
Bioconversion of Crude Glycerol into 1,3-Propanediol(1,3-PDO) with Bioelectrochemical System and Zero-Valent Iron Using Klebsiella pneumoniae L17


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Cultivation
2.2. BES Configuration and Operation
2.3. Analytical Methods
2.4. Metabolic Flux Analysis (MFA)
3. Results and Discussion
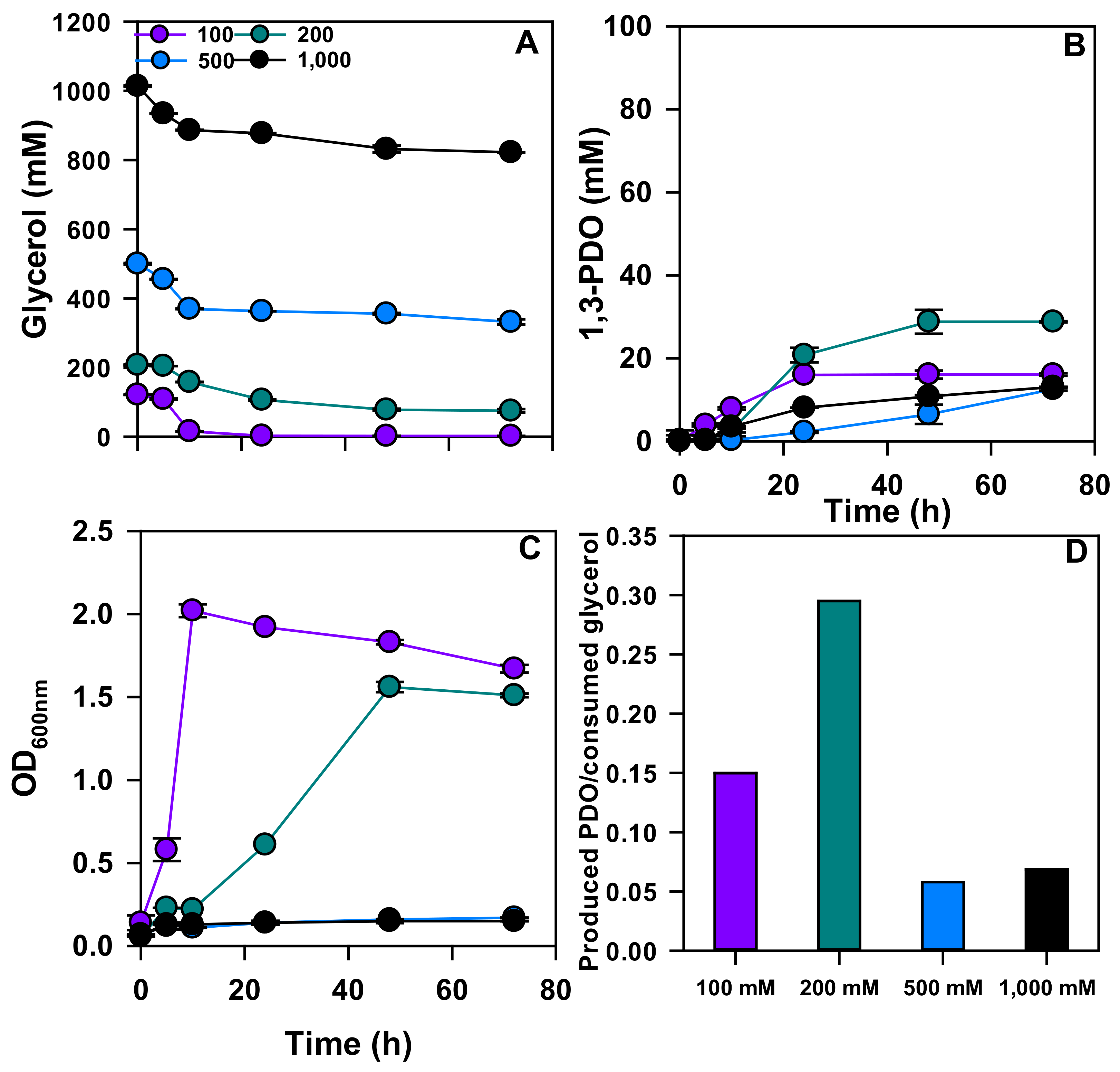
3.1. 1,3-PDO Production in Flask Experiments
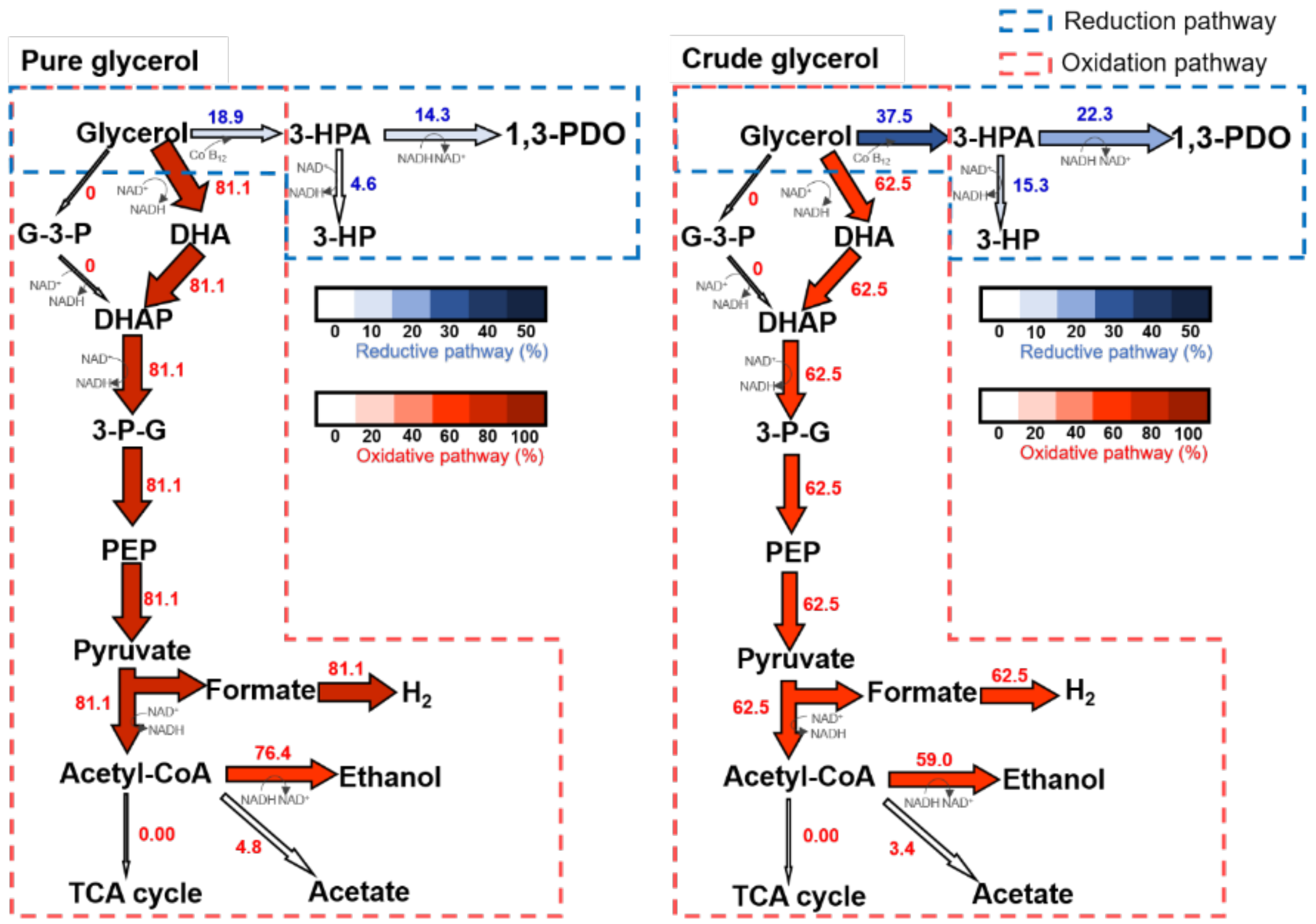
3.2. Flux Balance Analysis (FBA) of Crude and Pure Glycerol
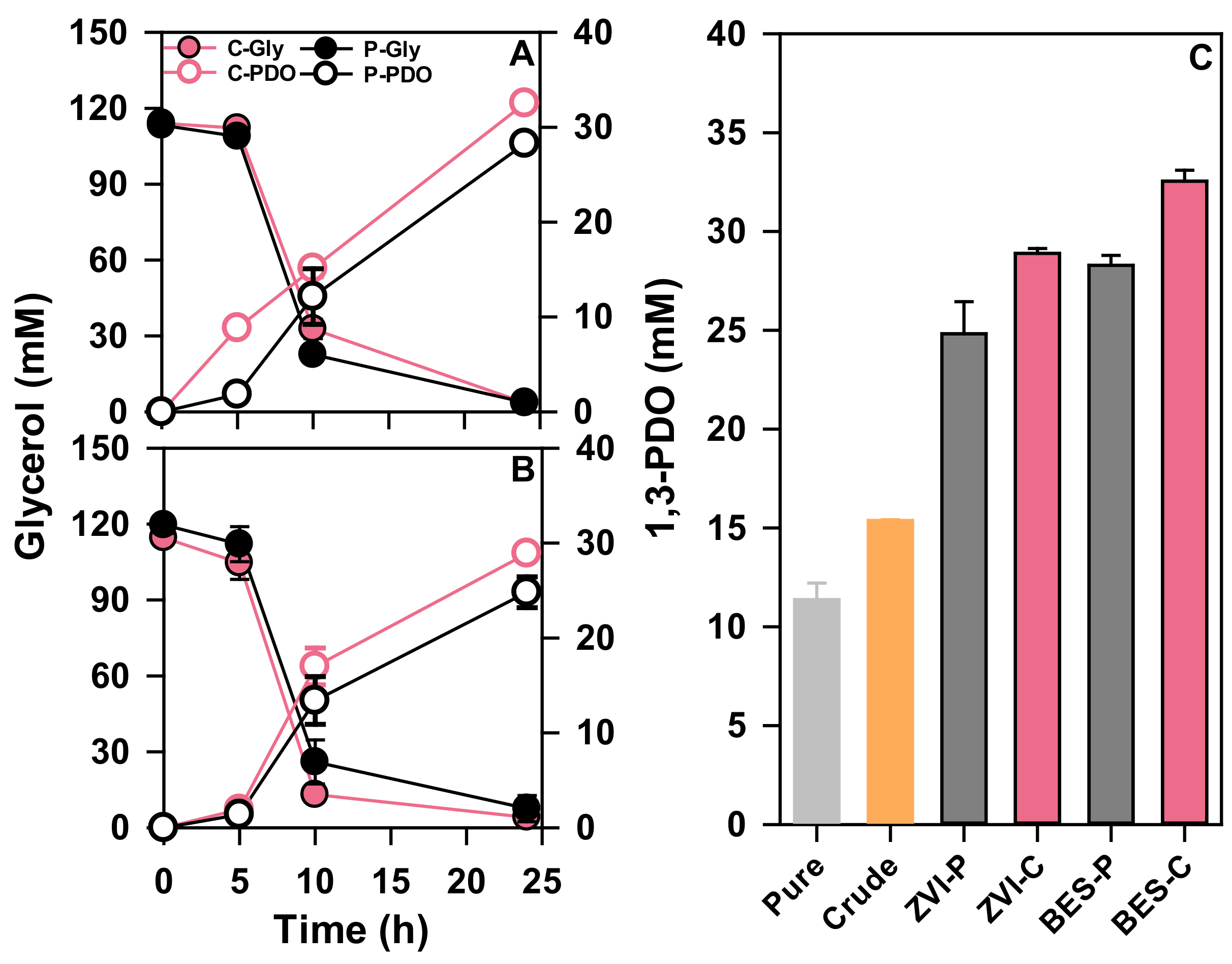
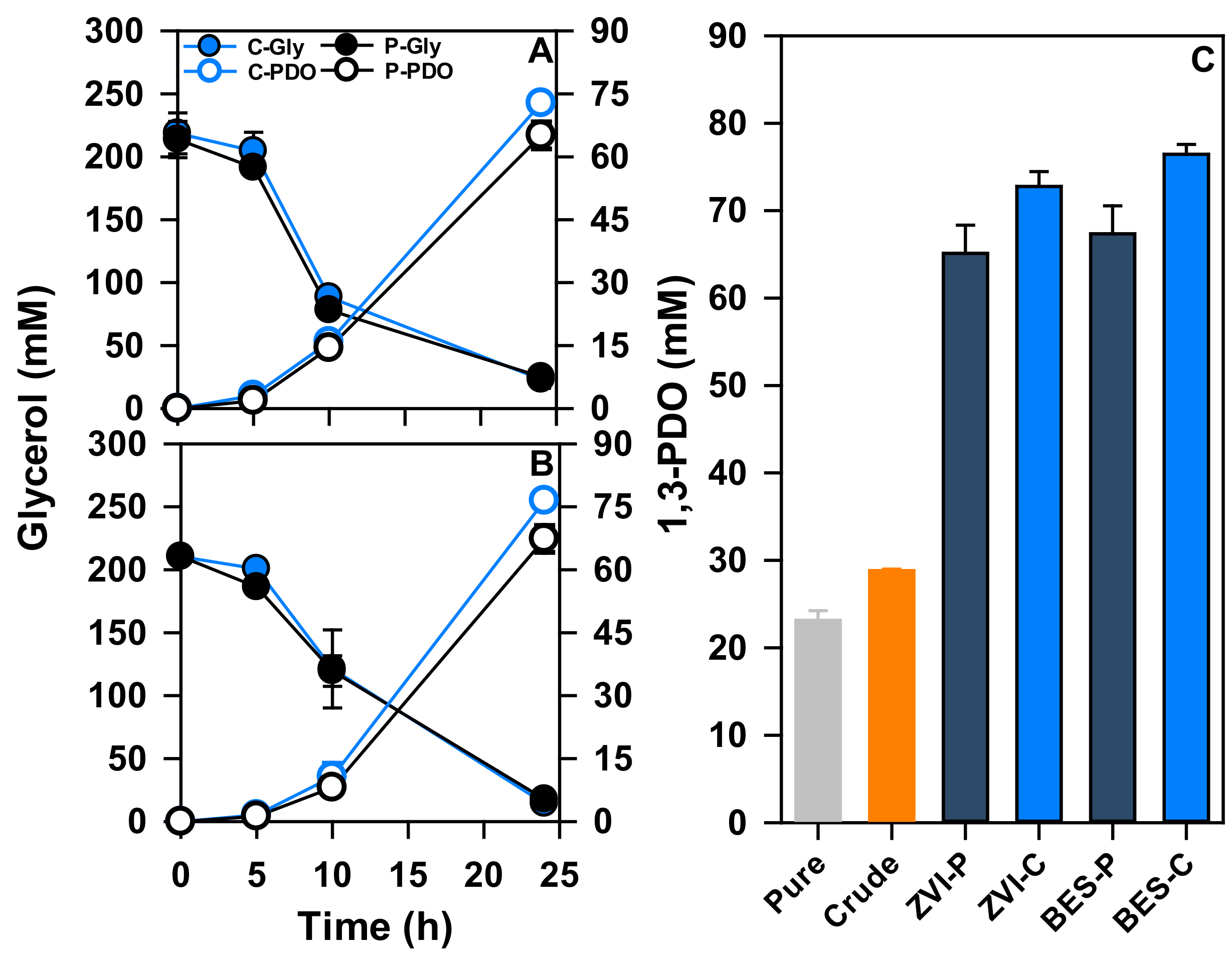
3.3. Electron Uptake for Improvement of 1,3-PDO Production
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dobson, R.; Gray, V.; Rumbold, K. Microbial utilization of crude glycerol for the production of value-added products. J. Ind. Microbiol. Biotechnol. 2012, 39, 217–226. [Google Scholar] [CrossRef]
- Da Silva Ruy, A.D.; Ferreira, A.L.F.; Bresciani, A.É.; de Brito Alves, R.M.; Pontes, L.A.M. Market Prospecting and Assessment of the Economic Potential of Glycerol from Biodiesel. In Biomass; IntechOpen: London, UK, 2020. [Google Scholar]
- Chen, C.Y.; Lee, M.H.; Leong, Y.K.; Chang, J.S.; Lee, D.J. Biodiesel production from heterotrophic oleaginous microalga Thraustochytrium sp. BM2 with enhanced lipid accumulation using crude glycerol as alternative carbon source. Bioresour. Technol. 2020, 306, 123113. [Google Scholar] [CrossRef]
- Talebian-Kiakalaieh, A.; Amin, N.A.S.; Rajaei, K.; Tarighi, S. Oxidation of bio-renewable glycerol to value-added chemicals through catalytic and electro-chemical processes. Appl. Energy 2018, 230, 1347–1379. [Google Scholar] [CrossRef]
- Yang, F.X.; Hanna, M.A.; Sun, R.C. Value-added uses for crude glycerol-a byproduct of biodiesel production. Biotechnol. Biofuels 2012, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrowolski, A.; Drzymala, K.; Mitula, P.; Mironczuk, A.M. Production of tailor-made fatty acids from crude glycerol at low pH by Yarrowia lipolytica. Bioresour.Technol. 2020, 314, 123746. [Google Scholar] [CrossRef] [PubMed]
- Veiga, S.; Faccio, R.; Romero, M.; Bussi, J.J.B. Bioenergy, Utilization of waste crude glycerol for hydrogen production via steam reforming over Ni–La–Zr catalysts. Biomass Bioenergy 2020, 135, 105508. [Google Scholar] [CrossRef]
- Sittijunda, S.; Reungsang, A. Methane Production from the Co-digestion of Algal Biomass with Crude Glycerol by Anaerobic Mixed Cultures. Waste Biomass Valoriz. 2020, 11, 1873–1881. [Google Scholar] [CrossRef]
- Laura, M.; Monica, T.; Dan-Cristian, V. The effect of crude glycerol impurities on 1,3-propanediol biosynthesis by Klebsiella pneumoniae DSMZ 2026. Renew. Energy 2020, 153, 1418–1427. [Google Scholar] [CrossRef]
- Mizielinska, M.; Kowalska, U.; Sobol, M. The continuous bioconversion of glycerol to 1,3-propanediol using immobilized Citrobacter freundii. Rom. Biotech. Lett. 2020, 25, 1448–1455. [Google Scholar] [CrossRef]
- De Oliveira Paranhos, A.G.; Silva, E.L.J.B. Bioenergy, Statistical optimization of H2, 1, 3-propanediol and propionic acid production from crude glycerol using an anaerobic fluidized bed reactor: Interaction effects of substrate concentration and hydraulic retention time. Biomass Bioenergy 2020, 138, 105575. [Google Scholar] [CrossRef]
- Casali, S.; Gungormusler, M.; Bertin, L.; Fava, F.; Azbar, N. Development of a biofilm technology for the production of 1,3-propanediol (1,3-PDO) from crude glycerol. Biochem. Eng. J. 2012, 64, 84–90. [Google Scholar] [CrossRef]
- Kim, C.; Lee, J.H.; Baek, J.; Kong, D.S.; Na, J.G.; Lee, J.; Sundstrom, E.; Park, S.; Kim, J.R. Small Current but Highly Productive Synthesis of 1,3-Propanediol from Glycerol by an Electrode-Driven Metabolic Shift in Klebsiella pneumoniae L17. ChemSusChem 2020, 13, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Aroua, M.K.; Daud, W.M.A.W.; Cognet, P.; Peres-Lucchese, Y.; Fabre, P.L.; Reynes, O.; Latapie, L. A review: Conversion of bioglycerol into 1,3-propanediol via biological and chemical method. Renew. Sustain. Energy Rev. 2015, 42, 963–972. [Google Scholar] [CrossRef] [Green Version]
- Vivek, N.; Pandey, A.; Binod, P. Biological valorization of pure and crude glycerol into 1,3-propanediol using a novel isolate Lactobacillus brevis N1E9.3.3. Bioresour. Technol. 2016, 213, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Gungormusler-Yilmaz, M.; Cicek, N.; Levin, D.B.; Azbar, N. Cell immobilization for microbial production of 1,3-propanediol. Crit. Rev. Biotechnol. 2016, 36, 482–494. [Google Scholar] [CrossRef]
- Kim, C.; Kim, M.Y.; Michie, I.; Jeon, B.H.; Premier, G.C.; Park, S.; Kim, J.R. Anodic electro-fermentation of 3-hydroxypropionic acid from glycerol by recombinant Klebsiella pneumoniae L17 in a bioelectrochemical system. Biotechnol. Biofuels 2017, 10, 199. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Sankaranarayanan, M.; Jae, K.E.; Durgapal, M.; Ashok, S.; Ko, Y.; Sarkar, R.; Park, S. Co-production of 3-hydroxypropionic acid and 1,3-propanediol from glycerol using resting cells of recombinant Klebsiella pneumoniae J2B strain overexpressing aldehyde dehydrogenase. Appl. Microbiol. Biotechnol. 2012, 96, 373–383. [Google Scholar] [CrossRef]
- Li, X.M.; Zhou, S.G.; Li, F.B.; Wu, C.Y.; Zhuang, L.; Xu, W.; Liu, L. Fe(III) oxide reduction and carbon tetrachloride dechlorination by a newly isolated Klebsiella pneumoniae strain L17. J. Appl. Microbiol. 2009, 106, 130–139. [Google Scholar] [CrossRef]
- Liu, T.; Li, X.; Zhang, W.; Hu, M.; Li, F. Fe(III) oxides accelerate microbial nitrate reduction and electricity generation by Klebsiella pneumoniae L17. J. Colloid Interface Sci. 2014, 423, 25–32. [Google Scholar] [CrossRef]
- Kim, M.Y.; Kim, C.; Ainala, S.K.; Bae, H.; Jeon, B.H.; Park, S.; Kim, J.R. Metabolic shift of Klebsiella pneumoniae L17 by electrode-based electron transfer using glycerol in a microbial fuel cell. Bioelectrochemistry 2019, 125, 1–7. [Google Scholar] [CrossRef]
- Kim, C.; Ainala, S.K.; Oh, Y.K.; Jeon, B.H.; Park, S.; Kim, J.R. Metabolic flux change in Klebsiella pneumoniae L17 by anaerobic respiration in microbial fuel cell. Biotechnol. Bioprocess Eng. 2016, 21, 250–260. [Google Scholar] [CrossRef]
- Kong, D.S.; Kim, C.; Song, Y.E.; Baek, J.; Im, H.S.; Kim, J.R. Zero-valent iron driven bioconversion of glycerol to 1, 3-propanediol using Klebsiella pneumoniae L17. J. Process. Biochem. 2021, 106, 158–162. [Google Scholar] [CrossRef]
- Im, H.S.; Kim, C.; Song, Y.E.; Baek, J.; Im, C.H.; Kim, J.R. Isolation of Novel CO Converting Microorganism Using Zero Valent Iron for a Bioelectrochemical System (BES). Biotechnol. Bioprocess Eng. 2019, 24, 232–239. [Google Scholar] [CrossRef]
- Lee, D.Y.; Yun, H.; Park, S.; Lee, S.Y. MetaFluxNet: The management of metabolic reaction information and quantitative metabolic flux analysis. Bioinformatics 2003, 19, 2144–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayoub, M.; Abdullah, A.Z. Critical review on the current scenario and significance of crude glycerol resulting from biodiesel industry towards more sustainable renewable energy industry. Renew. Sustain Energy Rev. 2012, 16, 2671–2686. [Google Scholar] [CrossRef]
- Streekstra, H.; Demattos, M.J.T.; Neijssel, O.M.; Tempest, D.W. Overflow Metabolism during Anaerobic Growth of Klebsiella-Aerogenes Nctc 418 on Glycerol and Dihydroxyacetone in Chemostat Culture. Arch. Microbiol. 1987, 147, 268–275. [Google Scholar] [CrossRef]
- Vemuri, G.N.; Eiteman, M.A.; McEwen, J.E.; Olsson, L.; Nielsen, J. Increasing NADH oxidation reduces overflow metabolism in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2007, 104, 2402–2407. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Mohapatra, S.K.; Ragit, S.S.; Kundu, K.; Karmakar, R. Optimization of safflower oil transesterification using the Taguchi approach. Petrol. Sci. 2017, 14, 798–805. [Google Scholar] [CrossRef] [Green Version]
- Salakkam, A.; Webb, C. The inhibition effect of methanol, as a component of crude glycerol, on the growth rate of Cupriavidus necator and other micro-organisms. Biochem. Eng. J. 2015, 98, 84–90. [Google Scholar] [CrossRef]
- Valerio, O.; Horvath, T.; Pond, C.; Misra, M.; Mohanty, A. Improved utilization of crude glycerol from biodiesel industries: Synthesis and characterization of sustainable biobased polyesters. Ind. Crop. Prod. 2015, 78, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Chen, J.; Freguia, S.; Rabaey, K.; Keller, J. Carbon and electron fluxes during the electricity driven 1,3-propanediol biosynthesis from glycerol. Environ. Sci. Technol. 2013, 47, 11199–11205. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crude Glycerol | Pure Glycerol | 1,3-Propanediol | |
|---|---|---|---|
| Price (USD/ton) | 170 | 1275 | 2000 |
| Type of Glycerol | Initial Glycerol | Glycerol | 1,3-PDO | Produced PDO/ Consumed Glycerol Ratio |
|---|---|---|---|---|
| Concentration | Consumption | Production | ||
| (mM) | (mM) | (mM) | ||
| Pure glycerol | 100 | 110.2 ± 0.1 | 11.4 ± 0.1 | 0.10 |
| 200 | 197.6 ± 1.0 | 23.3 ± 1.1 | 0.14 | |
| 500 | 310.0 ± 3.6 | 49.0 ± 1.7 | 0.16 | |
| 1000 | 235.3 ± 3.8 | 74.1 ± 3.0 | 0.31 | |
| Crude glycerol | 100 | 119.6 ± 0.1 | 16.1 ± 0.3 | 0.15 |
| 200 | 132.9 ± 2.9 | 28.8 ± 0.2 | 0.29 | |
| 500 | 168.2 ± 7.4 | 12.6 ± 0.4 | 0.06 | |
| 1000 | 191.7 ± 2.4 | 13.1 ± 0.1 | 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, D.S.; Park, E.J.; Mutyala, S.; Kim, M.; Cho, Y.; Oh, S.E.; Kim, C.; Kim, J.R. Bioconversion of Crude Glycerol into 1,3-Propanediol(1,3-PDO) with Bioelectrochemical System and Zero-Valent Iron Using Klebsiella pneumoniae L17. Energies 2021, 14, 6806. https://doi.org/10.3390/en14206806
Kong DS, Park EJ, Mutyala S, Kim M, Cho Y, Oh SE, Kim C, Kim JR. Bioconversion of Crude Glycerol into 1,3-Propanediol(1,3-PDO) with Bioelectrochemical System and Zero-Valent Iron Using Klebsiella pneumoniae L17. Energies. 2021; 14(20):6806. https://doi.org/10.3390/en14206806
Chicago/Turabian StyleKong, Da Seul, Eun Joo Park, Sakuntala Mutyala, Minsoo Kim, Yunchul Cho, Sang Eun Oh, Changman Kim, and Jung Rae Kim. 2021. "Bioconversion of Crude Glycerol into 1,3-Propanediol(1,3-PDO) with Bioelectrochemical System and Zero-Valent Iron Using Klebsiella pneumoniae L17" Energies 14, no. 20: 6806. https://doi.org/10.3390/en14206806
APA StyleKong, D. S., Park, E. J., Mutyala, S., Kim, M., Cho, Y., Oh, S. E., Kim, C., & Kim, J. R. (2021). Bioconversion of Crude Glycerol into 1,3-Propanediol(1,3-PDO) with Bioelectrochemical System and Zero-Valent Iron Using Klebsiella pneumoniae L17. Energies, 14(20), 6806. https://doi.org/10.3390/en14206806