerobic digestion. In Biomethanation I; Ahring, B.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2003; Volume 81, pp. 1–30. [Google Scholar]
- Bayr, S.; Rantanen, M.; Kaparaju, P.; Rintala, J. Mesophilic and thermophilic anaerobic co-digestion of rendering plant and slaughterhouse wastes. Bioresour. Technol. 2012, 104, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Ahn, Y.H.; Speece, R.E. Comparative process stability and efficiency of anaerobic digestion; mesophilic vs. thermophilic. Water Res. 2002, 36, 4369–4385. [Google Scholar] [CrossRef]
- Ma, Q.; Shen, F.; Yuan, H.R.; Zou, D.X.; Liu, Y.P.; Zhu, B.N.; Li, X.J. Investigation on anaerobic digestion of the organic fraction of municipal solid waste (OFMSW) after pretreatment of fast aerobic fermentation. Fresenius Environ. Bull. 2015, 24, 1039–1046. [Google Scholar]
- Muha, I.; Zielonka, S.; Lemmer, A.; Schonberg, M.; Linke, B.; Grillo, A.; Wittum, G. Do two-phase biogas plants separate anaerobic digestion phases?—A mathematical model for the distribution of anaerobic digestion phases among reactor stages. Bioresour. Technol. 2013, 132, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Gujer, W.; Zehnder, A.J.B. Conversion processes in anaerobic-digestion. Water Sci. Technol. 1983, 15, 127–167. [Google Scholar]
- Schievano, A.; D’Imporzano, G.; Malagutti, L.; Fragali, E.; Ruboni, G.; Adani, F. Evaluating inhibition conditions in high-solids anaerobic digestion of organic fraction of municipal solid waste. Bioresour. Technol. 2010, 101, 5728–5732. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.C.; Morrison, M.; Yu, Z.T. A meta-analysis of the microbial diversity observed in anaerobic digesters. Bioresour. Technol. 2011, 102, 3730–3739. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhang, T. Bacterial communities in different sections of a municipal wastewater treatment plant revealed by 16S rDNA 454 pyrosequencing. Appl. Microbiol. Biotechnol. 2013, 97, 2681–2690. [Google Scholar] [CrossRef] [PubMed]
- Riviere, D.; Desvignes, V.; Pelletier, E.; Chaussonnerie, S.; Guermazi, S.; Weissenbach, J.; Li, T.; Camacho, P.; Sghir, A. Towards the definition of a core of microorganisms involved in anaerobic digestion of sludge. ISME J. 2009, 3, 700–714. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.H.; Wang, C.; Sun, F.Q.; Zhu, W.J.; Wu, W.X. A comparison of microbial characteristics between the thermophilic and mesophilic anaerobic digesters exposed to elevated food waste loadings. Bioresour. Technol. 2014, 152, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Oleszkiewicz, J.A.; Marstaller, T.; McCartney, D.M. Effects of ph on sulfide toxicity to anaerobic processes. Environ. Technol. Lett. 1989, 10, 815–822. [Google Scholar] [CrossRef]
- Ripley, L.E.; Boyle, W.C.; Converse, J.C. Improved alkalimetric monitoring for anaerobic-digestion of high-strength wastes. J. Water Pollut. Control Fed. 1986, 58, 406–411. [Google Scholar]
- Poggivaraldo, H.M.; Tingley, J.; Oleszkiewicz, J.A. Inhibition of growth and acetate uptake by ammonia in batch anaerobic-digestion. J. Chem. Technol. Biotechnol. 1991, 52, 135–143. [Google Scholar] [CrossRef]
- Hansen, K.H.; Angelidaki, I.; Ahring, B.K. Anaerobic digestion of swine manure: Inhibition by ammonia. Water Res. 1998, 32, 5–12. [Google Scholar] [CrossRef]
- Yamada, T.; Sekiguchi, Y.; Hanada, S.; Imachi, H.; Ohashi, A.; Harada, H.; Kamagata, Y. Anaerolinea thermolimosa sp. nov., Levilinea saccharolytica gen. nov., sp. nov and Leptolinea tardivitalis gen. nov., so. nov., novel filamentous anaerobes, and description of the new classes anaerolineae classis nov and Caldilineae classis nov in the bacterial phylum Chloroflexi. Int. J. Syst. Evol. Microbiol. 2006, 56, 1331–1340. [Google Scholar] [PubMed]
- Yamada, T.; Imachi, H.; Ohashi, A.; Harada, H.; Hanada, S.; Kamagata, Y.; Sekiguchi, Y. Bellilinea caldifistulae gen. nov., sp. nov and Longilinea arvoryzae gen. nov., sp. nov., strictly anaerobic, filamentous bacteria of the phylum Chloroflexi isolated from methanogenic propionate-degrading consortia. Int. J. Syst. Evol. Microbiol. 2007, 57, 2299–2306. [Google Scholar] [CrossRef] [PubMed]
- Arun, A.B.; Chen, W.M.; Lai, W.A.; Chou, J.H.; Shen, F.T.; Rekha, P.D.; Young, C.C. Lutaonella thermophila gen. nov., sp. nov., a moderately thermophilic member of the family Flavobacteriaceae isolated from a coastal hot spring. Int. J. Syst. Evol. Microbiol. 2009, 59, 2069–2073. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.Q.; Shen, Q.Y.; Chen, G.J.; Du, Z.J. Mariniphaga sediminis sp. nov., isolated from coastal sediment. Int. J. Syst. Evol. Microbiol. 2015, 65, 2908–2912. [Google Scholar] [CrossRef] [PubMed]
- Irgens, R.L. Meniscus, a new genus of aerotolerant, gas-vacuolated bacteria. Int. J. Syst. Bacteriol. 1977, 27, 38–43. [Google Scholar] [CrossRef]
- Patel, B.; Morgan, H. A Combination of Stable Isotope Probing, Illumina Sequencing, and Co-occurrence Network to Investigate Thermophilic Acetate- and Lactate-Utilizing Bacteria. Microb. Ecol. 2018, 75, 113–122. [Google Scholar]
- Zhang, Y.; Alam, M.A.; Kong, X.; Wang, Z.; Li, L.; Sun, Y.; Yuan, Z. Effect of salinity on the microbial community and performance on anaerobic digestion of marine macroalgae. J. Chem. Technol. Biotechnol. 2017, 92, 2392–2399. [Google Scholar] [CrossRef]
- Demirel, B.; Scherer, P. The roles of acetotrophic and hydrogenotrophic methanogens during anaerobic conversion of biomass to methane: A review. Rev. Environ. Sci. Biotechnol. 2008, 7, 173–190. [Google Scholar] [CrossRef]
- Kendall, M.M.; Boone, D.R. The Order Methanosarcinales. In The Prokaryotes: Volume 3: Archaea. Bacteria: Firmicutes, Actinomycetes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 244–256. [Google Scholar]
- Garcia, J.L.; Patel, B.K.C.; Ollivier, B. Taxonomic phylogenetic and ecological diversity of methanogenic Archaea. Anaerobe 2000, 6, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.-L.; Ollivier, B.; Whitman, W.B. The Order Methanomicrobiales. In The Prokaryotes: Volume 3: Archaea. Bacteria: Firmicutes, Actinomycetes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 208–230. [Google Scholar]
- Zhang, J.X.; Mao, L.W.; Zhang, L.; Loh, K.C.; Dai, Y.J.; Tong, Y.W. Metagenomic insight into the microbial networks and metabolic mechanism in anaerobic digesters for food waste by incorporating activated carbon. Sci. Rep. 2017, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Cleceri, L.S.; Greenberg, A.E.; Eaton, A.D. Standard Methods for the Examination of Water and Waste Water; American Public Health Association: Washington, DC, USA, 1998; p. 113. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Seq num | Shannon | ACE | Chao1 |
|---|---|---|---|---|
| Inoculum | 36,968 | 4.83 | 2085.72 | 2003.94 |
| M42 | 50,721 | 4.35 | 1993.82 | 1993.82 |
| M81 | 34,881 | 3.94 | 1482.31 | 1196.27 |
| M106 | 43,495 | 3.87 | 1333.54 | 1290.58 |
| T42 | 48,268 | 4.84 | 2107.76 | 2010.29 |
| T81 | 24,640 | 3.53 | 819.58 | 740.92 |
| T106 | 39,521 | 3.28 | 812.26 | 754.70 |
| Sample ID | Seq num | Shannon | ACE | Chao1 |
|---|---|---|---|---|
| Inoculum | 37,349 | 1.81 | 400.96 | 358.27 |
| M42 | 35,726 | 1.91 | 479.86 | 395.59 |
| M81 | 38,486 | 2.14 | 597.13 | 458.31 |
| M106 | 38,703 | 2.33 | 622.17 | 497.01 |
| T42 | 38,494 | 2.64 | 608.64 | 552.93 |
| T81 | 38,095 | 2.12 | 476.57 | 330.21 |
| T106 | 39,425 | 1.13 | 175.41 | 158 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Kong, X.; Xing, T.; Sun, Y.; Zhang, Y.; Luo, X.; Sun, Y. Digestion Performance and Microbial Metabolic Mechanism in Thermophilic and Mesophilic Anaerobic Digesters Exposed to Elevated Loadings of Organic Fraction of Municipal Solid Waste. Energies 2018, 11, 952. https://doi.org/10.3390/en11040952
Gao Y, Kong X, Xing T, Sun Y, Zhang Y, Luo X, Sun Y. Digestion Performance and Microbial Metabolic Mechanism in Thermophilic and Mesophilic Anaerobic Digesters Exposed to Elevated Loadings of Organic Fraction of Municipal Solid Waste. Energies. 2018; 11(4):952. https://doi.org/10.3390/en11040952
Chicago/Turabian StyleGao, Yiming, Xiaoying Kong, Tao Xing, Yongming Sun, Yi Zhang, Xingjian Luo, and Yong Sun. 2018. "Digestion Performance and Microbial Metabolic Mechanism in Thermophilic and Mesophilic Anaerobic Digesters Exposed to Elevated Loadings of Organic Fraction of Municipal Solid Waste" Energies 11, no. 4: 952. https://doi.org/10.3390/en11040952
APA StyleGao, Y., Kong, X., Xing, T., Sun, Y., Zhang, Y., Luo, X., & Sun, Y. (2018). Digestion Performance and Microbial Metabolic Mechanism in Thermophilic and Mesophilic Anaerobic Digesters Exposed to Elevated Loadings of Organic Fraction of Municipal Solid Waste. Energies, 11(4), 952. https://doi.org/10.3390/en11040952