Measurement of Lipid Droplet Accumulation Kinetics in Chlamydomonas reinhardtii Using Seoul-Fluor


 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
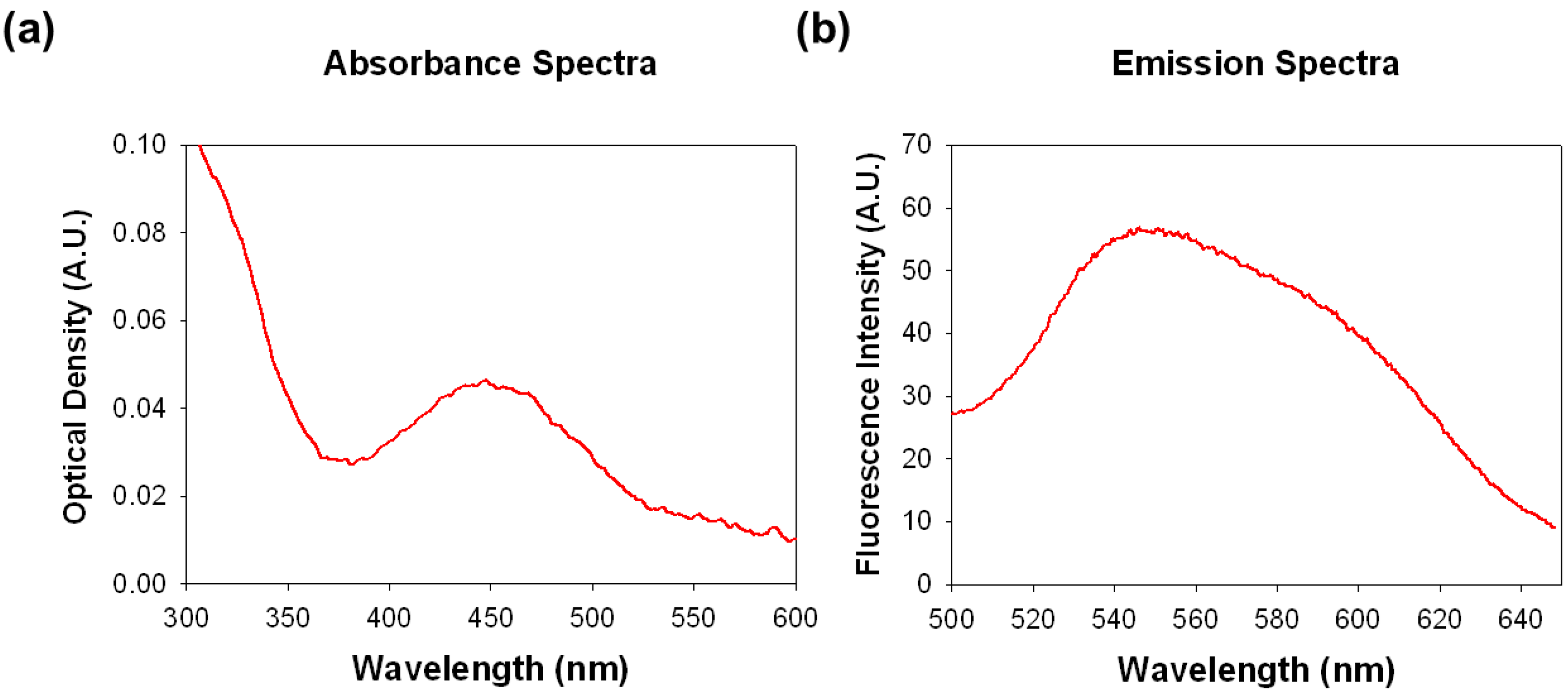
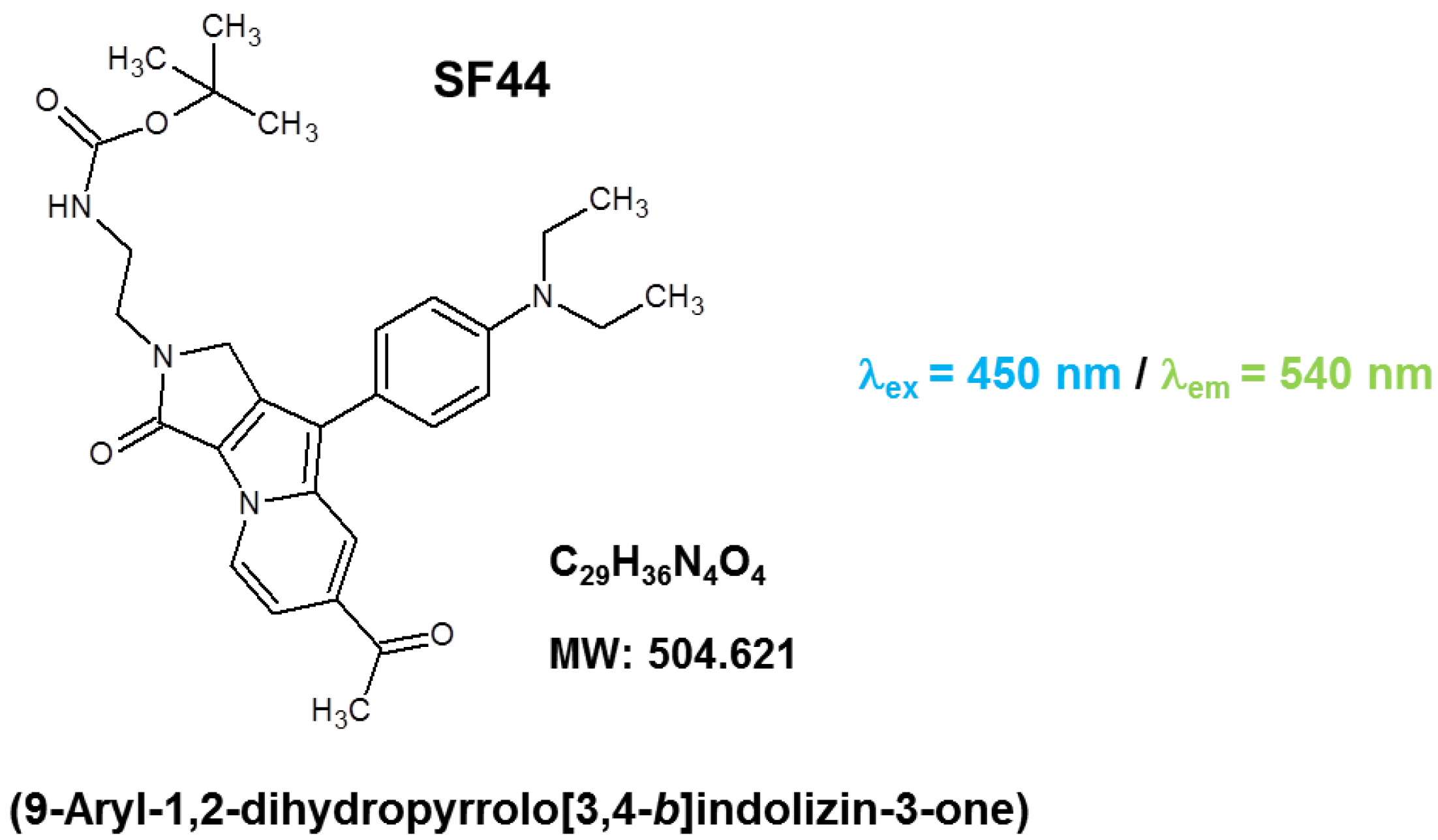
2.1. Optimization of SF44 Staining for Microalgae


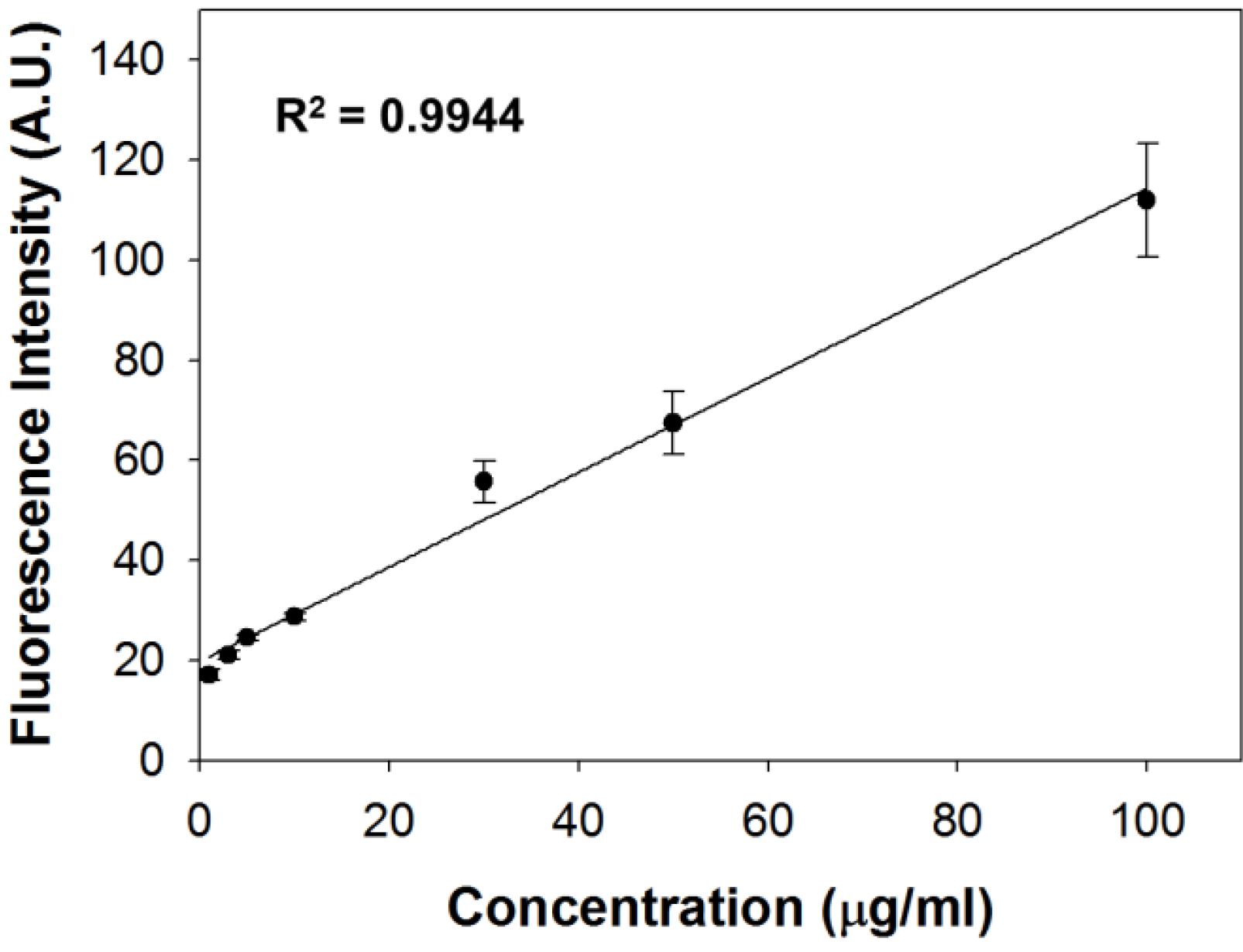
2.2. Quantification of Lipid Contents Using SF44



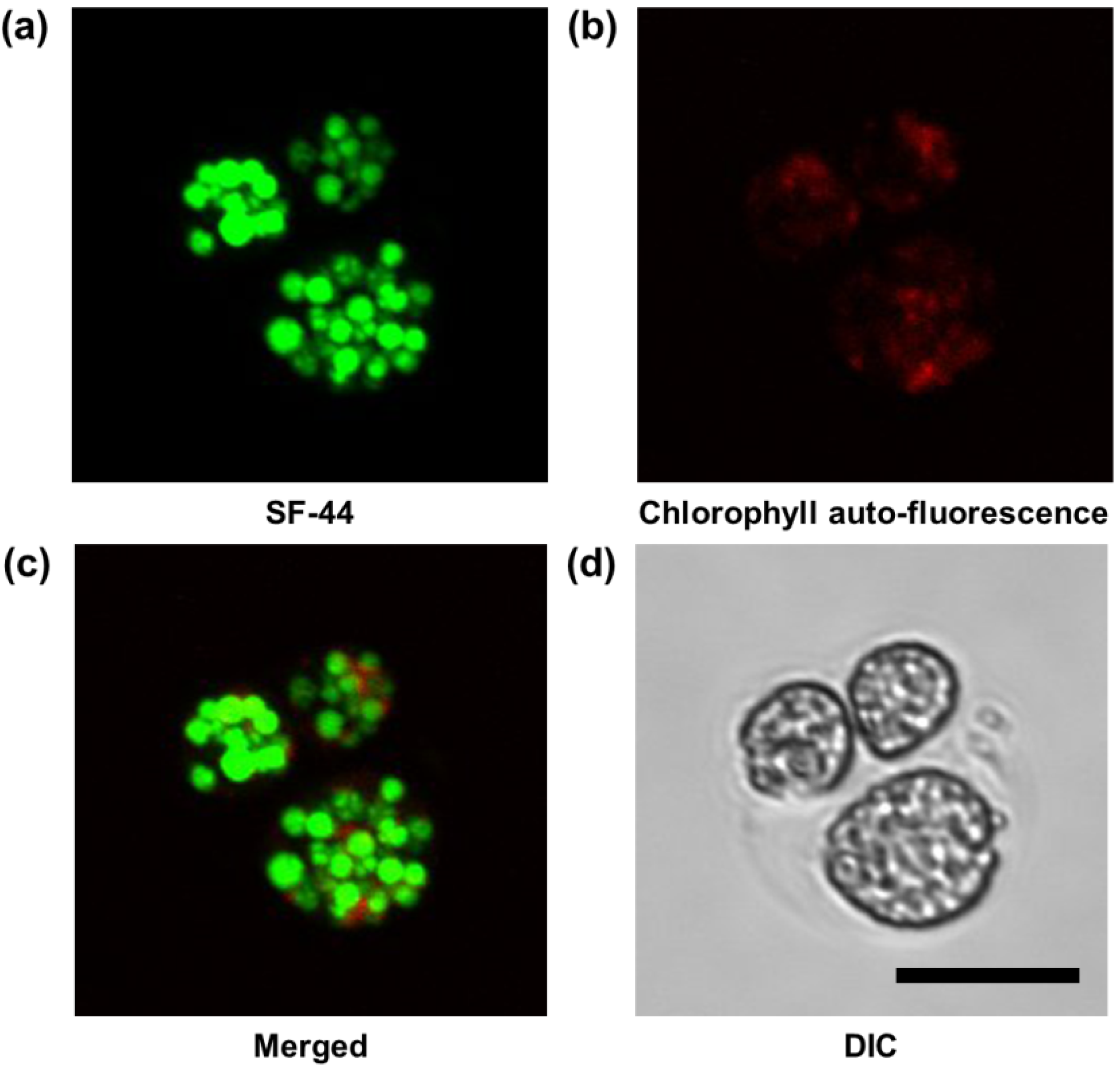
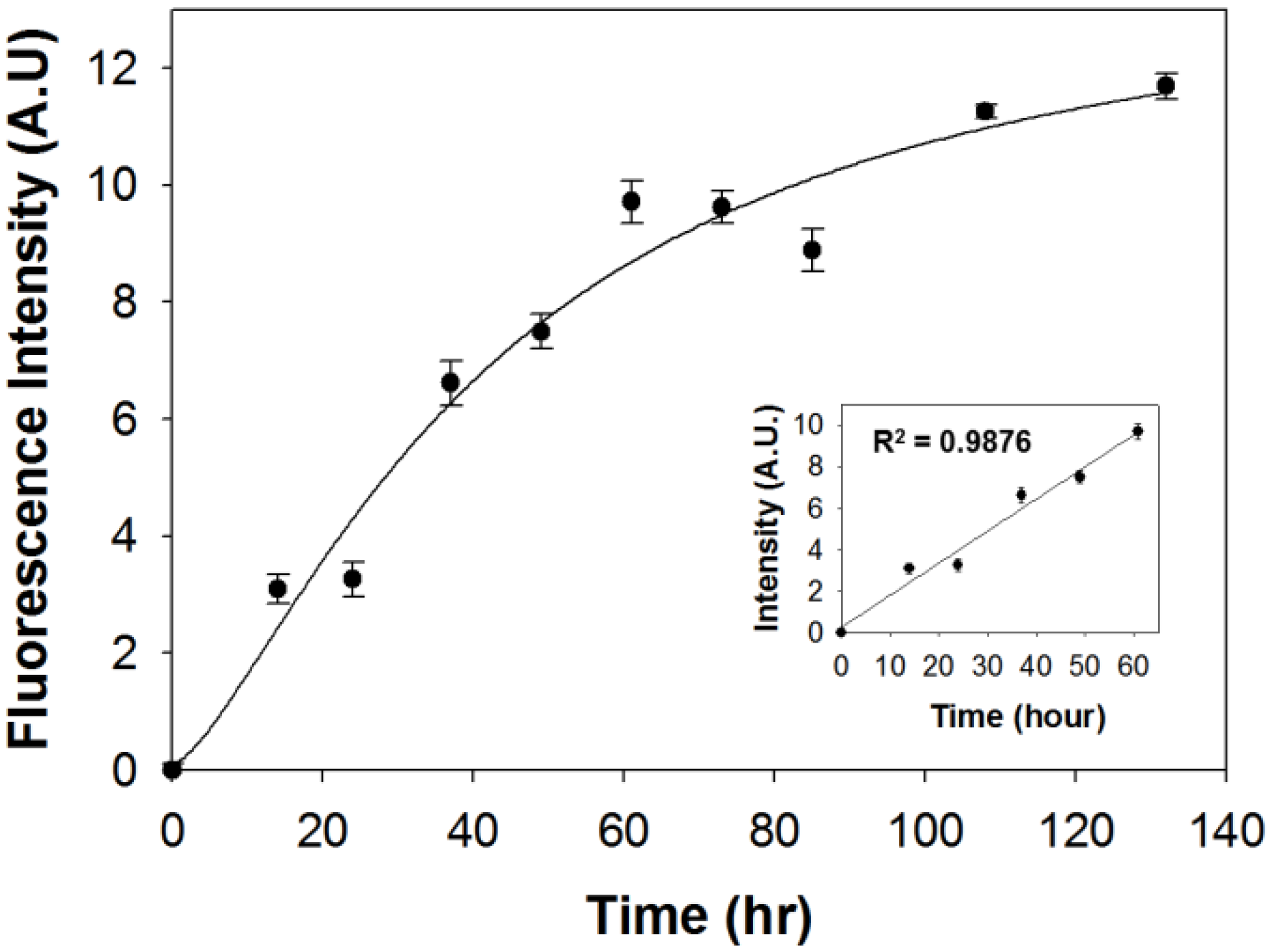
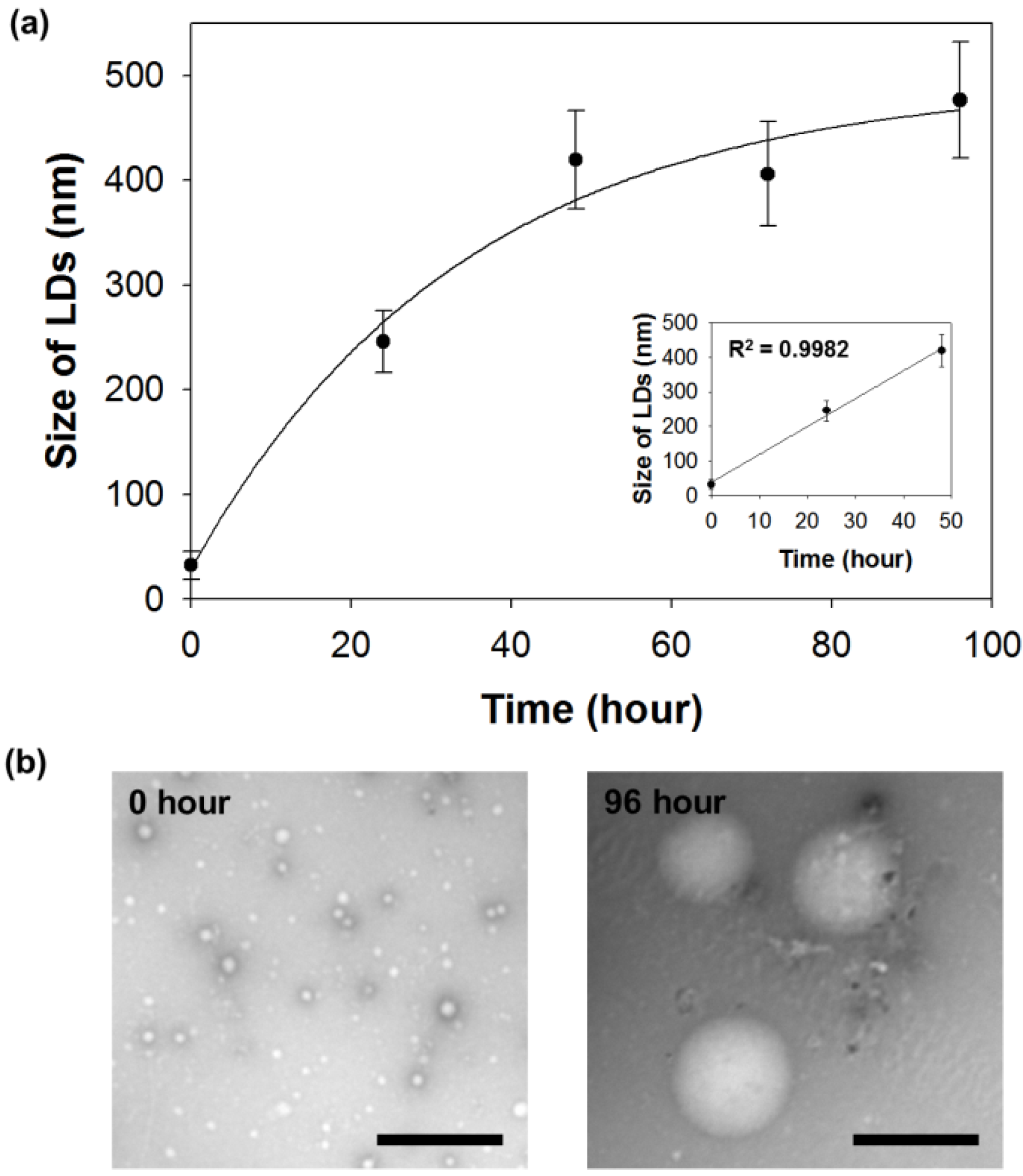
2.3. Measurement of Intracellular LDs Accumulation through SF44



3. Experimental Section
3.1. Material
3.2. Culture Conditions
3.3. Visualization of LDs in C. reinhardtii with SF44
3.4. Optimization of SF44 Staining
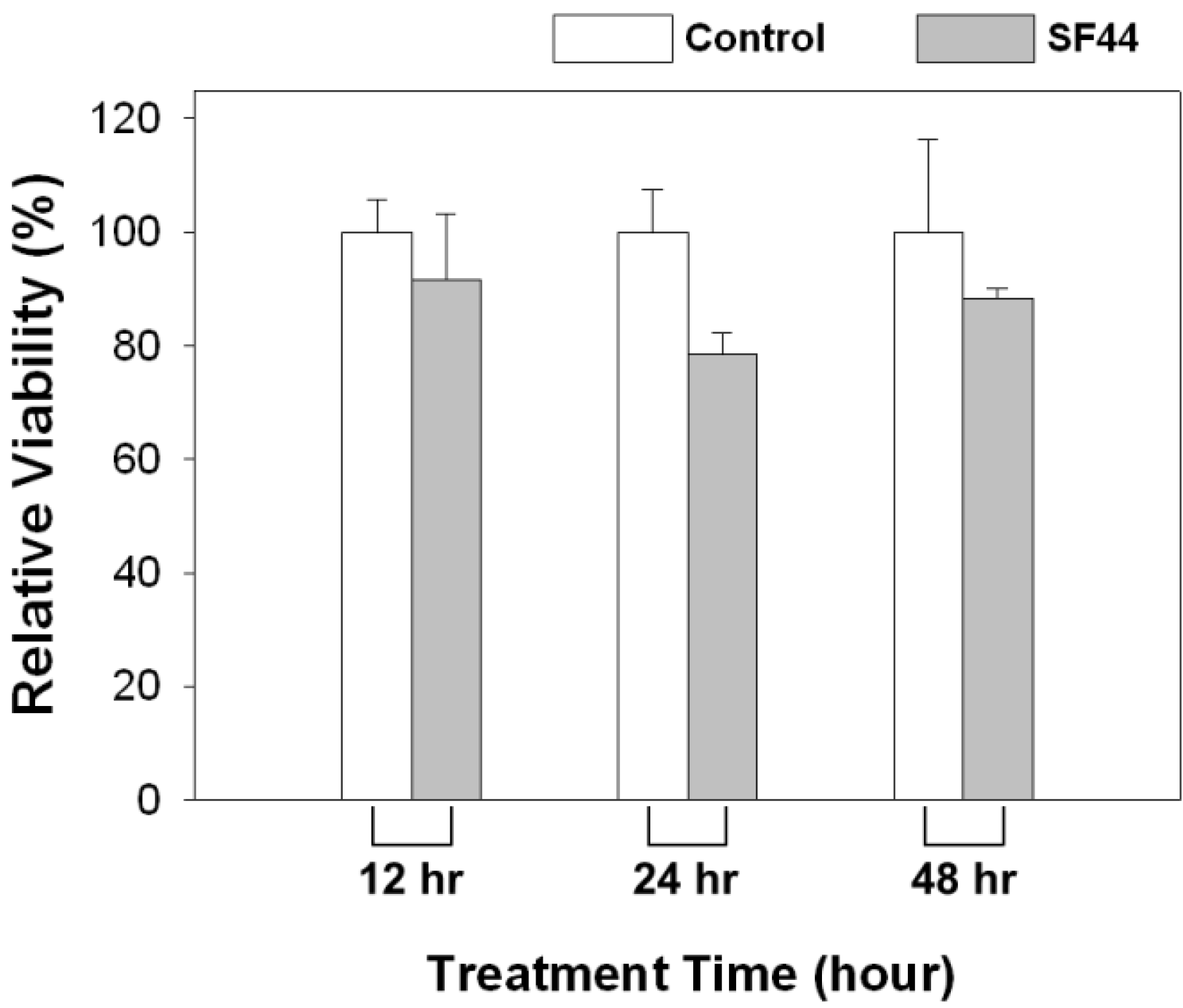
3.5. Viability Assay
3. 6. Liposome Concentration Measurement Using SF44
3.7. Determination of LDs Contents via Luminescence Spectrometry
3.8. LDs Purification and Size Measurement
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Oswald, W.J.; Golueke, C.G. Biological transformation of solar energy. Adv. Appl. Microbiol. 1960, 2, 223–262. [Google Scholar] [PubMed]
- Ghirardi, M.L.; Zhang, L.; Lee, J.W.; Flynn, T.; Seibert, M.; Greenbaum, E.; Melis, A. Microalgae: A green source of renewable H2. Trends Biotechnol. 2000, 18, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Akkerman, I.; Janssen, M.; Rocha, J.; Wijffels, R.H. Photobiological hydrogen production: Photochemical efficiency and bioreactor design. Int. J. Hydrog. Energy 2002, 27, 1195–1208. [Google Scholar] [CrossRef]
- Banerjee, A.; Sharma, R.; Chisti, Y.; Banerjee, U.C. Botryococcus braunii: A renewable source of hydrocarbons and other chemicals. Crit. Rev. Biotechnol. 2002, 22, 245–279. [Google Scholar] [CrossRef] [PubMed]
- Melis, A. Green alga hydrogen production: Progress, challenges and prospects. Int. J. Hydrog. Energy 2002, 27, 1217–1228. [Google Scholar] [CrossRef]
- Metzger, P.; Largeau, C. Botryococcus braunii: A rich source for hydrocarbons and related ether lipids. Appl. Microbiol. Biotechnol. 2005, 66, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Kay, R.A.; Barton, L.L. Microalgae as food and supplement. Crit. Rev. Food Sci. Nutr. 1991, 30, 555–573. [Google Scholar] [CrossRef] [PubMed]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metting, B.; Pyne, J.W. Biologically-active compounds from microalgae. Enzyme. Microb. Technol. 1986, 8, 386–394. [Google Scholar] [CrossRef]
- Schwartz, R.E.; Hirsch, C.F.; Sesin, D.F.; Flor, J.E.; Chartrain, M.; Fromtling, R.E.; Harris, G.H.; Salvatore, M.J.; Liesch, J.M.; Yudin, K. Pharmaceuticals from cultured algae. J. Ind. Microbiol. 1990, 5, 113–123. [Google Scholar] [CrossRef]
- Shimizu, Y. Microalgal metabolites: A new perspective. Annu. Rev. Microbiol. 1996, 50, 431–465. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y. Microalgal metabolites. Curr. Opin. Microbiol. 2003, 6, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, R.T.; Cysewski, G.R. Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends Biotechnol. 2000, 18, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kate, B.N.; Banerjee, U.C. Bioactive compounds from cyanobacteria and microalgae: An overview. Cri. Rev. Biotechnol. 2005, 25, 73–95. [Google Scholar] [CrossRef]
- Raja, R.; Hemaiswarya, S.; Kumar, N.A.; Sridhar, S.; Rengasamy, R. A perspective on the biotechnological potential of microalgae. Crit. Rev. Microbiol. 2008, 34, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Demirbas, A. Importance of biodiesel as transportation fuel. Energy Policy 2007, 35, 4661–4670. [Google Scholar] [CrossRef]
- Walther, T.C.; Farese, R.V., Jr. The life of lipid droplets. BBA-Mol. Cell Biol. Lipids 2009, 1791, 459–466. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method for total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Eltgroth, M.L.; Watwood, R.L.; Wolfe, G.V. Production and cellular localization of neutral long-chain lipids in the haptophyte algae Isochrysis galbana and Emiliania huxleyi. J. Phycol. 2005, 41, 1000–1009. [Google Scholar] [CrossRef]
- Izard, J.; Limberger, R.J. Rapid screening method for quantitation of bacterial cell lipids from whole cells. J. Microbiol. Meth. 2003, 55, 411–418. [Google Scholar] [CrossRef]
- Genicot, G.; Leroy, J.L.M.R.; Soom, A.V. The use of a fluorescent dye, Nile red, to evaluate the lipid content of single mammalian oocytes. Theriogenology 2005, 63, 1181–1194. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Yamaoka, M.; Kamisaka, Y. Rapid estimation of lipids in oleaginous fungi and yeasts using Nile red fluorescence. J. Microbiol. Meth. 2004, 56, 331–338. [Google Scholar] [CrossRef]
- Kamisaka, Y.; Noda, N.; Sakai, T.; Kawasaki, K. Lipid bodies and lipid body formation in an oleaginous fungus, Mortierella ramanniana var. angulispora. BBA-Mol. Cell Biol. Lipids 1999, 1438, 185–198. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, C.; Song, L.; Sommerfeld, M.; Hu, Q. A high throughput Nile red method for quantitative measurement of neutral lipids in microalgae. J. Microbiol. Meth. 2009, 77, 41–47. [Google Scholar] [CrossRef]
- Cooksey, K.E.; Guckert, J.B.; Williams, S.A.; Callis, P.R. Fluorometric-determination of the neutral lipid-content of microalgal cells using nile red. J. Microbiol. Meth. 1987, 6, 333–345. [Google Scholar] [CrossRef]
- Erickson, R.A.; Jimenez, R. Microfluidic cytometer for high-throughput measurement of photosynthetic characteristics and lipid accumulation in individual algal cells. Lab Chip 2013, 13, 2893–2901. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Lee, S.; Park, S.B. A Seoul-Fluor-based bioprobe for lipid droplets and its application in image-based high throughput screening. Chem. Commun. 2012, 48, 2331–2333. [Google Scholar] [CrossRef]
- Lee, Y.; Na, S.C.; Lee, S.; Jeon, N.L.; Park, S.B. Optimization of Seoul-Fluor-based lipid droplet bioprobe and its application in microalgae for bio-fuel study. Mol. Biosyst. 2013, 9, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Bollig, K.; Lamshöft, M.; Schweimer, K.; Marner, F.-J.; Budzikiewicz, H.; Waffenschmidt, S. Structural analysis of linear hydroxyproline-bound O-glycans of Chlamydomonas reinhardtii—Conservation of the inner core in Chlamydomonas and land plants. Carbohyd. Res. 2007, 342, 2557–2566. [Google Scholar] [CrossRef]
- Huang, Z.; Lee, H.; Lee, E.; Kang, S.-K.; Nam, J.-M.; Lee, M. Responsive nematic gels from the self-assembly of aqueous nanofibres. Nat. Commun. 2011, 2, 459. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.U.; Asami, Y.; Lee, D.; Jang, J.-H.; Ahn, J.S.; Oh, H. Protuboxepins A and B and protubonines A and B from the marine-derived fungus Aspergillus sp. SF-5044. J. Nat. Prod. 2011, 74, 1284–1287. [Google Scholar] [CrossRef] [PubMed]
- Hell, S.W. Toward fluorescence nanoscopy. Nat. Biotechnol. 2003, 21, 1347–1355. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.T.; Ullrich, N.; Joo, S.; Waffenschmidt, S.; Goodenough, U. Algal lipid biodies: Stress induction, purification, and biochemical characterization in wild-type and starchless Chlamydomonas reinhardtii. Eukaryot. Cell 2009, 8, 1856–1868. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Yoon, B.D.; Oh, H.M. Rapid method for the determination of lipid from the green alga Botryococcus braunii. Biotechnol. Tech. 1998, 12, 553–556. [Google Scholar] [CrossRef]
- Elsey, D.; Jameson, D.; Raleigh, B.; Cooney, M.J. Fluorescent measurement of microalgal neutral lipids. J. Microbiol. Meth. 2007, 68, 639–642. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Park, J.W.; Na, S.C.; Lee, Y.; Lee, S.; Park, S.B.; Jeon, N.L. Measurement of Lipid Droplet Accumulation Kinetics in Chlamydomonas reinhardtii Using Seoul-Fluor. Energies 2013, 6, 5703-5716. https://doi.org/10.3390/en6115703
Park JW, Na SC, Lee Y, Lee S, Park SB, Jeon NL. Measurement of Lipid Droplet Accumulation Kinetics in Chlamydomonas reinhardtii Using Seoul-Fluor. Energies. 2013; 6(11):5703-5716. https://doi.org/10.3390/en6115703
Chicago/Turabian StylePark, Jae Woo, Sang Cheol Na, Youngjun Lee, Sanghee Lee, Seung Bum Park, and Noo Li Jeon. 2013. "Measurement of Lipid Droplet Accumulation Kinetics in Chlamydomonas reinhardtii Using Seoul-Fluor" Energies 6, no. 11: 5703-5716. https://doi.org/10.3390/en6115703
APA StylePark, J. W., Na, S. C., Lee, Y., Lee, S., Park, S. B., & Jeon, N. L. (2013). Measurement of Lipid Droplet Accumulation Kinetics in Chlamydomonas reinhardtii Using Seoul-Fluor. Energies, 6(11), 5703-5716. https://doi.org/10.3390/en6115703