1. Introduction
The industrialized world’s primary infrastructure for transportation fuels and specialty chemical substrates utilizes petroleum based products. However, the basic economic principle of supply and demand has escalated petroleum prices, which is unavoidable for any important non-renewable natural resource. In short, global petroleum is not endless and the world’s energy consumption is projected to increase from 524 quadrillion British thermal units (Btu) to 820 quadrillion Btu (a 56% increase) between 2010 and 2040 [
1]. Further, society has become more sensitive to atmospheric CO
2 levels due to increased concern about rapid global climate change. Fossil fuels consumed globally contain 400 times the amount of annual carbon which can be fixed by the current global biota. Furthermore, fossil fuel consumption has increased annually from those levels [
2]. These numbers are staggering and if current practices and trends continue, traditional energy resources will be diminished much faster than regenerated and environments will be eroded/destroyed in an effort to supply these resources [
3]. However, advancement in renewable energy technologies can counteract part of this bleak scenario. Biodiesel, defined as fatty acid methyl esters (FAME) derived from plant, animal, or algal lipids, is an attractive alternative for portions of the current petroleum dependency. Algae have great potential for use in FAME production due to their inherent growth advantages, high TAG content per cell, and the potential to balance carbon levels; natural marine phytoplankton are responsible for 50% of the total global primary productivity [
4]. However, as was recently pointed out by the United States National Renewable Energy Laboratory, significant technological advancements that maximize lipid content must be made to improve the near-term industrial feasibility of utilizing microalgae as a biofuel feedstock [
5].
Algal TAG accumulation typically occurs when cellular cycling (the process of a parent cell splitting into daughter cells) is retarded or arrested due to a nutrient limitation [
6,
7,
8,
9,
10,
11], some environmental stress (e.g., pH, light, temperature stress) [
7,
12,
13,
14], or by chemical addition [
15,
16,
17]. However, this would appear to be counterproductive for industrial algal biofuel systems, which require rapid growth rates and a wide tolerance of environmental conditions (including extreme environments) [
18,
19]. Therefore, one optimal growth and TAG accumulation scenario for algal biofuel production involves high biomass productivity during the exponential growth phase and sequential arresting of the cell cycle, while still maintaining a high photosynthetic rate, at the onset of the stationary phase. Most production scenarios accomplish this by timing medium nitrogen depletion with a predetermined cellular density, while controlling the pH of the system with CO
2 addition. However, we and others have recently shown that some algal strains accumulate TAG at a higher rate when bicarbonate is added to culturing systems [
15,
20]. Furthermore, TAG and starch accumulation in
Chlamydomonas reinhardtii were shown to be more stable when accumulation occurred in a bicarbonate rich system [
16], which offers flexibility in the harvest timing. Finally, high bicarbonate concentrations can minimize contamination from invasive microorganisms [
21], which is especially important during longer periods of stress (
i.e., TAG accumulation) when cellular integrity may be compromised, and predators and/or bacteria can flourish on energy-rich cell debris.
Carbon assimilation by microalgae, through carbon concentrating mechanisms (CCMs), has been extensively studied [
22,
23,
24,
25,
26,
27,
28], and it is generally accepted that green algae can assimilate bicarbonate under certain environmental conditions (low CO
2 and alkaline conditions). Of particular note are carbon assimilation studies done on
Scenedesmus strains [
29,
30,
31], in which it was argued that bicarbonate is the major carbon source taken up at alkaline pH [
30]. However, these studies were performed using bicarbonate concentrations of approximately 10 µM–10 mM, which are significantly less than what was shown to induce TAG accumulation in
Scenedesmus sp. WC-1 (50 mM HCO
3−) [
15]. Due to this unconventional approach of enhancing TAG accumulation in microalgae, additional physiological research is needed to evaluate cellular responses, in terms of cell cycle inhibition, inorganic carbon utilization, and photosynthetic oxygen production, when high concentrations of bicarbonate are added to cultures.
This work reviews the current state of knowledge of Scenedesmus sp. WC-1 when bicarbonate is used to induce TAG accumulation. Results detailing TAG accumulation from sodium bicarbonate and sodium carbonate additions, at varying concentrations, or when combined are presented. The minimum concentration of bicarbonate required to arrest the cell cycle was determined. However, even these cultures were found to be carbon limited after 24 h. Therefore, abiotic and biotic studies were conducted to evaluate CO2 into a given medium, dissolved inorganic carbon utilization, and photosynthetic oxygen production during high bicarbonate culturing and when Scenedesmus sp. WC-1 was in a bicarbonate-induced TAG accumulation state. Taken together, fundamental physiological knowledge of cellular cycling, carbon utilization, and photosynthetic oxygen production in Scenedesmus sp. WC-1 during bicarbonate-induced TAG accumulation is further elucidated.
3. Results and Discussion
Previous research utilizing WC-1 evaluated the effects of pH, N-stress, combination of pH and N-stress, high CO
2 (5%
v/
v), and the addition of 50 mM sodium bicarbonate on the algal cellular cycle and on TAG accumulation [
12,
15].
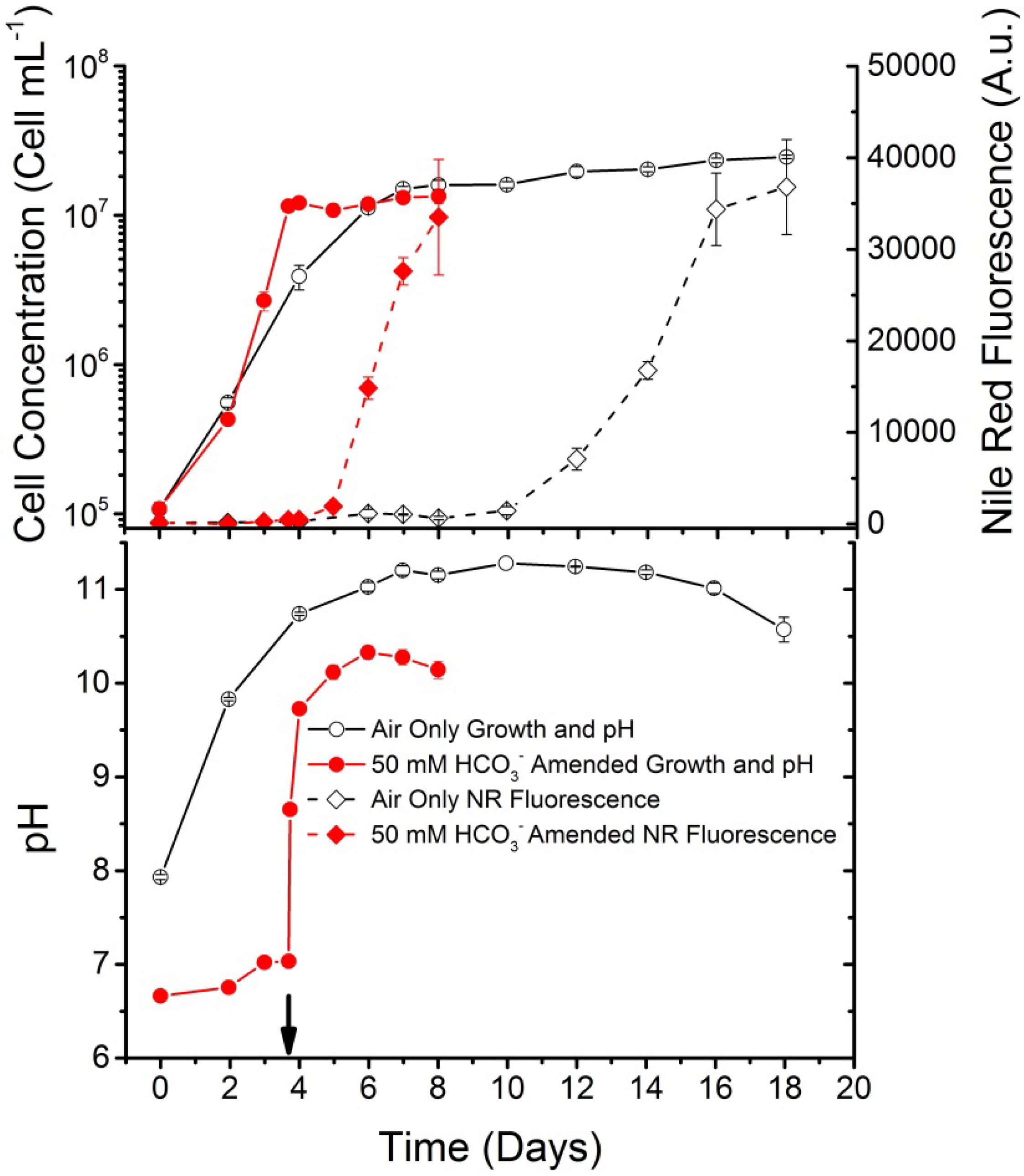
Figure 1, adapted from previously published work [
15], summarizes the use of elevated CO
2 and sodium bicarbonate to increase WC-1 growth rate and “trigger” TAG accumulation, as compared to cultures that only received inorganic carbon from an atmospheric air sparge. The main points to consider from this work are the following:
- (1)
Cultures which received elevated CO2 became nitrate deplete at approximately 3.7 days as compared to the cultures sparged with air, which became deplete at approximately 9 days;
- (2)
After the gas sparge was switched to atmospheric air and 50 mM sodium bicarbonate was added, there was a cessation of the cellular cycle and an increase in the TAG accumulation rate which resulted in a total culturing time of 8 days, to reach equivalent TAG accumulation, as compared to 16 days required for the other cultures. It should be noted that cultures which received elevated CO
2 (5%
v/
v) after nitrate became depleted reached a fluorescence level of only 3200 A.u. (data from [
15]);
- (3)
The added sodium bicarbonate buffered the cultures so that the pH was approximately one pH unit below the cultures which were only grown on air. This is important because the pKa for bicarbonate-carbonate equilibrium is pH 10.3 and at pH 11.3 the majority of the inorganic carbon in the system is carbonate, which is potentially unavailable for carbon fixation.
Thus, the use of sodium bicarbonate as an alternative inorganic carbon source and a TAG accumulation “trigger” has the potential to aid industrial algal biofuel production. However, further fundamental investigation is warranted to determine which ion (sodium, bicarbonate, or carbonate) is responsible for the algal metabolic changes. In addition, the minimal amount of sodium bicarbonate needed to arrest cellular replication and induce TAG accumulation must be identified. Finally, inorganic carbon use and photosynthetic properties associated with elevated sodium bicarbonate use must be investigated.
Figure 1.
Scenedesmus sp. WC-1 mean and standard deviation of cellular concentration, total Nile Red fluorescence, and pH of cultures aerated with ambient air and cultures which were sparged with 5% CO
2 (
v/
v, in air) until near nitrate depletion, at which time they were amended with 50 mM sodium bicarbonate and switched to ambient air. Arrow indicates time of the gas sparge change and bicarbonate addition. Growth was maintained in unbuffered Bold’s basal medium illuminated with a 14:10 L/D cycle (
n = 3). Figure adapted from [
15].
Figure 1.
Scenedesmus sp. WC-1 mean and standard deviation of cellular concentration, total Nile Red fluorescence, and pH of cultures aerated with ambient air and cultures which were sparged with 5% CO
2 (
v/
v, in air) until near nitrate depletion, at which time they were amended with 50 mM sodium bicarbonate and switched to ambient air. Arrow indicates time of the gas sparge change and bicarbonate addition. Growth was maintained in unbuffered Bold’s basal medium illuminated with a 14:10 L/D cycle (
n = 3). Figure adapted from [
15].
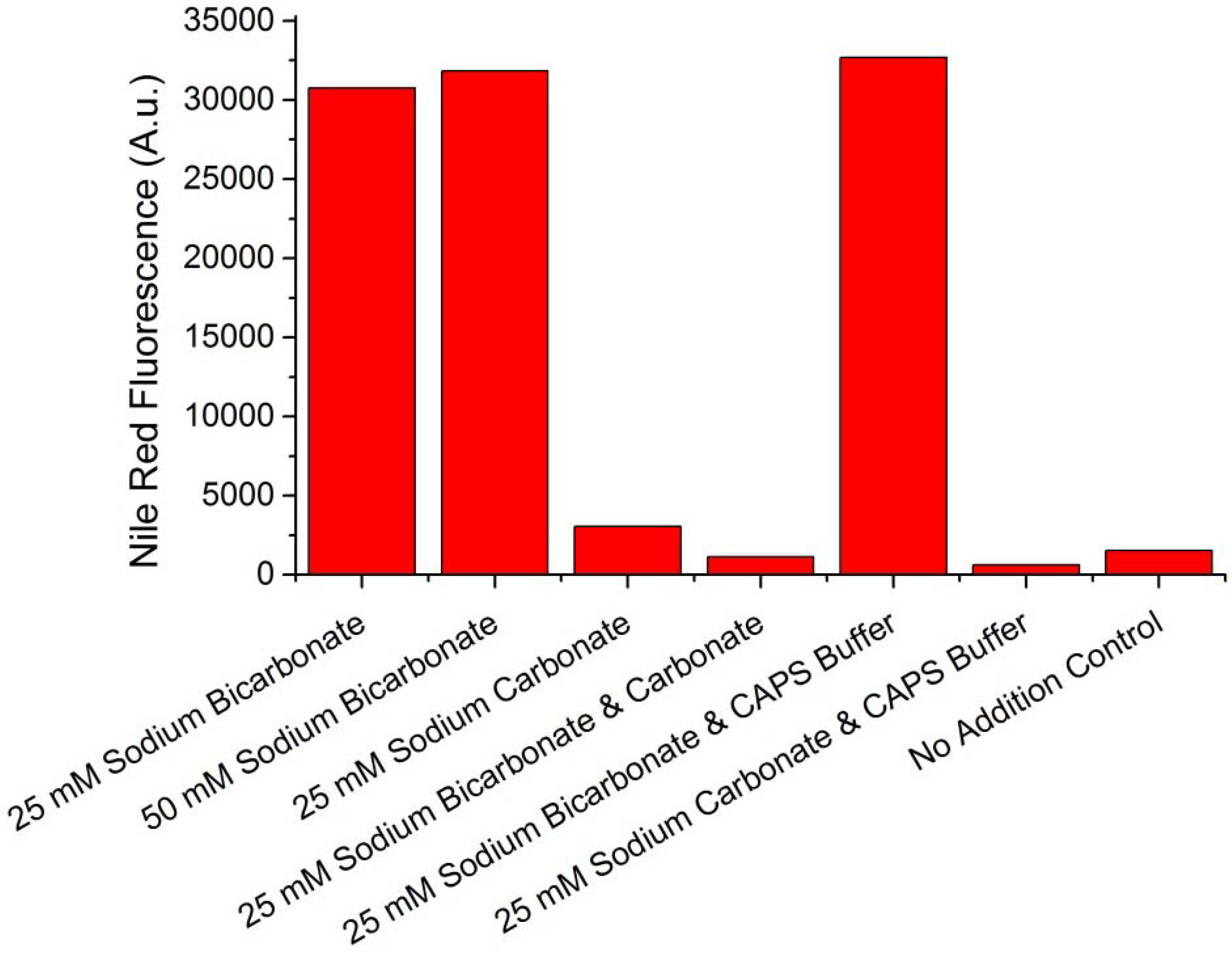
To assess which ion was responsible for the cessation of the cell cycle and subsequent TAG accumulation, WC-1 was screened for TAG accumulation when sodium bicarbonate and sodium carbonate were added individually or in combination, and when combined with 50 mM CAPS buffer (pKa 10.3). CAPS buffer was added in an effort to minimize changes of the culture pH. All additions resulted in a cessation of the cellular cycle, and Nile Red fluorescence values (
i.e., TAG accumulation) taken at the end of the experiments are shown in
Figure 2. All cultures which received bicarbonate or bicarbonate in addition to the CAPS buffer accumulated TAG. All cultures which received carbonate did not accumulate TAG, either when added individually or in combination with bicarbonate or CAPS buffer. The CAPS buffer was able to maintain the pH within a difference of 0.5 units between the cultures that received bicarbonate and carbonate, thus ion speciation differences were minimized. Comparison of the 25 mM sodium carbonate and the 50 mM sodium bicarbonate offers evidence that that bicarbonate ion is responsible for TAG accumulation and not the sodium ion. Sodium was present in equimolar concentration in these two treatments but only the bicarbonate system accumulated TAG. Additional evidence that bicarbonate, and not sodium, was influencing TAG accumulation is discussed below.
Figure 2.
Nile Red fluorescence responses of Scenedesmus sp. WC-1 cultures that were amended with sodium bicarbonate, sodium carbonate, and combinations of the two with and without 50 mM CAPS buffer (pKa 10.3) (values are for the conclusion of the experiments). Growth was maintained in unbuffered Bold’s basal medium illuminated with a 14:10 L/D cycle.
Figure 2.
Nile Red fluorescence responses of Scenedesmus sp. WC-1 cultures that were amended with sodium bicarbonate, sodium carbonate, and combinations of the two with and without 50 mM CAPS buffer (pKa 10.3) (values are for the conclusion of the experiments). Growth was maintained in unbuffered Bold’s basal medium illuminated with a 14:10 L/D cycle.
To begin optimizing algal systems for bicarbonate-induced TAG accumulation, the minimum amount of bicarbonate needed to change the cellular metabolism from high growth to fast TAG accumulation, must be identified.
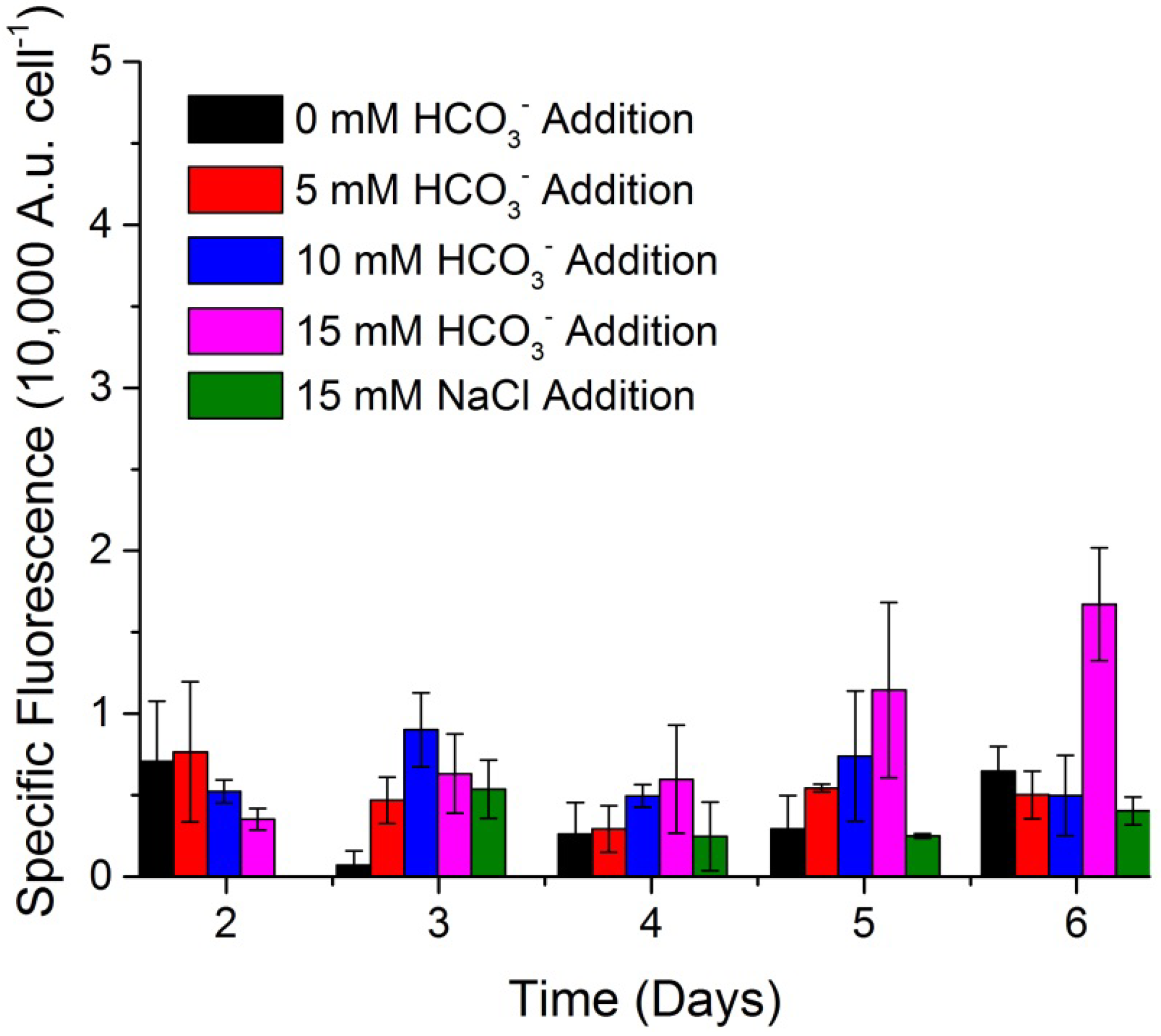
Figure 3 shows the growth characteristics of WC-1 when 0 mM, 5 mM, 10 mM, and 15 mM bicarbonate were added to WC-1 cultures. Additionally, the effect of sodium chloride addition was demonstrated to further verify the sodium ion is not responsible for the cessation of the cell cycle. All cultures were sparged with 5% CO
2 until near medium nitrate depletion, at which time the sparge was changed to atmospheric air and the salts were added. Cultures that received less than 10 mM bicarbonate and 15 mM sodium chloride continued to divide, producing new cells. The cultures that received 10 mM bicarbonate showed a delayed cell cycle, and the cultures that received 15 mM bicarbonate exhibited an arrested cell cycle.
Nile Red fluorescence was monitored over the course of the experiment as it is a rapid indication of TAG accumulation in WC-1 [
12].
Figure 4 shows the specific Nile Red fluorescence of WC-1 when 0 mM, 5 mM, 10 mM, and 15 mM bicarbonate, or 15 mM sodium chloride, were added to the cultures. Specific Nile Red fluorescence was calculated based on Equation (1), and represents the total Nile Red fluorescence normalized to the cell number. This number allows comparison of the average TAG content of each algal cell, and a large number represents a high TAG content:
where
sNR is the specific Nile Red (A.u. cell
−1),
NR is the total Nile Red signal of the algal culture (A.u. mL
−1), and
NC is cellular concentration (cell mL
−1). An arbitrary scaling factor of 10,000 has previously been used to shift results to 0–100 [
12,
15,
16].
Figure 3.
Scenedesmus sp. WC-1 mean growth and standard deviation of cultures which were sparged with 5% CO2 (v/v) in air and switched to ambient air, at which time they were amended with 0, 5, 10, and 15 mM sodium bicarbonate, along with 15 mM sodium chloride as a control amendment. The arrow indicates time of the gas sparge change and salt amendments. Growth was maintained in unbuffered Bold’s basal medium illuminated with a 14:10 L/D cycle (n = 3).
Figure 3.
Scenedesmus sp. WC-1 mean growth and standard deviation of cultures which were sparged with 5% CO2 (v/v) in air and switched to ambient air, at which time they were amended with 0, 5, 10, and 15 mM sodium bicarbonate, along with 15 mM sodium chloride as a control amendment. The arrow indicates time of the gas sparge change and salt amendments. Growth was maintained in unbuffered Bold’s basal medium illuminated with a 14:10 L/D cycle (n = 3).
Figure 4.
Mean and standard deviation of Scenedesmus sp. WC-1 specific Nile Red fluorescence in cultures which were sparged with 5% CO2 (v/v) in air and switched to ambient air, at which time they were amended with 0, 5, 10, and 15 mM sodium bicarbonate, along with 15 mM sodium chloride as a control amendment. The gas sparge was changed and salt amendments were added at 3 days. Growth was maintained in unbuffered Bold’s basal medium illuminated with a 14:10 L/D cycle (n = 3).
Figure 4.
Mean and standard deviation of Scenedesmus sp. WC-1 specific Nile Red fluorescence in cultures which were sparged with 5% CO2 (v/v) in air and switched to ambient air, at which time they were amended with 0, 5, 10, and 15 mM sodium bicarbonate, along with 15 mM sodium chloride as a control amendment. The gas sparge was changed and salt amendments were added at 3 days. Growth was maintained in unbuffered Bold’s basal medium illuminated with a 14:10 L/D cycle (n = 3).
As shown in
Figure 4, only the cultures which received a 15 mM bicarbonate addition began to accumulate TAG (6 days) after the gas sparge was shifted from 5% CO
2 to air and the addition of bicarbonate at 3 days. However, this addition resulted only in a specific Nile Red fluorescence of 1.7 ± 0.4 A.u. cell
−1 which is far below the previously observed specific Nile Red result of 24.8 ± 2.2 A.u. cell
−1 (error is one standard deviation,
n = 3) when 50 mM bicarbonate was used to induce TAG accumulation (with identical culturing conditions) [
15]. This suggests that while 15 mM bicarbonate was sufficient to stop the algal cell cycle, it was not sufficient to maximize TAG accumulation during the arrested cell cycle.
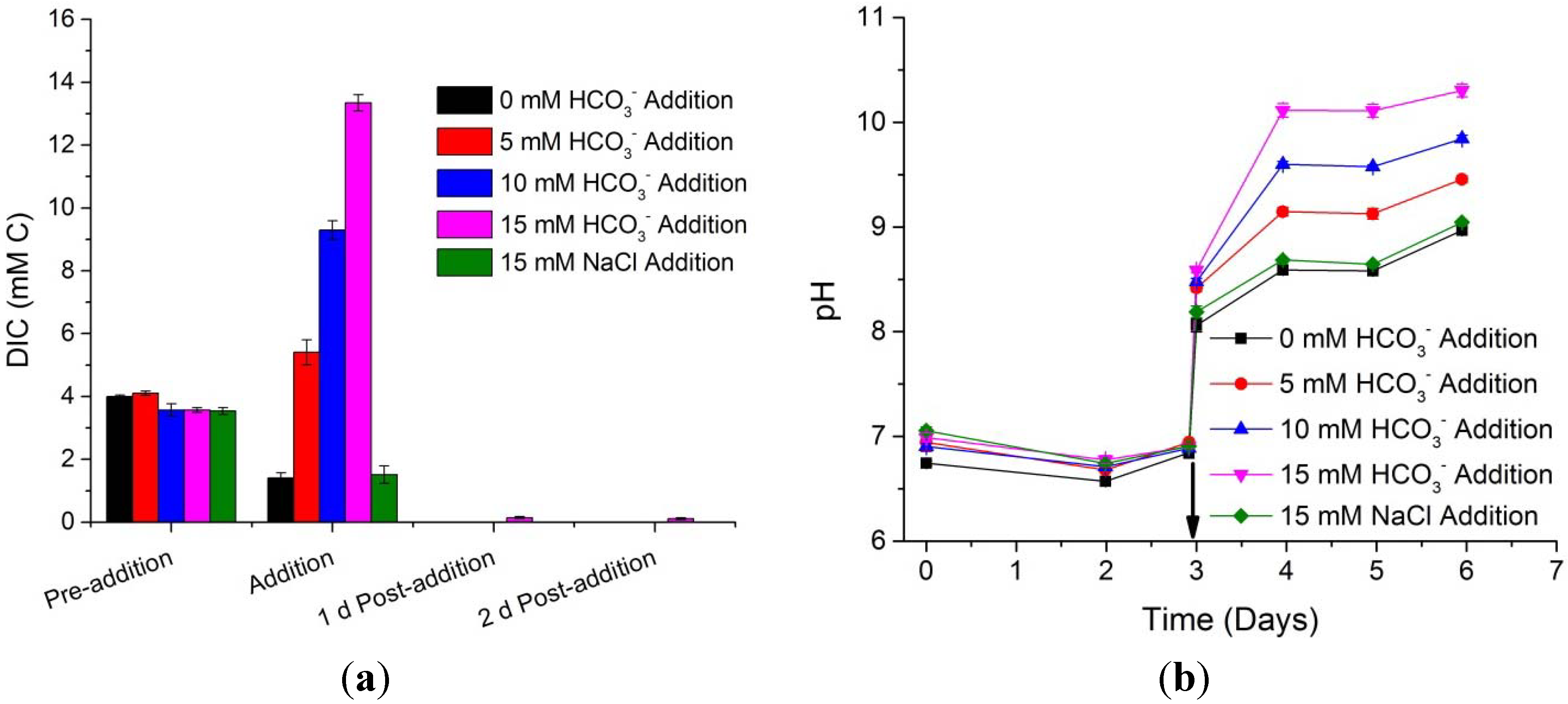
To advance our understanding of the total dissolved inorganic carbon (DIC, sum of the dissolved CO
2, bicarbonate, and carbonate) was monitored before the gas sparge change and bicarbonate additions, as well as two days post addition. As shown in
Figure 5, prior to the gas sparge change, when all cultures were receiving 5% CO
2, the DIC was 3.75 ± 0.3 mM as carbon in all the cultures. Directly after the gas sparge change to atmospheric CO
2 concentrations and the bicarbonate additions, the 0 mM, 5 mM, 10 mM, and 15 mM bicarbonate additions had 1.4, 5.4, 9.3, and 13.3 mM C, respectively. The 15 mM sodium chloride addition had 1.5 mM C and can be compared to the 0 mM bicarbonate experiment as the addition of sodium chloride did not add any extra inorganic carbon. Due to the rapid sampling directly after the gas sparge change (<3 min), the 0 mM bicarbonate additions and the 15 mM sodium chloride additions still had CO
2 remaining from the 5% CO
2 sparge. It is expected that the extra CO
2 did not remain in these systems for long after the measurements at the time of gas change and salt additions. The most striking finding is that the DIC in all the systems was completely utilized 1 day after the additions; effectively demonstrating the rapid carbon utilization and consumption capacity of this microalga.
Figure 5.
Mean and standard deviation of DIC (a) and culture pH (b) for Scenedesmus WC-1 cultures which were sparged with 5% CO2 in air and then switched to ambient air, at which time they were amended with 0, 5, 10, and 15 mM sodium bicarbonate, along with 15 mM sodium chloride as a control amendment. Arrow indicates time of the gas sparge change and salt amendments. Growth was maintained in unbuffered Bold’s basal medium illuminated with a 14:10 L/D cycle (n = 3).
Figure 5.
Mean and standard deviation of DIC (a) and culture pH (b) for Scenedesmus WC-1 cultures which were sparged with 5% CO2 in air and then switched to ambient air, at which time they were amended with 0, 5, 10, and 15 mM sodium bicarbonate, along with 15 mM sodium chloride as a control amendment. Arrow indicates time of the gas sparge change and salt amendments. Growth was maintained in unbuffered Bold’s basal medium illuminated with a 14:10 L/D cycle (n = 3).
In addition to the DIC information shown on
Figure 5, the culture pH was monitored over the course of the experiments (
Figure 5b). Before the gas change and salt additions, the pH remained at approximately pH 7 due to the acidification of the culture medium by the 5% CO
2. At the time of gas change and salt addition, there was a rapid increase in culture pH from the bicarbonate additions and the off gassing of CO
2. After which, and over the next 24 h (10:14 D/L cycle beginning at 3 days and ending at 4 days), the pH increased and was correlated with the bicarbonate addition. This was due to increased carbon utilization and hence increased photosynthetic activity for each respective bicarbonate load. However, after 4 days the pH stabilized due to the establishment of a steady-state between CO
2 supply (ingassing into the culture) and CO
2 consumption by the algal cells. It should be noted that during the dark hours CO
2 was net ingassing from the atmospheric air being supplied to the systems, and that in-gassing CO
2 will cause a decrease in culture pH; thus providing DIC for light dependent carbon fixation (
i.e., Calvin-Benson cycle).
When culturing on a diel cycle (L/D cycle), the addition of bicarbonate to induce TAG accumulation causes immediate changes in cell cycling, culture pH, buffering capacity, and the external DIC available to the alga for carbon fixation. While most of these parameters readily change throughout a typical algal culturing experiment (e.g., photosynthetically driven pH increase over time or external DIC decrease in availability as cultures become increasingly dense), the addition of bicarbonate causes rapid changes and complicates equilibrium prediction. Therefore, additional experimentation was pursued to elucidate CO
2 in- and out-gassing kinetics, culture pH buffering, and photosynthetic oxygen production when excess bicarbonate (over that needed to arrest cellular cycling) was added to induce TAG accumulation.
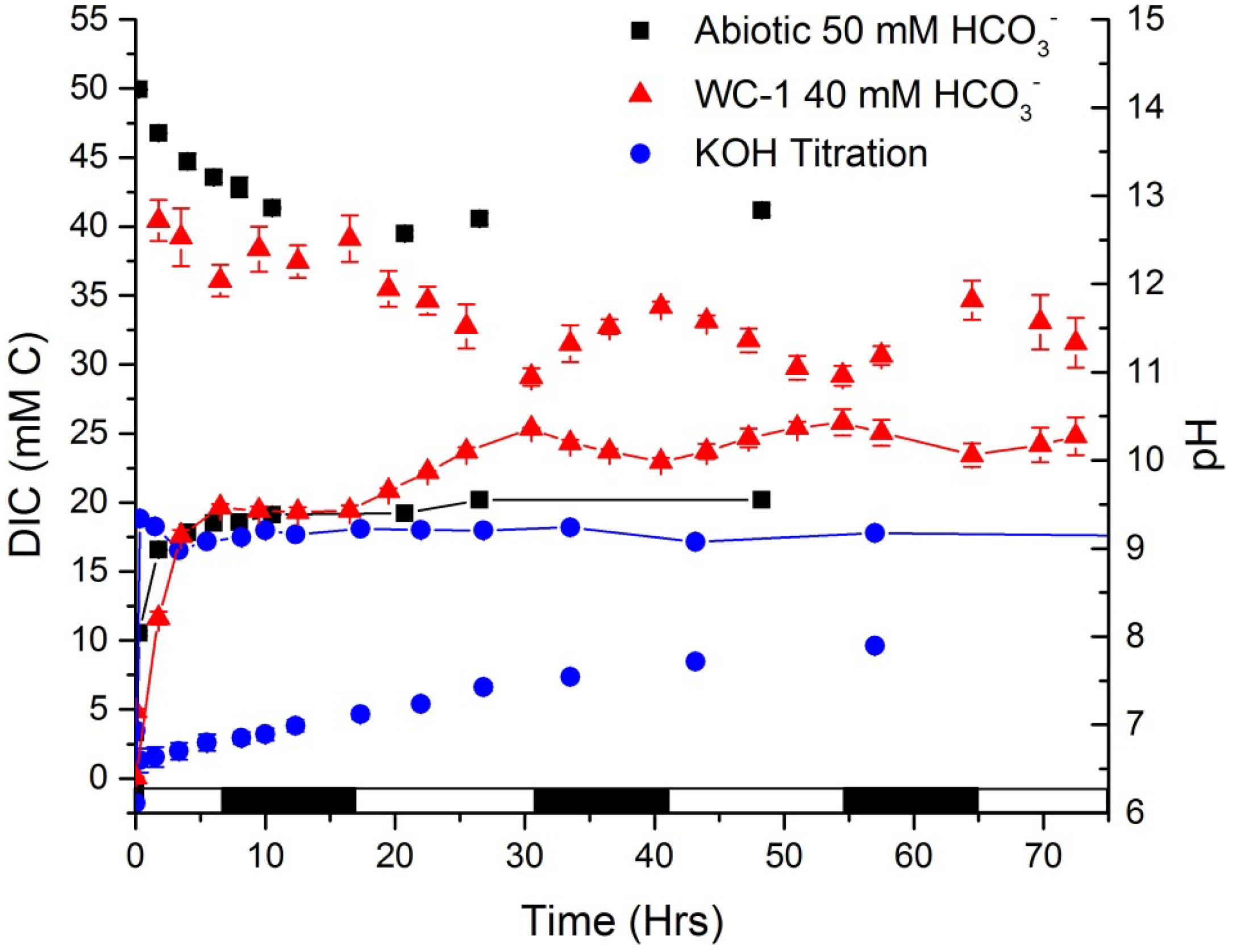
Figure 6 shows DIC and pH measurements over time for two abiotic experiments, one to monitor offgassing of CO
2 during bicarbonate addition and the other to monitor DIC ingassing during pH titration, and these are compared to a bicarbonate addition scenario in WC-1 cultures. An abiotic addition of 50 mM bicarbonate changed the medium pH from pH 6.8 to 8.3 which corresponds to almost 100% of the DIC being present as bicarbonate. Over the next 10 h, 10 mM C offgassed as CO
2 and the medium pH increased to pH 9.3. Conversely, when 50 mM CHES buffer and KOH were added to maintain the pH (KOH was added as needed) at approximately pH 9.3, carbon ingassed over 60 h to an equilibrium concentration of 11 mM C. Thus, there was approximately 30 mM more carbon in the system that received the bicarbonate amendment; also, equilibrium was reached in 10 h compared to 60 h for the pH titrated system.
In abiotic experiments equilibrium conditions are easily predicted; however, once a photosynthetic alga (
i.e., a sink term) is introduced to the system, a steady state condition is often approached, rather than the system reaching a state of equilibrium. Steady state conditions are difficult to predict, and may be significantly different from equilibrium, which is often assumed incorrectly. To visualize DIC and pH changes in a biotic system, 40 mM bicarbonate was added to WC-1 that was cultured on a 14:10 L/D cycle (
Figure 6). Fifty mM C was targeted for direct comparison to the abiotic system that was amended with bicarbonate. While this concentration was missed, similar trends in DIC in- or out-gassing and pH values were still observed. The amendment was made with 6.5 h remaining in the light cycle and there was a decrease of approximately 5 mM C due to offgassing of CO
2 and/or inorganic carbon fixation.
Figure 6.
Bold’s basal medium DIC (symbol only) and pH (symbol with line) when 50 mM NaHCO3 was added, when 50 mM CHES buffer (pKa 9.3) and KOH was added to titrate the pH to 9.3 (abiotic experiments, n = 1), or mean and standard deviation when 40 mM NaHCO3 was added to cultures of Scenedesmus WC-1 to induce TAG accumulation (biotic experiments, n = 3). Time axis begins at onset of abiotic experimentation or time of the gas sparge change and bicarbonate addition for the biotic experiment that was illuminated with a 14:10 L/D cycle (bottom bar).
Figure 6.
Bold’s basal medium DIC (symbol only) and pH (symbol with line) when 50 mM NaHCO3 was added, when 50 mM CHES buffer (pKa 9.3) and KOH was added to titrate the pH to 9.3 (abiotic experiments, n = 1), or mean and standard deviation when 40 mM NaHCO3 was added to cultures of Scenedesmus WC-1 to induce TAG accumulation (biotic experiments, n = 3). Time axis begins at onset of abiotic experimentation or time of the gas sparge change and bicarbonate addition for the biotic experiment that was illuminated with a 14:10 L/D cycle (bottom bar).
The pH of the cultures increased to pH 9.4. During the following 10 h dark cycle (starting at 6.5 h), the pH and DIC concentration remained statistically unchanged at 9.4 and 37 mM C, respectively. After the initial dark cycle there was a clear trend during the remaining light and dark cycles (beginning at 16.5 h) where the DIC concentration decreased during the light periods and then increased during the dark hours. Conversely, the pH in the system increased during the light hours and decreased in the dark. These trends indicate that during the light hours, DIC was consumed by the alga at a faster rate than CO
2 could ingas, otherwise the carbon concentration would be dictated by carbon dioxide and bicarbonate equilibrium and there would not have been an increase in DIC during the dark hours as the system tried to regain equilibrium DIC concentrations (with no photosynthesis taking place). Additionally, the DIC increased at a rate of 0.19 mM C h
−1 and 0.49 mM C h
−1 for the abiotic pH titrated system and for the cultures of WC-1 during the dark, respectively. The increased DIC ingassing rate of the biotic system can be explained by the increased alkalinity from the bicarbonate addition and the higher pH due to photosynthesis; however, it must be noted that respiration and other CO
2 producing metabolic processes may have increased the DIC concentration in the dark. Although, this would consume significant amounts of oxygen and that was not observed in similar experiments on WC-1 where dissolved oxygen concentrations were monitored over L/D cycles (
Figure 7 and discussed below).
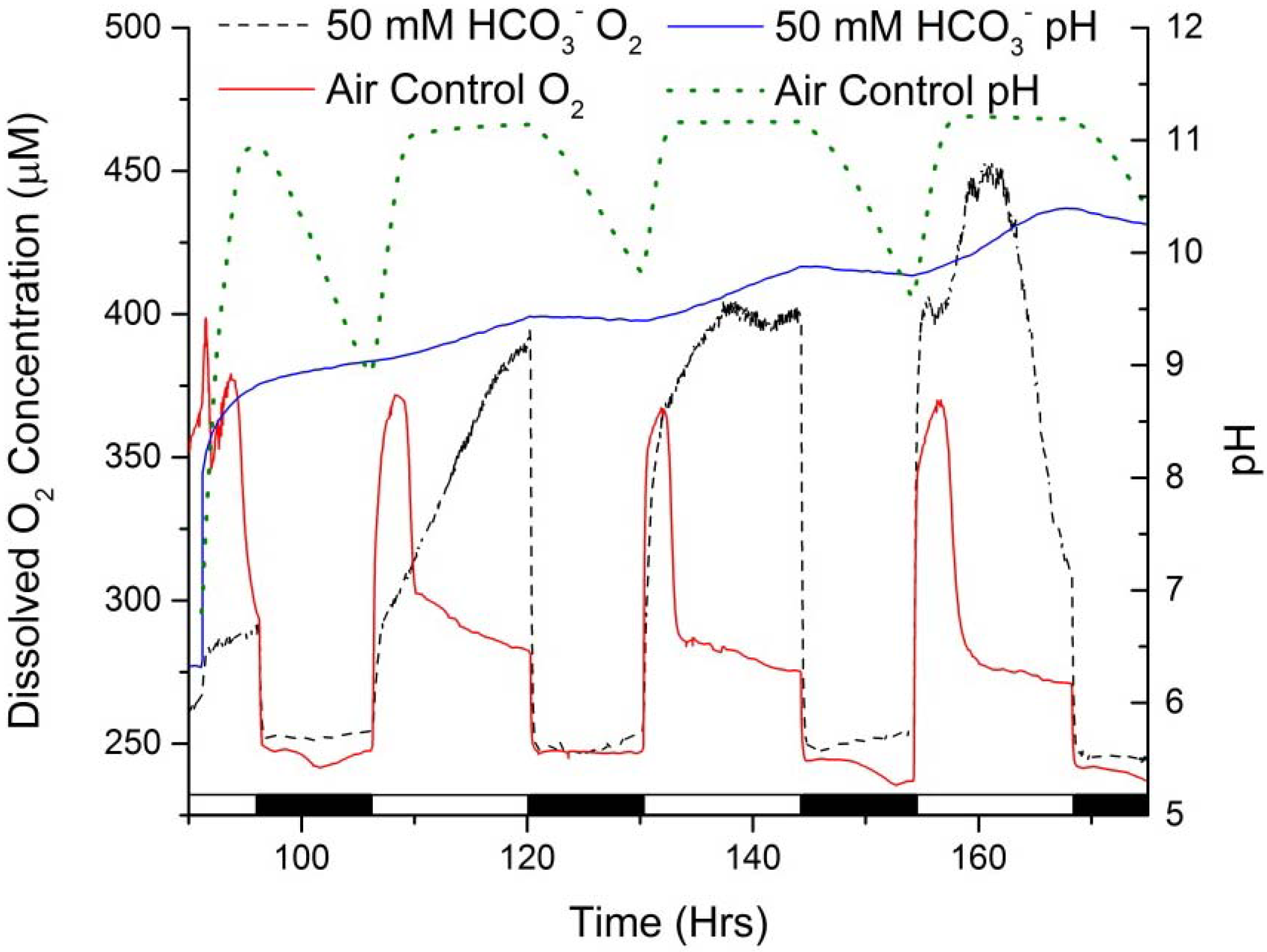
Figure 7.
Dissolved oxygen (DO) concentration and medium pH of Scenedesmus WC-1 cultures which were sparged with 5% CO2 (v/v) in air until near nitrate depletion, at which time (92 h) they were switched to ambient air and amended with 50 mM sodium bicarbonate. Comparison is made to cultures that did not receive the bicarbonate amendment. Growth was maintained in unbuffered Bold’s basal medium illuminated with a 14:10 L/D cycle (bottom bar) (representative individual pH and DO profiles for triplicate culturing, n = 3).
Figure 7.
Dissolved oxygen (DO) concentration and medium pH of Scenedesmus WC-1 cultures which were sparged with 5% CO2 (v/v) in air until near nitrate depletion, at which time (92 h) they were switched to ambient air and amended with 50 mM sodium bicarbonate. Comparison is made to cultures that did not receive the bicarbonate amendment. Growth was maintained in unbuffered Bold’s basal medium illuminated with a 14:10 L/D cycle (bottom bar) (representative individual pH and DO profiles for triplicate culturing, n = 3).
To further evaluate carbon fixation, photosynthetic oxygen production, and oxygenic respiration during the dark hours when bicarbonate is used to induce TAG accumulation in WC-1, experiments were conducted with data logging dissolved oxygen (DO) and pH probes.
Figure 7 shows representative pH and DO profiles for WC-1 cultures that were grown on 5% CO
2 until near nitrate depletion (92 h of culturing), at which time the gas sparge was switched to atmospheric air and half of the cultures were amended with 50 mM sodium bicarbonate. Directly after the gas sparge shift, the cultures to which bicarbonate was added increased to pH 8.3, and these cultures continued to increase in pH until the end of the light cycle (pH 9.2). The cultures that did not receive a bicarbonate amendment increased from pH 6.8 to pH 11 at the conclusion of the light cycle. After which, there was periodic decrease and sequential increase of the pH correlated directly to the light/dark cycle. The pH oscillated between pH 9.0 and pH 11.2 for the remainder of the experiment. The cultures to which bicarbonate was added also exhibited an increase in pH during the light hours and a sequential decrease during the dark hours, but the pH oscillation was significantly dampened due to the buffering capacity of the bicarbonate. The pH profile for the cultures to which 50 mM bicarbonate was added is nearly identical to the pH trend observed when 40 mM bicarbonate was added and the DIC was monitored over time (
Figure 6).
Most interesting (
Figure 7) are the trends in the DO and the difference between cultures amended with bicarbonate and those that were not amended. The cultures which did not receive bicarbonate initially had a higher photosynthetic oxygen production rate. However, during the start of each of the following light cycles, there was a short period of high oxygen production for approximately 4 h, after which the oxygen production rate drastically decreased. This correlates potentially with two factors: (1) the pH in these systems approaching pH 11 which effectively drives the DIC equilibrium towards carbonate, which minimizes carbon bioavailablity for carbon fixation; and (2) complete consumption of available DIC which had accrued during the dark as the system ingassed CO
2. The decreased DO concentration after 4 h, in each light cycle, was caused by carbon limitation; which in turn limited photosynthesis and hence DO production. The cultures to which bicarbonate was added increased in oxygen production over each sequential light cycle following the initial amendment. This could be due to increased biomass concentration (as opposed to cell number), or a higher chlorophyll per cell content; however, it must be noted that WC-1 did not increase in cell concentration after the bicarbonate amendment (
Figure 1). Another possible explanation is the onset of the operation of carbon concentrating mechanisms (CCMs) which employ carbonic anhydrase and bicarbonate transport proteins to effectively shuttle bicarbonate to the ribulose-1,5-bisphosphate carboxylase oxygenase (Rubisco) enzyme located in the pyrenoid of the chloroplast [
24,
25,
26,
27]. Besides the fundamental importance of increasing the carbon flux into microalgae, many studies have been done on the induction of algae’s CCM by shifting from high CO
2 (typically 1%–5%
v/
v) to atmospheric levels (0.04%) and they report 2–10 light hours are needed to synthesize the metabolic machinery necessary to concentrate inorganic carbon [
29,
30,
31,
39,
40,
41,
42,
43]. Further experimentation is needed to elucidate definitively the observed increase in photosynthetic oxygen production over time in the cultures which received additional bicarbonate. However, it should be noted that the total amount of photosynthetically-driven oxygen production was much greater for the cultures which received bicarbonate as compared to those that did not receive an amendment, and it is these cultures that accumulated more TAG. As a final observation, in the bicarbonate amended cultures there was no noticeable increase in DO consumption during the dark hours, which would be indicative of increased respiration and CO
2 production. This argues that the increase in DIC observed in the cultures of WC-1 during the dark hours was due to CO
2 ingassing and not from accelerated respiration (
Figure 6).
The use of sodium bicarbonate to enhance TAG accumulation has been demonstrated in both freshwater and marine chlorophytes as well as diatoms [
15,
16,
20,
44,
45], and its use offers inherent advantages. Gaseous CO
2 is costly to store and transport, whereas bicarbonate salts can easily be transported to algal facilities and stored until needed. Further, gas sparging systems are inefficient at delivering CO
2 into the culture medium in shallow raceways or ponds, with the majority of the CO
2 off-gassing into the atmosphere [
46]. However, at elevated pH, loss of inorganic carbon is drastically reduced if inorganic carbon is added. Additionally, with increasing concerns about greenhouse gases, algal systems are being proposed to sequester CO
2 from coal-fired power plants and other CO
2 point sources. If the emitted CO
2 is directly transferred to an algal growth facility, it may have to be stored during the night requiring extra capital cost for compressors and storage tanks. An alternative approach would be to temporarily sequester the CO
2 as bicarbonate which has the aforementioned storage and transport advantages [
47]. However, use of bicarbonate in this manner is new; thus further fundamental understanding and optimization is needed to determine effects on algal physiology, and evaluate cost comparisons.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






