Optimization of Alkaline Flocculation for Harvesting of Scenedesmus quadricauda #507 and Chaetoceros muelleri #862


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgal Strain and Culture Conditions
2.2. Determination of Flocculating Activity
2.3. Alkaline Flocculation
2.4. Starving the Cultures of CO2 Prior to Harvest
2.5. Effect of Synthetic Ocean Water on Flocculating Activity
2.6. Experimental Design and Statistical Analysis
3. Results and Discussion
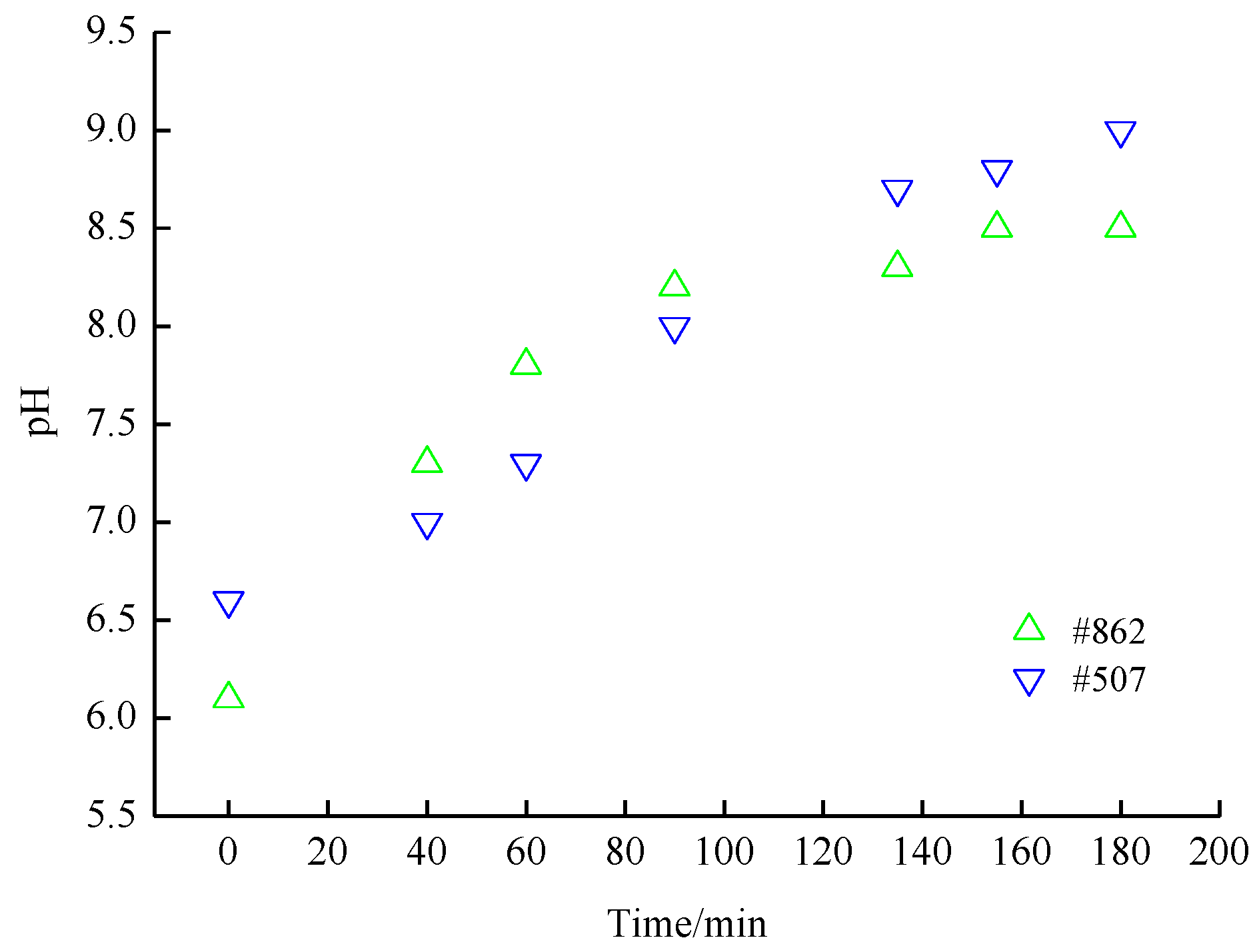
3.1. Starving the Cultures of CO2 Prior to Harvest

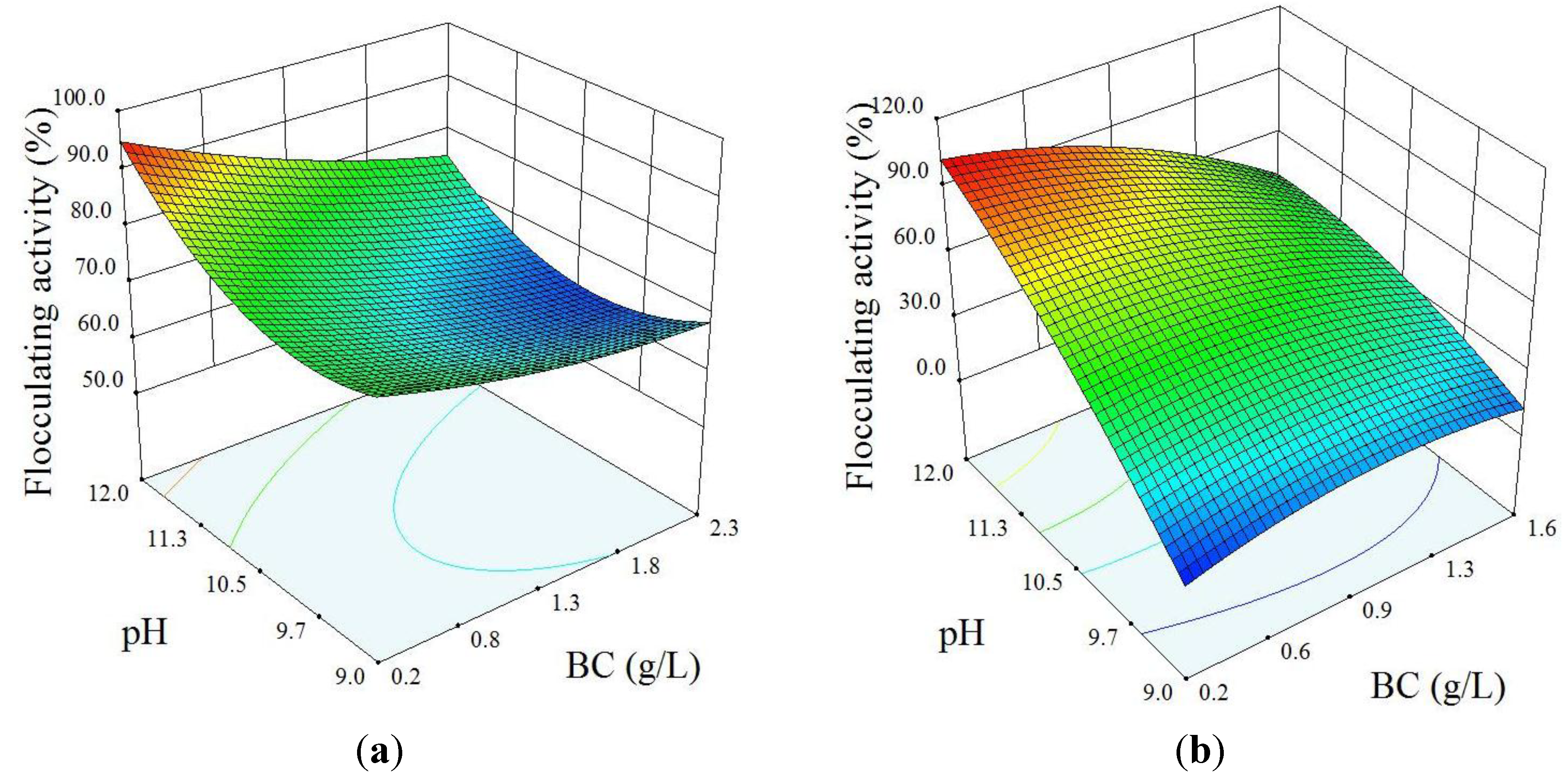
3.2. Optimization of Alkaline Flocculation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Sum of Squares | DF a | Mean Square | F-Value | p-value |
|---|---|---|---|---|---|
| Scenedesmus quadricauda #507 | |||||
| Model | 1,207.78 | 5 | 241.56 | 104.87 | <0.0001 |
| BC | 414.70 | 1 | 414.70 | 180.04 | <0.0001 |
| pH | 268.41 | 1 | 268.41 | 116.53 | <0.0001 |
| BC × pH | 26.11 | 1 | 26.11 | 11.34 | 0.0120 |
| BC2 | 26.38 | 1 | 26.38 | 11.45 | 0.0117 |
| pH2 | 493.46 | 1 | 493.46 | 214.23 | <0.0001 |
| Residual | 16.12 | 7 | 2.30 | – | – |
| Lack of Fit | 14.24 | 3 | 4.75 | 10.10 | 0.0245 |
| Pure Error | 1.88 | 4 | 0.47 | – | – |
| Cor Total | 1,223.90 | 12 | – | – | – |
| Chaetoceros muelleri #862 | |||||
| Model | 11,517.07 | 5 | 2,303.41 | 73.46 | <0.0001 |
| BC | 1,423.51 | 1 | 1,423.51 | 45.40 | 0.0003 |
| pH | 7,940.07 | 1 | 7,940.07 | 253.23 | <0.0001 |
| BC × pH | 1,216.27 | 1 | 1,216.27 | 38.79 | 0.0004 |
| BC2 | 600.57 | 1 | 600.57 | 19.15 | 0.0032 |
| pH2 | 457.43 | 1 | 457.43 | 14.59 | 0.0065 |
| Residual | 219.48 | 7 | 31.35 | – | – |
| Lack of Fit | 125.43 | 3 | 41.81 | 1.78 | 0.2903 |
| Pure Error | 94.05 | 4 | 23.51 | – | – |
| Cor Total | 11,736.55 | 12 | – | – | – |
| Factor | Coefficient Estimate | DF a | Standard Error | 95% CI b Low | 95% CI High | VIF c |
|---|---|---|---|---|---|---|
| Scenedesmus quadricauda #507 | ||||||
| Intercept | 68.94 | 1 | 0.68 | 67.33 | 70.54 | – |
| BC | −7.20 | 1 | 0.54 | −8.47 | −5.93 | 1.00 |
| pH | 5.79 | 1 | 0.54 | 4.52 | 7.06 | 1.00 |
| BC × pH | −2.56 | 1 | 0.76 | −4.35 | −0.76 | 1.00 |
| BC2 | 1.95 | 1 | 0.58 | 0.59 | 3.31 | 1.02 |
| pH2 | 8.42 | 1 | 0.58 | 7.06 | 9.78 | 1.02 |
| Chaetoceros muelleri #862 | ||||||
| Intercept | 57.50 | 1 | 2.50 | 51.58 | 63.42 | – |
| BC | −13.34 | 1 | 1.98 | −18.02 | −8.66 | 1.00 |
| pH | 31.50 | 1 | 1.98 | 26.82 | 36.19 | 1.00 |
| BC × pH | −17.44 | 1 | 2.80 | −24.06 | −10.82 | 1.00 |
| BC2 | −9.29 | 1 | 2.12 | −14.31 | −4.27 | 1.02 |
| pH2 | −8.11 | 1 | 2.12 | −13.13 | −3.09 | 1.02 |
| Strains | Equations | R2 | Adj-R2 | Pred-R2 |
|---|---|---|---|---|
| #507 a | Y = +811.5 + 15.5 × BC − 147.6 × pH − 3.3 × BC × pH + 3.6 × BC2 + 7.5 × pH2 | 0.987 | 0.977 | 0.915 |
| #862 b | Y = −1351.3 + 370.6 × BC + 209.4 × pH − 31.8 × BC × pH − 34.9 × BC2 − 7.2 × pH2 | 0.981 | 0.968 | 0.912 |
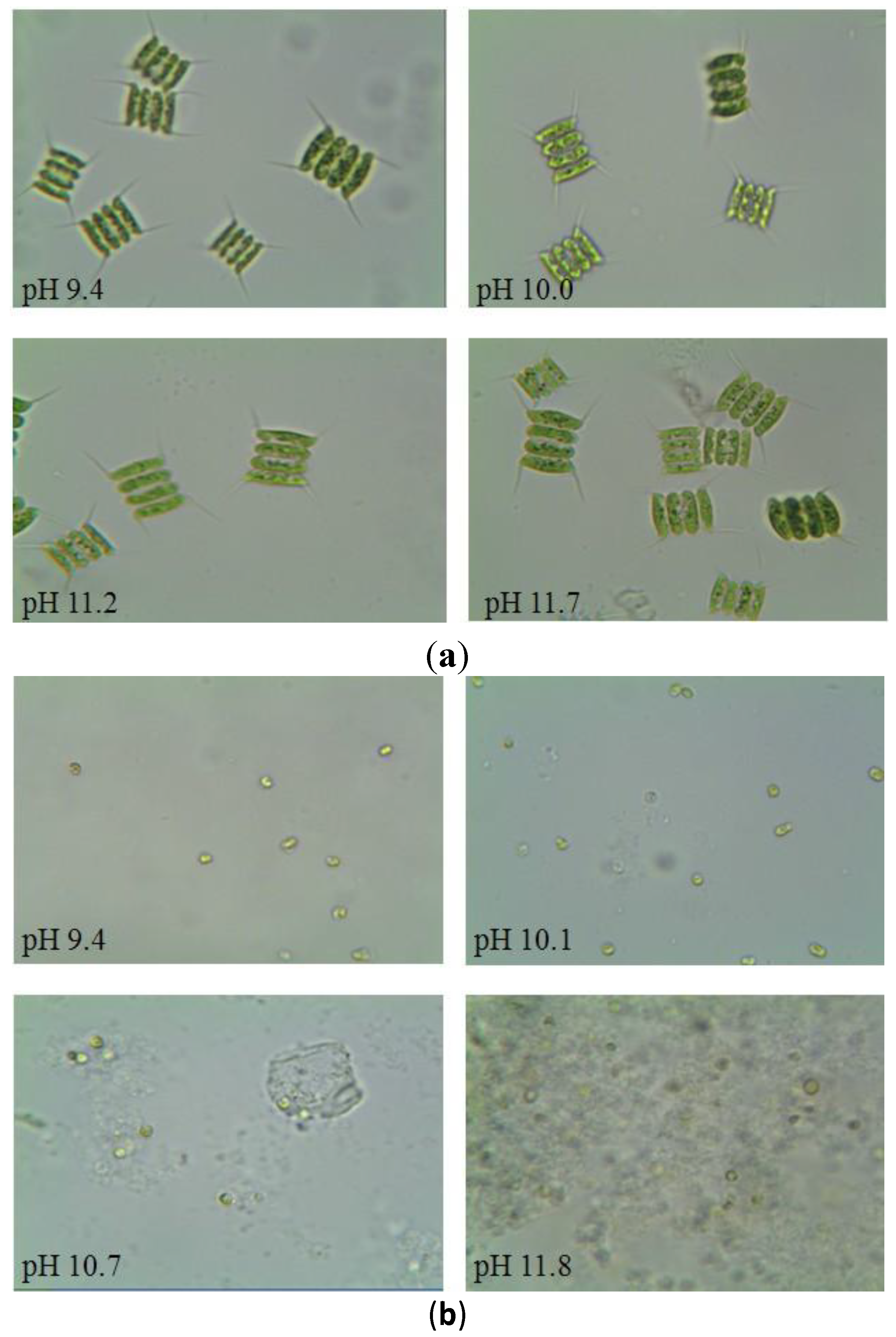
3.3. Microscopic Observation of Algal Cells with Increasing pH

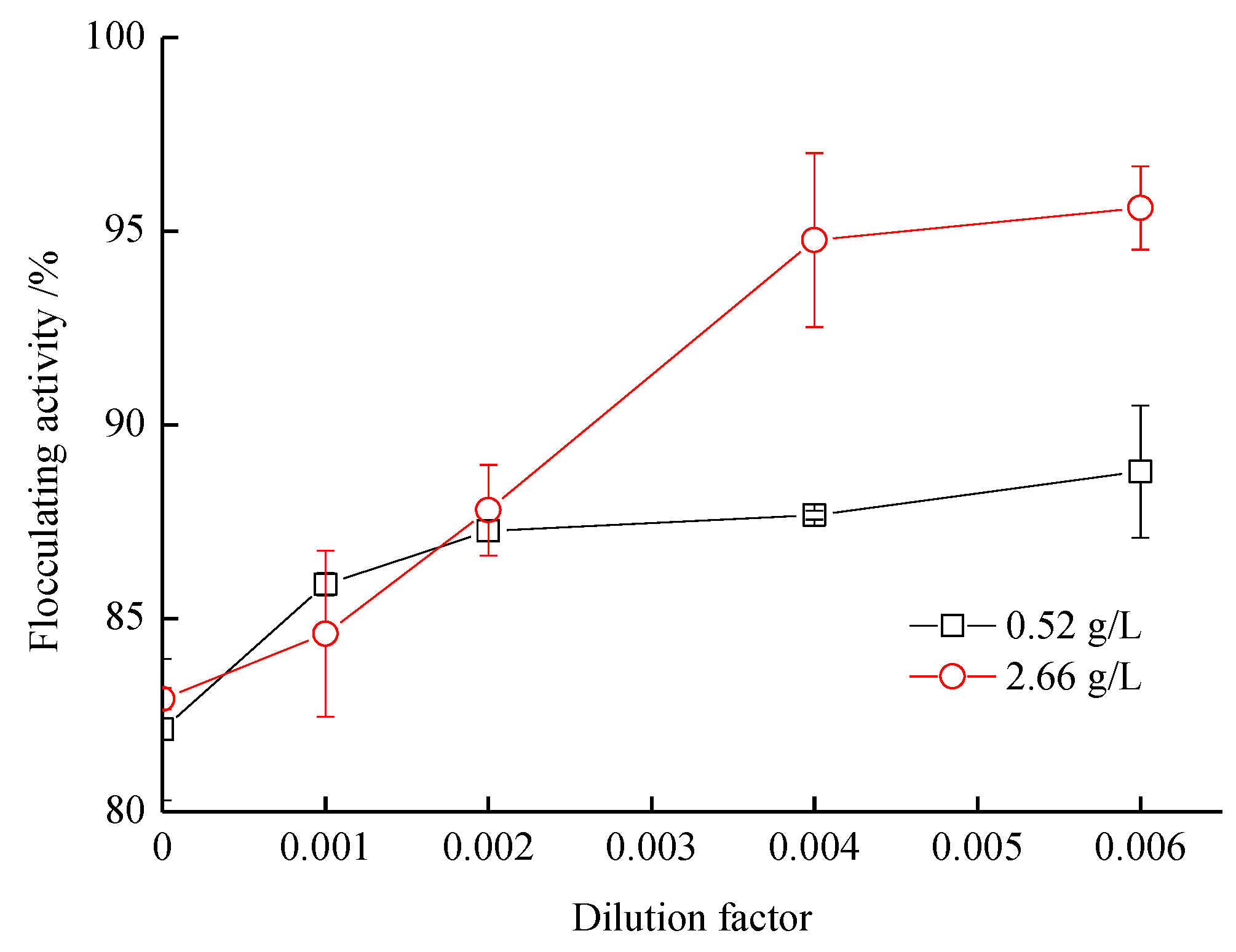
3.4. Improvement of Flocculating Activity with the Addition of Synthetic Ocean Water

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Demirbas, A.; Demirbas, M.F. Algae Energy: Algae as a New Source of Biodiesel; Springer-Verlag: London, UK, 2010. [Google Scholar]
- Huo, S.; Dong, R.; Wang, Z.; Pang, C.; Yuan, Z.; Zhu, S.; Chen, L. Available resources for algal biofuel development in China. Energies 2011, 4, 1321–1335. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Li, Y.; Wu, N.; Lan, C.Q. CO2 bio-mitigation using microalgae. Appl. Microbiol. Biotechnol. 2008, 79, 707–718. [Google Scholar] [CrossRef] [PubMed]
- National Algal Biofuel Technology Roadmap; U.S. Department of Energy, Office of Energy Efficiency and Renewable Energy, Biomass Program: Washington, DC, USA, 2010.
- Duan, J.; Gregory, J. Coagulation by hydrolysing metal salts. Adv. Colloid Interface Sci. 2003, 100–102, 475–502. [Google Scholar]
- Ahmad, A.L.; Mat Yasin, N.H.; Derek, C.J.C.; Lim, J.K. Optimization of microalgae coagulation process using chitosan. Chem. Eng. J. 2011, 173, 879–882. [Google Scholar] [CrossRef]
- Wu, Z.; Zhu, Y.; Huang, W.; Zhang, C.; Li, T.; Zhang, Y.; Li, A. Evaluation of flocculation induced by pH increase for harvesting microalgae and reuse of flocculated medium. Bioresour. Technol. 2012, 110, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Yahi, H.; Elmaleh, S.; Coma, J. Algal flocculation-sedimentation by pH increase in a continuous reactor. Water Sci. Technol. 1994, 30, 259–267. [Google Scholar]
- Schlesinger, A.; Eisenstadt, D.; Bar-Gil, A.; Carmely, H.; Einbinder, S.; Gressel, J. Inexpensive non-toxic flocculation of microalgae contradicts theories; overcoming a major hurdle to bulk algal production. Biotechnol. Adv. 2012, 30, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez, J.S.; Beuckels, A.; Vandamme, D.; Depraetere, O.; Foubert, I.; Parra, R.; Muylaert, K. Influence of magnesium concentration, biomass concentration and pH on flocculation of Chlorella vulgaris. Algal Res. 2014, 3, 24–29. [Google Scholar] [CrossRef]
- Vandamme, D.; Foubert, I.; Fraeye, I.; Meesschaert, B.; Muylaert, K. Flocculation of Chlorella vulgaris induced by high pH: Role of magnesium and calcium and practical implications. Bioresour. Technol. 2012, 105, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Besson, A.; Guiraud, P. High-pH-induced flocculation–flotation of the hypersaline microalga Dunaliella salina. Bioresour. Technol. 2013, 147, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, T.S.; Robert, H.D. Sedimentation of algae flocculated using naturally-available, magnesium-based flocculants. Algal Res. 2012, 1, 32–39. [Google Scholar] [CrossRef]
- Brady, P.V.; Pohl, P.I.; Hewson, J.C. A coordination chemistry model of algal autoflocculation. Algal Res. 2014, 5, 226–230. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.; Herdman, M.; Stanier, R. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Plenum Press: New York, NY, USA, 1975; pp. 26–60. [Google Scholar]
- Yang, Z.H.; Huang, J.; Zeng, G.M.; Ruan, M.; Zhou, C.S.; Li, L.; Rong, Z.G. Optimization of flocculation conditions for kaolin suspension using the composite flocculant of MBFGA1 and PAC by response surface methodology. Bioresour. Technol. 2009, 100, 4233–4239. [Google Scholar] [CrossRef] [PubMed]
- Blanchemain, A.; Grizeau, D.; Guary, J.C. Effect of different organic buffers on the growth of Skeletonema costatum cultures: Further evidence for an autoinhibitory effect. J. Plankton Res. 1994, 16, 1433–1440. [Google Scholar] [CrossRef]
- Lee, S.J.; Kim, S.B.; Kim, J.E.; Kwon, G.S.; Yoon, B.D.; Oh, H.M. Effects of harvesting method and growth stage on the flocculation of the green alga Botryococcus braunii. Lett. Appl. Microbiol. 1998, 27, 14–18. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huo, S.; Wang, Z.; Zhu, S.; Cui, F.; Zou, B.; You, W.; Yuan, Z.; Dong, R. Optimization of Alkaline Flocculation for Harvesting of Scenedesmus quadricauda #507 and Chaetoceros muelleri #862. Energies 2014, 7, 6186-6195. https://doi.org/10.3390/en7096186
Huo S, Wang Z, Zhu S, Cui F, Zou B, You W, Yuan Z, Dong R. Optimization of Alkaline Flocculation for Harvesting of Scenedesmus quadricauda #507 and Chaetoceros muelleri #862. Energies. 2014; 7(9):6186-6195. https://doi.org/10.3390/en7096186
Chicago/Turabian StyleHuo, Shuhao, Zhongming Wang, Shunni Zhu, Fengjie Cui, Bin Zou, Wenhua You, Zhenhong Yuan, and Renjie Dong. 2014. "Optimization of Alkaline Flocculation for Harvesting of Scenedesmus quadricauda #507 and Chaetoceros muelleri #862" Energies 7, no. 9: 6186-6195. https://doi.org/10.3390/en7096186
APA StyleHuo, S., Wang, Z., Zhu, S., Cui, F., Zou, B., You, W., Yuan, Z., & Dong, R. (2014). Optimization of Alkaline Flocculation for Harvesting of Scenedesmus quadricauda #507 and Chaetoceros muelleri #862. Energies, 7(9), 6186-6195. https://doi.org/10.3390/en7096186