Evaluation of Scenedesmus rubescens for Lipid Production from Swine Wastewater Blended with Municipal Wastewater


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation, Identification, and Cultivation of Microalgae
2.2. Design of Experiment
2.3. Physioco-Chemical Analysis
2.4. Microbial Community Analysis
3. Results and Discussion
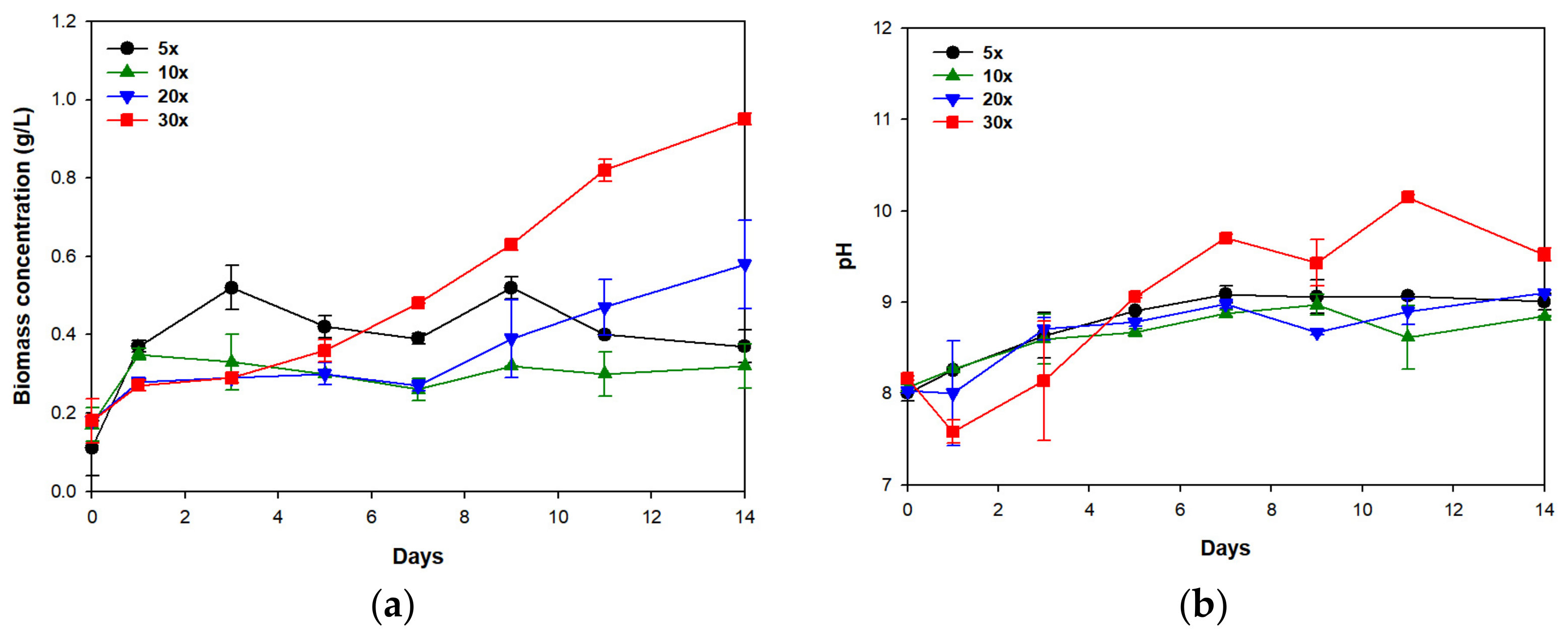
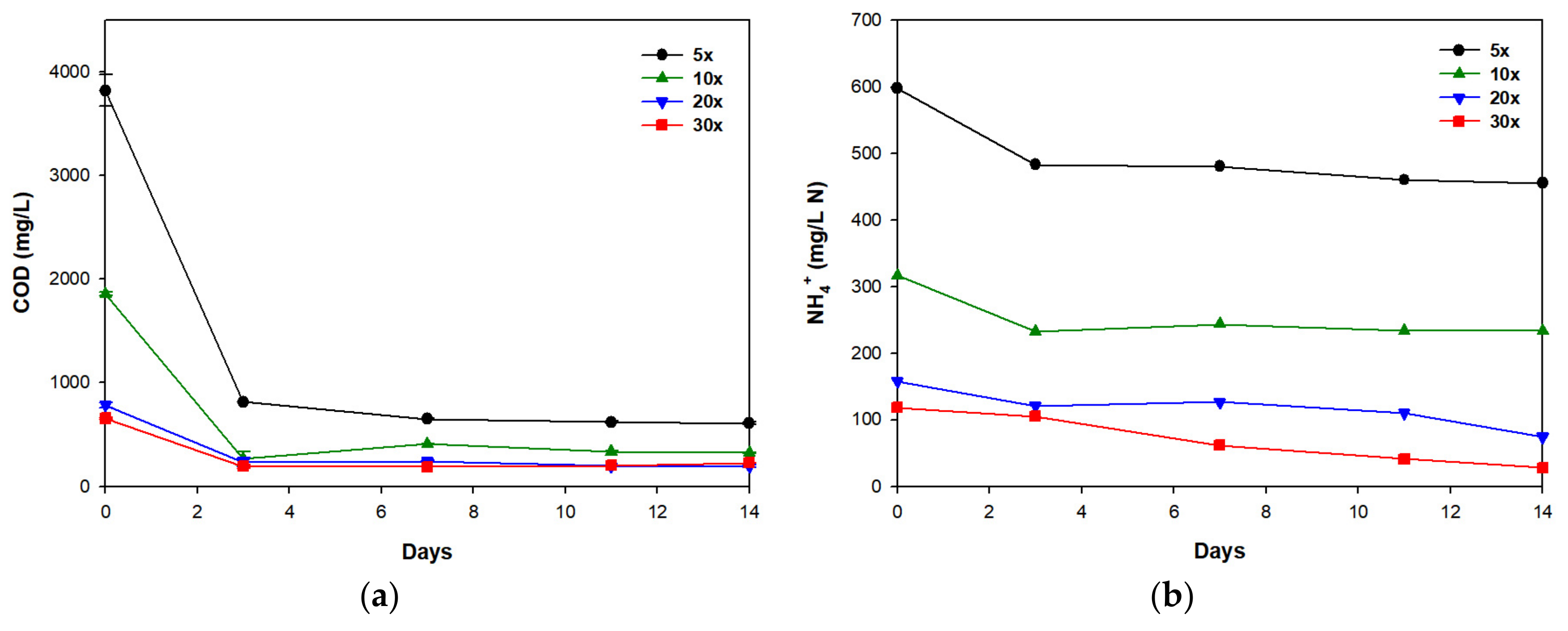
3.1. Biomass Production and Nutrient Removal
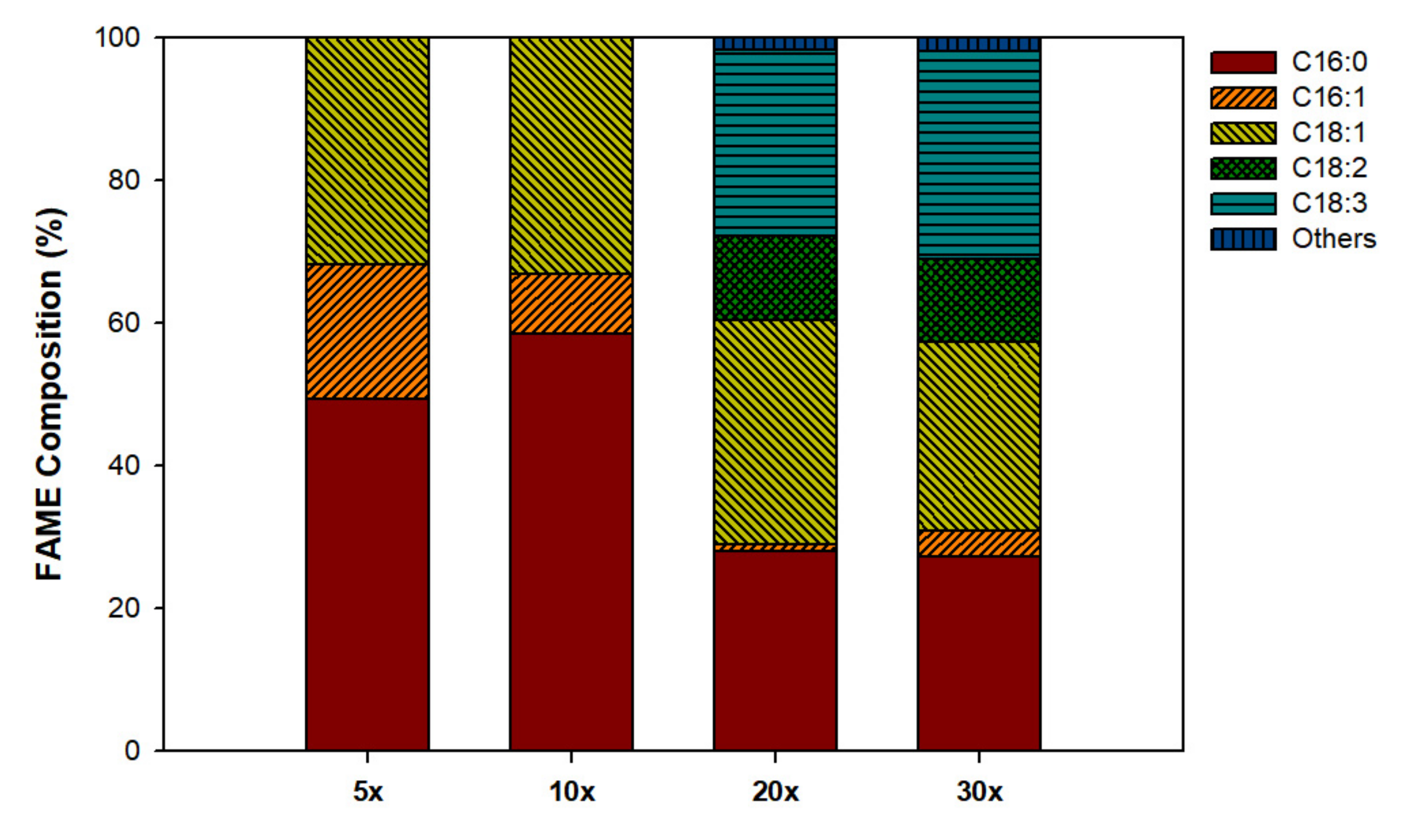
3.2. Lipid and FAMEs Profile
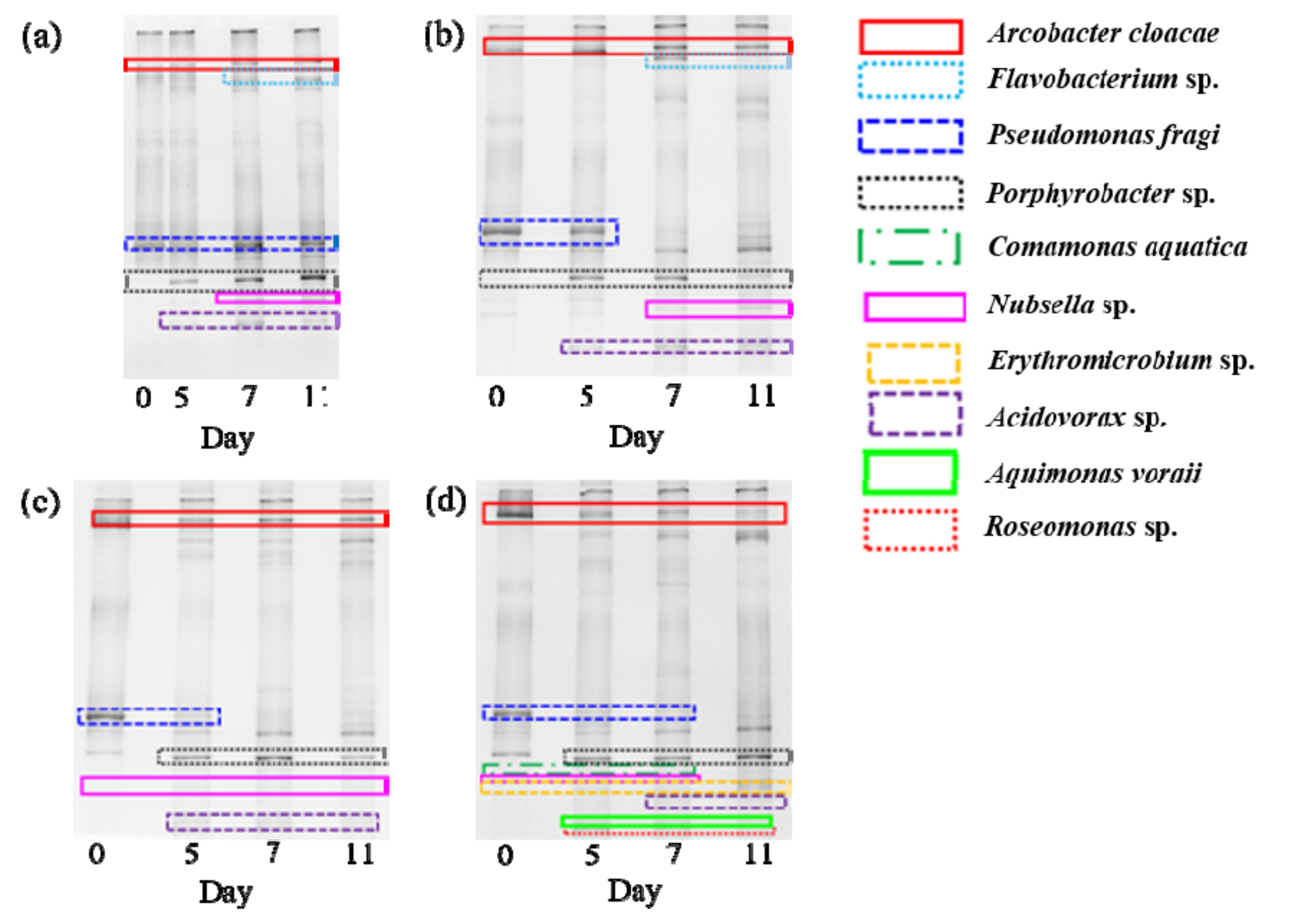
3.3. Microbial Community Structure
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- An, M.; Gao, L.; Zhao, W.; Chen, W.; Li, M. Effects of nitrogen forms and supply mode on lipid production of microalga Scenedesmus obliquus. Energies 2020, 13, 697. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.G.; Hong, C.-H.; Choi, Y.-E. Efficient algal biodiesel production with an optimal harvest condition obtained via response surface methodology. Bioresour. Technol. Rep. 2019, 7, 100213. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Ling, T.C.; Show, P.L.; Juan, J.C.; Chang, J.-S.; Lee, D.-J. Cultivation in wastewaters for energy: A microalgae platform. Appl. Energy 2016, 179, 609–625. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Fu, D.; Chu, H.; Wang, Z.; Qi, H. Biofuel production from microalgae: A review. Environ. Chem. Lett. 2020, 18, 285–297. [Google Scholar] [CrossRef]
- Bodenes, P.; Wang, H.-Y.; Lee, T.-H.; Chen, H.-Y.; Wang, C.-Y. Microfluidic techniques for enhancing biofuel and biorefinery industry based on microalgae. Biotechnol. Biofuels 2019, 12, 33. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.-Y.; Roh, K.; Han, J.-I. The use of bicarbonate for microalgae cultivation and its carbon footprint analysis. Green Chem. 2019, 21, 5053–5062. [Google Scholar] [CrossRef]
- Yang, J.; Xu, M.; Zhang, X.; Hu, Q.; Sommerfeld, M.; Chen, Y. Life-cycle analysis on biodiesel production from microalgae: Water footprint and nutrients balance. Bioresour. Technol. 2011, 102, 159–165. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, L.; Riddicka, B.A.; Li, R.; Able, J.R.; Boakye-Boaten, N.A.; Shahbazi, A. Sustainable production of algal biomass and biofuels using swine wastewater in North Carolina, US. Sustainability 2016, 8, 477. [Google Scholar] [CrossRef]
- Kwon, G.; Nam, J.-H.; Kim, D.-M.; Song, C.; Jahng, D. Growth and nutrient removal of Chlorella vulgaris in ammonia-reduced raw and anaerobically-digested piggery wastewaters. Environ. Eng. Res. 2020, 25, 135–146. [Google Scholar] [CrossRef]
- Zou, G.; Liu, Y.; Zhang, Q.; Zhou, T.; Xiang, S.; Gu, Z.; Huang, Q.; Yan, H.; Zheng, H.; Wu, X.; et al. Cultivation of Chlorella vulgaris in a light-receiving-plate (LRP)-enhanced raceway pond for ammonium and phosphorus removal from pretreated pig urine. Energies 2020, 13, 1644. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.-M.; Chen, T.-Y.; Lin, T.-H.; Kao, C.-Y.; Lai, J.-T.; Chang, J.-S.; Lin, C.-S. Cultivation of Chlorella sp. GD using piggery wastewater for biomass and lipid production. Bioresour. Technol. 2015, 194, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Erol, H.B.U.; Menegazzo, M.L.; Sanderfur, H.; Gottberg, E.; Vaden, J.; Asgharpour, M.; Hestekin, C.N.; Hestekin, J.A. Porphyridium cruentum grown in ultra-filtered swine wastewater and its effects on microalgae growth productivity and fatty acid composition. Energies 2020, 13, 3194. [Google Scholar] [CrossRef]
- Franchino, M.; Tigini, V.; Varese, G.C.; Sartor, R.M.; Bona, F. Microalgae treatment removes nutrients and reduces ecotoxicity of diluted piggery digestate. Sci. Total Environ. 2016, 569, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Cheng, J.J.; Cobb, K.; Zhou, C.; Zhou, N.; Addy, M.; Chen, P.; Yan, X.; Ruan, R. Tribonema sp. and Chlorella zofingiensis co-culture to treat swine wastewater diluted with fishery wastewater to facilitate harvest. Bioresour. Technol. 2020, 297, 122516. [Google Scholar] [CrossRef] [PubMed]
- Eland, L.E.; Davenport, R.; Mota, C.R. Evaluation of DNA extraction methods for freshwater eukaryotic microalgae. Water Res. 2012, 46, 5355–5364. [Google Scholar] [CrossRef]
- Lv, J.; Guo, B.; Feng, J.; Liu, Q.; Nan, F.; Liu, X.; Xie, S. Integration of wastewater treatment and flocculation for harvesting biomass for lipid production by a newly isolated self-flocculating microalga Scenedesmus rubescens SX. J. Clean Prod. 2019, 240, 118211. [Google Scholar] [CrossRef]
- Abinandan, S.; Shanthankumar, S. Challenges and opportunities in application of microalgae (Chlorophyta) for wastewater treatment: A review. Renew. Sustain. Energy Rev. 2015, 52, 123–132. [Google Scholar] [CrossRef]
- Clesceri, L.S.; Greenberg, A.E.; Eaton, A.D. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Bourque, S.D.; Titorenko, V.I. A quantitative assessment of the yeast lipidome using electrospray ionization mass spectrometry. J. Vis. Exp. 2009, 30, 1513. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Kim, J.Y.; Cho, H.U.; Park, H.R.; Park, J.M. Bioconversion of volatile fatty acids from macroalgae fermentation into microbial lipids by oleaginous yeast. Chem. Eng. J. 2015, 264, 735–743. [Google Scholar] [CrossRef]
- Pinzi, S.; Leiva, D.; Arzamendi, G.; Gandia, L.M.; Dorado, M.P. Multiple response optimization of vegetable oils fatty acid composition to improve biodiesel physical properties. Bioreour. Technol. 2011, 102, 7280–7288. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Hwang, K.; Shin, S.G.; Lee, S.; Hwang, S. Effect of high temperature on bacterial community dynamics in anaerobic acidogenesis using mesophilic sludge inoculum. Bioreour. Technol. 2010, 101, S17–S22. [Google Scholar] [CrossRef] [PubMed]
- Hatt, J.W.; Germain, E.; Judd, S.J. Powered activated carbon-microfiltration for waste-water reuse. Sep. Sci. Technol. 2013, 48, 690–698. [Google Scholar] [CrossRef]
- Yeh, K.L.; Chen, C.Y.; Chang, J.S. PH-stat photoheterotrophic cultivation of indigenous Chlorella vulgaris ESP-31 for biomass and lipid production using acetic acid as the carbon source. Biochem. Eng. J. 2012, 64, 1–7. [Google Scholar] [CrossRef]
- Shin, D.Y.; Cho, H.U.; Utomo, J.C.; Choi, Y.N.; Xu, X.; Park, J.M. Biodiesel production from Scenedesmus bijuga grown in anaerobically digested food wastewater effluent. Bioresour. Technol. 2015, 184, 215–221. [Google Scholar] [CrossRef]
- Cho, H.U.; Kim, Y.M.; Park, J.M. Enhanced microalgal biomass and lipid production from a consortium of indigenous microalgae and bacteria present in municipal wastewater under gradually mixotrophic culture conditions. Bioresour. Technol. 2017, 228, 290–297. [Google Scholar] [CrossRef]
- Fenchel, T. Intrinsic rate of natural increase: The relationship with body size. Oecologia 1974, 14, 317–326. [Google Scholar] [CrossRef]
- He, P.J.; Mao, B.; Shen, C.M.; Shao, L.M.; Lee, D.J.; Chang, J.S. Cultivation of Chlorella vulgaris on wastewater containing high levels of ammonia for biodiesel production. Bioresour. Technol. 2013, 129, 177–181. [Google Scholar] [CrossRef]
- Park, J.; Jin, H.-F.; Lim, B.-R.; Park, K.-Y.; Lee, K. Ammonia removal from anaerobic digestion effluent of livestock waste using green alga Scenedesmus sp. Bioresour. Technol. 2010, 101, 8649–8657. [Google Scholar] [CrossRef]
- Cheng, P.; Wang, Y.; Osei-Wusu, D.; Wang, Y.; Liu, T. Development of nitrogen supply strategy for Scenedesmus rubescens attached cultivation toward growth and lipid accumulation. Bioprocess. Biosyst. Eng. 2018, 41, 435–442. [Google Scholar] [CrossRef]
- Ye, S.; Gao, L.; Zhao, J.; An, M.; Wu, H.; Li, M. Simultaneous wastewater treatment and lipid production by Scenedesmus sp. HXY2. Bioreour. Technol. 2020, 302, 122903. [Google Scholar] [CrossRef] [PubMed]
- Knothe, G. Dependence of biodiesel fuel properties on the structure of fatty acid alkyl esters. Fuel Process. Technol. 2005, 86, 1059–1070. [Google Scholar] [CrossRef]
- Suh, H.K.; Lee, C.S. A review on atomization and exhaust emissions of a biodiesel-fueled compression ignition engine. Renew. Sustain. Energy Rev. 2016, 58, 1601–1620. [Google Scholar] [CrossRef]
- Levican, A.; Collado, L.; Figueras, M.J. Arcobacter cloacae sp. nov. and Arcobacter suis sp. nov., two new species isolated from food and sewage. Syst. Appl. Microbiol. 2013, 36, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Heylen, K.; Lebbe, L.; Vos, P.D. Acidovorax caeni sp. nov., a denitrifying species with genetically diverse isolates from activated sludge. Int. J. Syst. Evol. Microbiol. 2008, 58, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Guo, L.; Shao, M.; Hu, F.; Wang, G.; Zhao, Y.; Gao, M.; Jin, C.; She, Z. Heterotrophic denitrification strategy for marine recirculating aquaculture wastewater treatment using mariculture solid wastes fermentation liquid as carbon source: Optimization of COD/NO3− -N ratio and hydraulic retention time. Bioreour. Technol. 2020, 304, 122982. [Google Scholar] [CrossRef]
- Ryu, S.H.; Park, M.; Jeon, Y.; Lee, J.R.; Park, W.; Jeon, C.O. Flavobacterium filum sp. nov., isolated from a wastewater treatment plant in Korea. Int. J. Syst. Evol. Microbiol. 2007, 57, 2026–2030. [Google Scholar] [CrossRef]
- Han, J.; Zhang, L.; Wang, S.; Yang, G.; Zhao, L.; Pan, K. Co-culturing bacteria and microalgae in organic carbon containing medium. J. Biol. Res. 2016, 23, 8. [Google Scholar] [CrossRef] [Green Version]
- Afi, L.; Metzger, P.; Largeau, C.; Connan, J.; Berkaloff, C.; Rousseau, B. Bacterial degradation of green microalgae: Incubation of Chlorella emersonii and Chlorella vulgaris with Pseudomonas oleovorans and Flavobacterium aquatile. Org. Geochem. 1996, 25, 117–130. [Google Scholar] [CrossRef]
- Krohn-Molt, I.; Alawi, M.; Forstner, K.U.; Wiegandt, A.; Burkhardt, L.; Indenbirken, D.; Thieb, M.; Grundhoff, A.; Kehr, J.; Tholey, A.; et al. Insights into microalga and bacteria interactions of selected phycosphere biofilms using metagenomic, transcriptomic, and proteomic approaches. Front. Microbiol. 2017, 8, 1941. [Google Scholar] [CrossRef]
- Xinjie, W.; Xin, N.; Qilu, C.; Ligen, X.; Yuhua, Z.; Qifa, Z. Vetiver and Dictyosphaerium sp. co-culture for the removal of nutrients and ecological inactivation of pathogens in swine wastewater. J. Adv. Res. 2019, 20, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Murinda, S.E.; Murry, M.A.; Schwartz, G.; Lundquist, T. Microbial community structures in high rate algae ponds for bioconversion of agricultural wastes from livestock industry for feed production. Sci. Total Environ. 2017, 580, 1185–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Wang, Y.; Yang, T.; He, Z.; Yan, Q. Size-fractioned aggregates within phycosphere define functional bacterial communities related Microcystis aeruginosa and Euglena sanguinea blooms. Aquat. Ecol. 2020, 54, 609–623. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COD (mg/L) c | TN (mg/L N) c | TP (mg/L PO4−3) c | pH c | UV400nm | |
|---|---|---|---|---|---|
| Swine wastewater | 23,495 ± 960.0 | 3850 ± 70.7 | 127 ± 21.5 | 7.7 ± 0.2 | 3.77 |
| Municipal wastewater | 60.3 ± 5.2 | 15.0 ± 1.4 | 4.2 ± 0.2 | 7.3 ± 0.1 | 0.03 |
| 5× a | 3818.6 + 152.4 b | 597.5 + 74.2 b | 27.8 + 0.4 b | 8.0 ± 0.1 b | - |
| 10× a | 1855.4 + 16.9 b | 320.0 + 22.6 b | 17.2 + 0.5 b | 8.1 ± 0.0 b | - |
| 20× a | 784.1 + 25.4 b | 149.0 + 2.8 b | 9.6 + 0.1 b | 8.0 ± 0.0 b | - |
| 30× a | 655.9 + 6.8 b | 116.0 + 2.8 b | 7.4 + 0.1 b | 8.1 ± 0.0 b | - |
| Identification | Nearest Sequence | Similarity (%) | Accession No. |
|---|---|---|---|
| A | Scenedesmus rubescens | 99 | JQ315585.1 |
| B | Parachlorella kessleri | 97 | KM020114.1 |
| C | Chlorella vulgaris | 99 | AB080307.1 |
| D | Chlorella sorokiniana | 99 | GQ122327 |
| E | Scenedesmus acuminatus | 99 | AB037088 |
| Biomass Productivity (mg/L/d) a | Lipid Productivity (mg/L/d) a | |
|---|---|---|
| 5× | 18.6 + 1.4 | 0.49 + 0.1 |
| 10× | 10.7 + 0.7 | 0.44 + 0.1 |
| 20× | 28.6 + 4.7 | 4.44 + 0.4 |
| 30× | 55.0 + 2.1 | 8.37 + 1.5 |
| COD Removal Efficiency (%) a | NH4+ Removal Efficiency (%) a | |||
|---|---|---|---|---|
| Day 3 | Final | Day 3 | Final | |
| 5× | 78.68 ± 0.62 | 84.09 ± 0.23 | 23.40 ± 5.72 | 28.41 ± 4.78 |
| 10× | 85.80 ± 2.51 | 82.63 ± 0.14 | 26.49 ± 3.32 | 26.22 ± 0.27 |
| 20× | 70.22 ± 3.23 | 75.50 ± 0.52 | 23.21 ± 0.99 | 52.87 ± 1.00 |
| 30× | 70.62 ± 1.44 | 65.62 ± 0.07 | 11.35 ± 1.06 | 76.49 ± 1.32 |
| LCV a (MJ/kg) | CN b | μ c (mm2/s) | FP d (°C) | CEPP e (°C) | |
|---|---|---|---|---|---|
| Standard of biodiesel based on EN14214 | 42.64 f | >51 | 3.5 to 5.0 | >120 | Country-specific |
| 20× | 37.3 | 54.9 | 4.6 | 154.9 | −5.4 |
| 30× | 37.3 | 53.7 | 4.5 | 153.2 | −6.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Utomo, J.C.; Kim, Y.M.; Cho, H.U.; Park, J.M. Evaluation of Scenedesmus rubescens for Lipid Production from Swine Wastewater Blended with Municipal Wastewater. Energies 2020, 13, 4895. https://doi.org/10.3390/en13184895
Utomo JC, Kim YM, Cho HU, Park JM. Evaluation of Scenedesmus rubescens for Lipid Production from Swine Wastewater Blended with Municipal Wastewater. Energies. 2020; 13(18):4895. https://doi.org/10.3390/en13184895
Chicago/Turabian StyleUtomo, Joseph Christian, Young Mo Kim, Hyun Uk Cho, and Jong Moon Park. 2020. "Evaluation of Scenedesmus rubescens for Lipid Production from Swine Wastewater Blended with Municipal Wastewater" Energies 13, no. 18: 4895. https://doi.org/10.3390/en13184895
APA StyleUtomo, J. C., Kim, Y. M., Cho, H. U., & Park, J. M. (2020). Evaluation of Scenedesmus rubescens for Lipid Production from Swine Wastewater Blended with Municipal Wastewater. Energies, 13(18), 4895. https://doi.org/10.3390/en13184895