Biological Aspects of Mountain Pine Beetle in Lodgepole Pine Stands of Different Densities in Colorado, USA

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Mountain Beetle Flight across Stands with Variable Densities
2.3. Attack Density, Insect Emergence, and Phloem Consumption
2.4. Probability of Individual Tree Attack
3. Results
3.1. Mountain Beetle Flight across Stands with Variable Densities
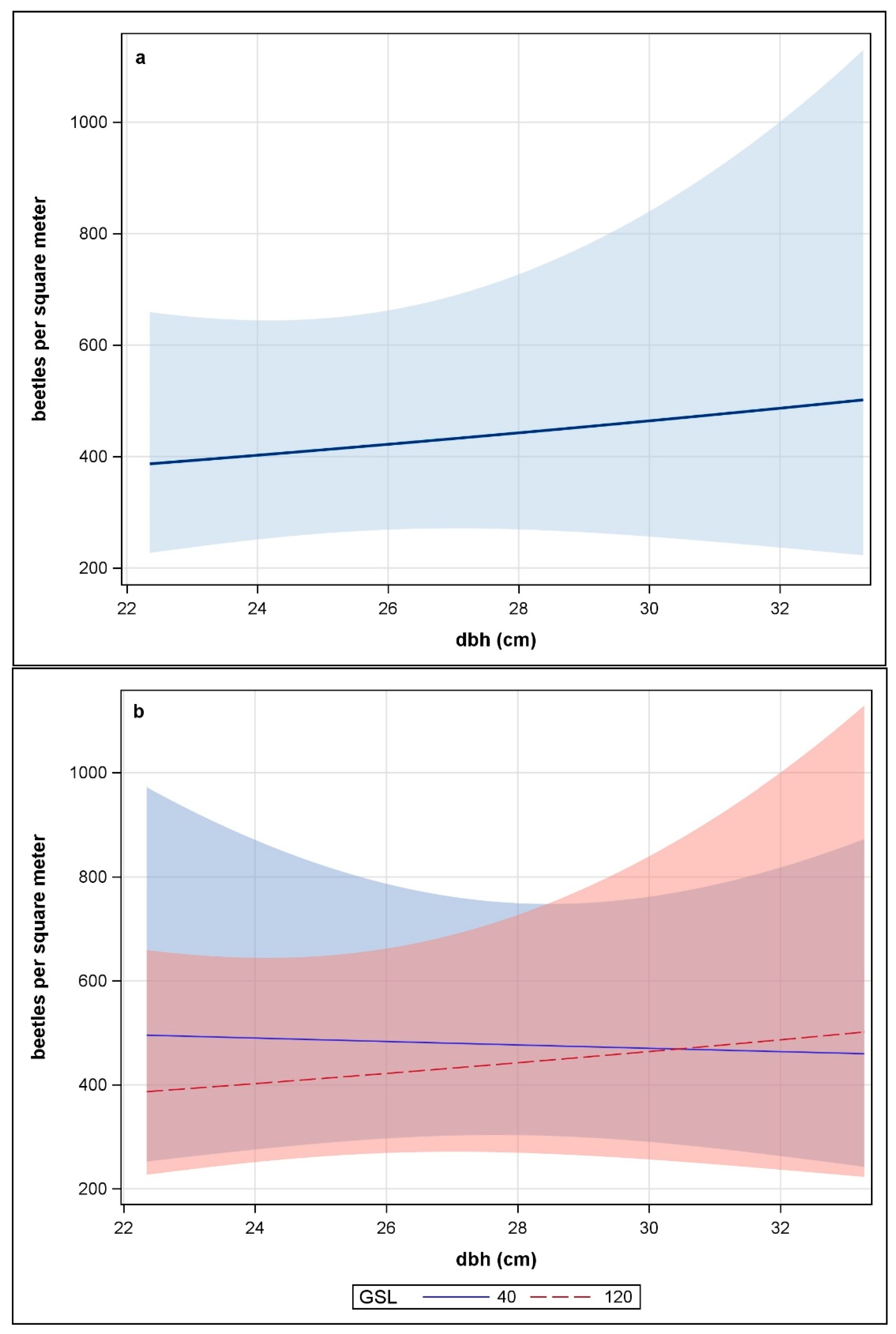
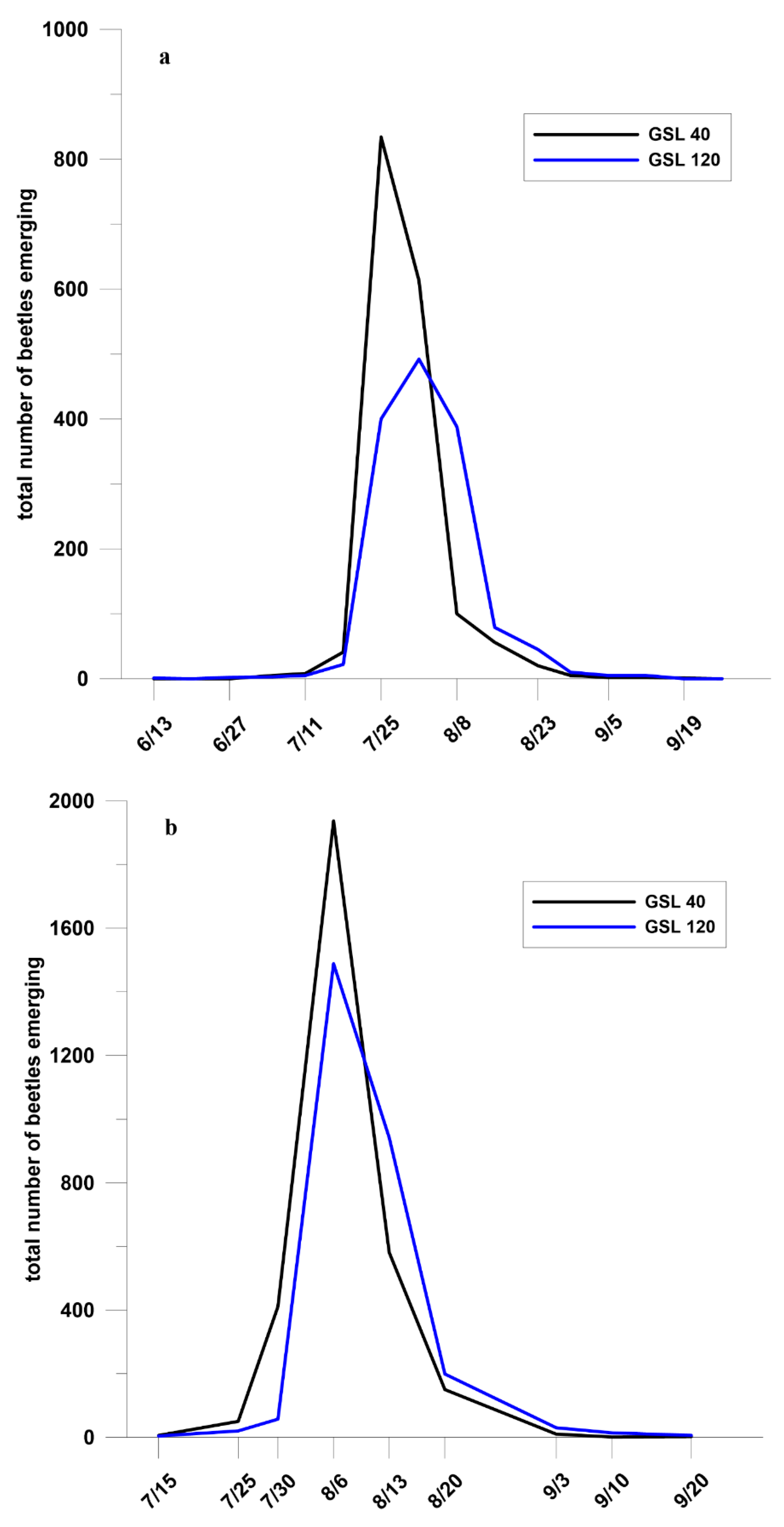
3.2. Attack Density, Insect Emergence, and Phloem Consumption
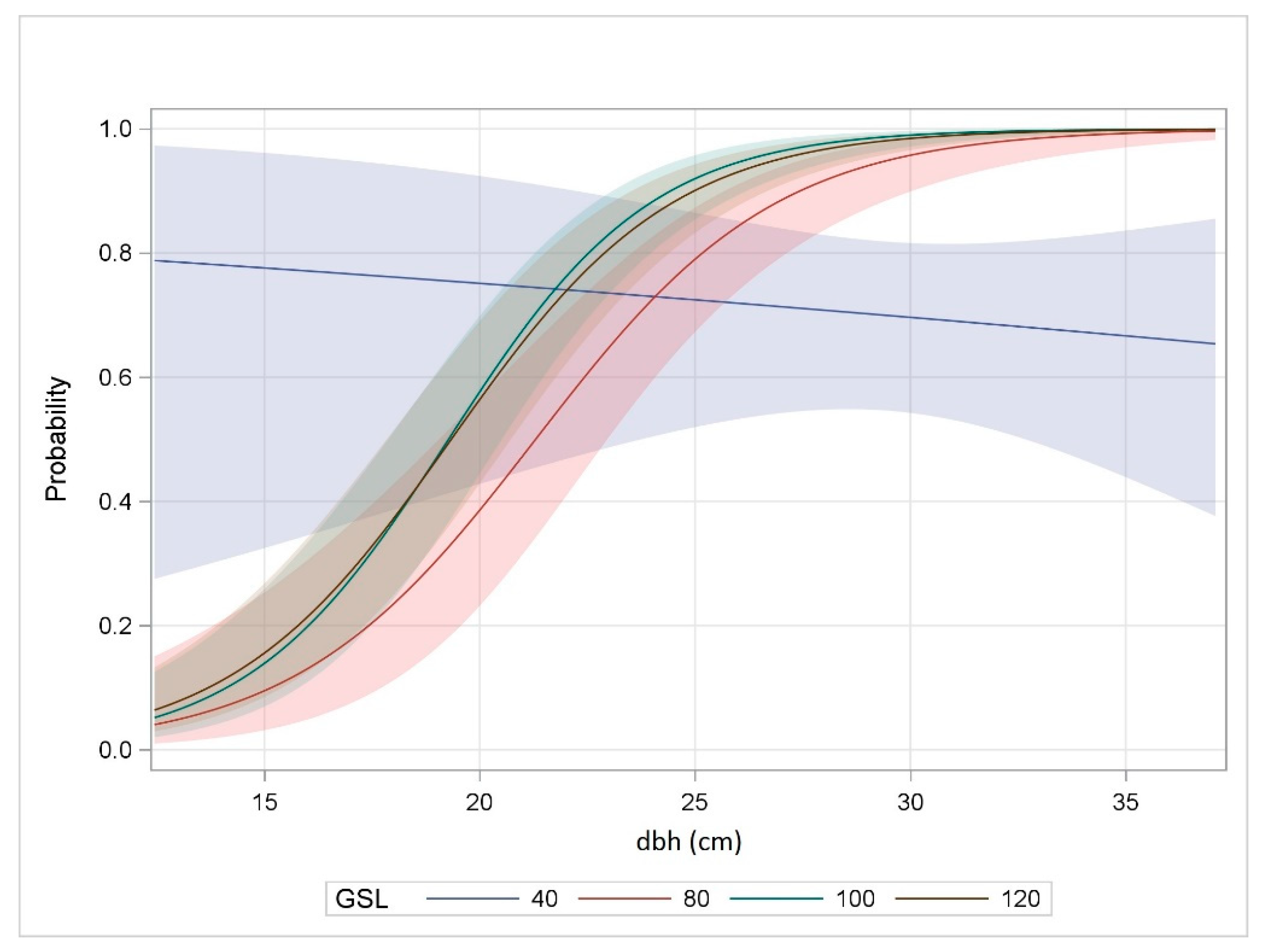
3.3. Probability of Individual Tree Attack
4. Discussion
4.1. Mountain Beetle Flight across Stands with Variable Densities
4.2. Attack Density, Insect Emergence, and Phloem Consumption
4.3. Probability of Individual Tree Attack
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Test of Fixed Effects | ||||
|---|---|---|---|---|
| Effect | Num DF | Den DF | F-Value | Pr |
| GSL | 2 | 35.9 | 14.5 | <0.0001 |
| Year | 2 | 35.9 | 17.5 | <0.0001 |
| GSL*Year | 4 | 35.39 | 0.4 | 0.78 |
| Test of Fixed Effects | |||||
|---|---|---|---|---|---|
| Response | Effect | Num DF | Den DF | F-Value | Pr |
| Number of attacks | GSL | 1 | 70.0 | 10.4 | 0.002 |
| No. of emerging beetles | GSL | 1 | 76.9 | 0.56 | 0.46 |
| dbh | GSL | 1 | 76.8 | 8.8 | 0.004 |
| Phloem consumption | GSL | 1 | 36 | 1.3 | 0.26 |
| Test of Fixed Effects | ||||
|---|---|---|---|---|
| Effect | Num DF | Den DF | F-Value | Pr |
| dbh | 1 | 75.4 | 0.07 | 0.79 |
| GSL | 1 | 74.9 | 0.30 | 0.59 |
| dbh*GSL | 1 | 74.9 | 0.24 | 0.63 |
| Test of Fixed Effects | ||||
|---|---|---|---|---|
| Effect | Num DF | Den DF | F-Value | Pr |
| dbh | 1 | 1407 | 105.4 | <0.0001 |
| GSL | 3 | 1407 | 8.2 | <0.0001 |
| dbh*GSL | 3 | 1407 | 13.1 | <0.0001 |
References
- Wood, S.L. The bark and ambrosia beetles of North and Central America (Coleoptera: Scolytidae), a taxonomic monograph. Great Basin Nat. Mem. 1982, 6, 1359. [Google Scholar]
- Costello, S.L.; Schaupp, W.C. First Nebraska state record of the mountain pine beetle, Dendroctonus ponderosae Hopkins (Coleoptera: Curculionidae: Scolytinae). Coleopts. Bull. 1988, 65, 21–23. [Google Scholar] [CrossRef]
- De la Giroday, H.C.; Carroll, A.L.; Aukema, B.H. Breach of the northern Rocky Mountain geoclimatic barrier: Initiation of range expansion by the mountain pine beetle. J. Biogeogr. 2012, 39, 112–1123. [Google Scholar] [CrossRef]
- Carroll, A.L.; Taylor, S.W.; Régnière, J.; Safranyik, L. Effects of climate change on range expansion by the mountain pine beetle in British Columbia. In Mountain Pine Beetle Symposium: Challenges and Solutions, Kelowna, British Columbia, Canada, 30–31 October 2003; Shore, T.L., Brooks, J.E., Stone, J.E., Eds.; Natural Resources Canada, Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 2003; pp. 223–232. [Google Scholar]
- Cullingham, C.I.; Cooke, J.E.K.; Dang, S.; Davis, C.S.; Cooke, B.J.; Coltman, D.W. Mountain pine beetle host-range expansion threatens the boreal forest. Mol. Ecol. 2011, 20, 2157–2171. [Google Scholar] [CrossRef] [PubMed]
- Furniss, M.M.; Schenk, J.A. Sustained natural infestations by the mountain pine beetle in seven new Pinus and Picea hosts. J. Econ. Entomol. 1969, 62, 518–519. [Google Scholar] [CrossRef]
- Negrón, J.F.; Fettig, C.J. Mountain pine beetle, a major disturbance agent in US western coniferous forests: A synthesis of the state of knowledge. For. Sci. 2014, 60, 409–413. [Google Scholar] [CrossRef]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: Dynamics of biome-wide bark beetle eruptions. Bioscience 2008, 58, 501–518. [Google Scholar] [CrossRef]
- Dordel, J.; Feller, M.C.; Simard, S.W. Effects of mountain pine beetle (Dendroctonus ponderosae Hopkins) infestations on forest stand structure in the southern Canadian Rocky Mountains. For. Ecol. Manag. 2008, 255, 3563–3570. [Google Scholar] [CrossRef]
- Axelson, J.N.; Alfaro, R.I.; Hawkes, B.C. Influence of fire and mountain pine beetle on the dynamics of lodgepole pine stands in British Columbia, Canada. For. Ecol. Manag. 2009, 257, 1874–1882. [Google Scholar] [CrossRef]
- Klutsch, J.G.; Negrón, J.F.; Costello, S.L.; Rhoades, C.C.; West, D.R.; Popp, J.; Caissie, R. Stand characteristics and downed woody debris accumulations associated with a mountain pine beetle (Dendroctonus ponderosae Hopkins) outbreak in Colorado. For. Ecol. Manag. 2009, 258, 641–649. [Google Scholar] [CrossRef]
- Negrón, J.F.; Cain, B. Mountain pine beetle in Colorado: A story of changing forests. J. For. 2018. [Google Scholar] [CrossRef]
- Colorado State Forest Service. 2014 Report on the Health of Colorado’s Forests. Colorado State Forest Service, Fort Collins, CO, USA. Available online: www.csfs.colostate.edu (accessed on 15 May 2018).
- Amman, G.D.; McGregor, M.D.; Cahill, D.B.; Klein, W.H. Guidelines for Reducing Losses of Lodgepole Pine to the Mountain Pine Beetle in Unmanaged Stands in the Rocky Mountains; General Technical Report, INT-GTR-262; US Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1977; p. 19.
- Shore, T.L.; Safranyik, L. Susceptibility and Risk Rating Systems for the Mountain Pine Beetle in Lodgepole Pine Stands; Information Report, BC-X-336; Forestry Canada, Pacific Forestry Centre: Victoria, BC, Canada, 1992; p. 12.
- Boone, C.K.; Aukema, B.H.; Bohlmann, J.; Carroll, A.L.; Raffa, K.F. Efficacy of tree defense physiology varies with bark beetle population density: A basis for positive feedback in eruptive species. Can. J. For. Res. 2011, 41, 1174–1188. [Google Scholar] [CrossRef]
- Waring, R.H.; Pitman, G.B. Modifying lodgepole pine stands to change susceptibility to mountain pine beetle attack. Ecology 1985, 66, 889–897. [Google Scholar] [CrossRef]
- Safranyik, L.; Shrimpton, D.M.; Whitney, H.S. Management of Lodgepole Pine to Reduce Losses from the Mountain Pine Beetle; Forestry Technical Report, 1; Environment Canada, Forestry Service, Pacific Forest Research Centre: Victoria, BC, Canada, 1974; p. 24.
- Amman, G.D. Mountain pine beetle brood production in relation to thickness of lodgepole pine phloem. J. Econ. Entomol. 1972, 65, 138–140. [Google Scholar] [CrossRef]
- Shrimpton, D.M.; Thomson, A.J. Relationship between phloem thickness and lodgepole pine growth characteristics. Can. J. For. Res. 1985, 15, 1004–1008. [Google Scholar] [CrossRef]
- Bartos, D.L. Microclimate as a factor in lodgepole pine (Pinus contorta Douglas) stands and tree selection by mountain pine beetles (Dendroctonus ponderosae Hopkins). In Proceedings of IUFRO Working Party and XVII International Congress of Entomology, Blacksburg, VA, USA, 4 July; Payne, T.L., Saarenmaa, H., Eds.; Department of Entomology, Virginia Tech University: Blacksburg, VA, USA, 1988; pp. 215–230. [Google Scholar]
- Bartos, D.L.; Amman, G.D. Microclimate: An Alternative to Tree Vigor as a Basis for Mountain Pine Beetle Infestations; Research Paper, INT-RP-400; US Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1989; p. 10.
- Amman, G.D.; Logan, J.A. Silvicultural control of mountain pine beetle: Prescriptions and the influence of microclimate. Am. Entomol. 1998, 44, 166–177. [Google Scholar] [CrossRef]
- Geiszler, D.R.; Gara, R.W. Mountain pine beetle attack dynamics in lodgepole pine. In Theory and Practice of Mountain Pine Beetle Management in Lodgepole Pine Forests, Symposium Proceedings, Pullman, WA, USA, 25–27 April 1978; Berryman, A.A., Amman, G.D., Stark, R.W., Eds.; University of Idaho Forest, Wildlife and Range Experiment Station: Moscow, ID, USA; U.S. Department of Agriculture, Forest Service, Forest Insect and Disease Research: Washington, DC, USA, 1978; pp. 182–187. [Google Scholar]
- Whitehead, R.J.; Russo, G.L. “Beetle-Proofed” Lodgepole Pine Stands in Interior British Columbia have Less Damage from Mountain Pine Beetle; Information Report, BC-X-402; Natural Resources Canada, Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 2005; p. 17.
- Thistle, H.W.; Peterson, H.; Allwine, G.; Lamb, B.; Strand, T.; Holsten, E.H.; Shea, P.J. Surrogate pheromone plumes in three forest trunk spaces: Composite statistics and case studies. For. Sci. 2004, 50, 610–625. [Google Scholar]
- Schmid, J.M.; Mata, S.A. Natural Variability of Specific Forest Insect Populations and their Associated Effects in Colorado; General Technical Report, RM-GTR-275; US Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1996; p. 14.
- Jarvis, D.S.; Kulakowski, D. Long-term history and synchrony of mountain pine beetle outbreaks in lodgepole pine forests. J. Biogeogr. 2015, 42, 1029–1039. [Google Scholar] [CrossRef]
- McCambridge, W.F.; Hawksworth, F.G.; Edminster, C.B.; Laut, J.G. Ponderosa Pine Mortality Resulting from a Mountain Pine Beetle Outbreak; Research Paper, RM-RP-235; US Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1982; p. 7.
- Negrón, J.F.; Popp, J.B. Probability of ponderosa pine infestation by mountain pine beetle in the Colorado Front Range. For. Ecol. Manag. 2004, 191, 17–27. [Google Scholar] [CrossRef]
- Colorado State Forest Service. 2011 Report on the Health of Colorado’s Forests. Colorado State Forest Service, Fort Collins, CO, USA. Available online: www.csfs.colostate.edu (accessed on 15 April 2018).
- Alexander, R.R.; Taclle, D.; Dahms, W.G. Site Indices for Lodgepole Pine, with Corrections for Stand Density: Methodology; Research Paper, RM-RP-29; US Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1967; p. 18.
- Huckaby, L.S.; Moir, W.H. Forest communities at Fraser Experimental Forest, Colorado. Southwest Nat. 1998, 43, 201–208. [Google Scholar]
- Tishmack, J.; Mata, S.A.; Schmid, J.M. Mountain Pine Beetle Emergence from Lodgepole Pine at Different Elevations Near Fraser, CO; Research Note, RMRS-RN-27; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2005; p. 5.
- McCambridge, W.F. Emergence Period of Black Hills Beetles from Ponderosa Pine in the Central Rocky Mountains; Research Note, RM-RN-32; US Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1964; p. 4.
- West, D.R.; Briggs, J.S.; Jacobi, W.R.; Negrón, J.F. Mountain pine beetle host selection between lodgepole and ponderosa pines in the southern Rocky Mountains. Environ. Entomol. 2016, 45, 127–141. [Google Scholar] [CrossRef] [PubMed]
- West, D.R.; Briggs, J.S.; Jacobi, W.R.; Negrón, J.F. Mountain pine beetle-caused mortality over eight years in two pine hosts in mixed-conifer stands of the southern Rocky Mountains. For. Ecol. Manag. 2014, 334, 321–330. [Google Scholar] [CrossRef]
- Mata, S.A.; Schmid, J.A.; Olsen, W.K. Growth of Lodgepole Pine Stands and its Relation to Mountain Pine Beetle Susceptibility; Research Paper, RMRS-RP-42; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2003; p. 19.
- Negrón, J.F.; Klutsch, J.G. Probability of Infestation and Extent of Mortality Models for Mountain Pine Beetle in Lodgepole Pine Forests in Colorado; Research Note, RMRS-RN-77; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2017; p. 13.
- Ross, D.; Daterman, G. Using pheromone-baited traps to control the amount and distribution of tree mortality during outbreaks of the Douglas-fir beetle. For. Sci. 1997, 43, 65–70. [Google Scholar]
- Furniss, M.M.; Kegley, S.J. Douglas-fir Beetle, Forest Insect and Disease Leaflet 5; US Department of Agriculture, Forest Service, Pacific Northwest Region (R6): Portland, Oregon, USA, 2014; p. 12.
- Lessard, E.D.; Schmid, J.M. Emergence, attack densities, and host relationships for the Douglas-fir beetle (Dendroctonus pseudotsugae Hopkins) in northern Colorado. Great Basin Nat. 1990, 50, 333–338. [Google Scholar]
- Negrón, J.F.; Schaupp, W.C., Jr.; Pederson, L. Flight periodicity of the Douglas-fir beetle, Dendroctonus pseudotsugae Hopkins (Coleoptera: Curculionidae: Scolytinae) in Colorado, U.S.A. Coleopts. Bull. 2011, 65, 182–184. [Google Scholar]
- Bentz, B.J.; Bracewell, R.R.; Mock, K.E.; Pfrender, M.E. Genetic architecture and phenotypic plasticity of thermally-regulated traits in an eruptive species, Dendroctonus ponderosae. Evol. Ecol. 2011, 25, 1269–1288. [Google Scholar] [CrossRef]
- Bentz, B.J.; Logan, J.A.; Vandygriff, J.C. Latitudinal variation in Dendroctonus ponderosae (Coleoptera: Scolytidae) development time and adult size. Can. Entomol. 2001, 133, 375–387. [Google Scholar] [CrossRef]
- Rehfeldt, G.E. A Spline Model of Climate for the Western United States; General Technical Report, RMRS-GTR-165; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2006; p. 21.
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. Global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Bentz, B.J.; Regniere, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate change and bark beetles of the western United States and Canada: Direct and indirect effects. Bioscience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Seidl, R.; Spies, T.A.; Peterson, D.L.; Stephens, S.L.; Hicke, J.A. Searching for resilience: Addressing the impacts of changing disturbance regimes on forest ecosystem services. J. Appl. Ecol. 2015, 53, 120–129. [Google Scholar] [CrossRef]
- Prism Climate Group. Available online: http://prism.oregonstate.edu/ (accessed on 28 April 2015).
- Myers, C.A. Growing Stock Levels in Even-Aged Ponderosa Pine; Research Paper, RM-RP-33; US Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1967; p. 8.
- SAS Institute. SAS/STAT 9.4 User’s Guide; SAS Institute: Cary, NC, USA, 2016. [Google Scholar]
- Lindgren, B.S. A multiple funnel trap for scolytid beetles (Coleoptera). Can. Entomol. 1983, 115, 299–302. [Google Scholar] [CrossRef]
- Reid, R.W. Biology of the mountain pine beetle, Dendroctonus monticolae Hopkins, in the east Kootenay region of British Columbia. III. Interaction between the beetle and its host, with emphasis on brood mortality and survival. Can. Entomol. 1963, 95, 225–238. [Google Scholar] [CrossRef]
- Rasmussen, L.A. Emergence and Attack Behavior of the Mountain Pine Beetle in Lodgepole Pine; Research Note, INT-RN-297; US Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1980; p. 7.
- Negrón, J.F.; Shepperd, W.D.; Mata, S.A.; Popp, J.B.; Asherin, L.A.; Schoettle, A.W.; Schmid, J.M.; Leatherman, D.A. Solar Treatment for Reducing Survival of Mountain Pine Beetle in Infested Ponderosa and Lodgepole Pine Logs; Research Paper, RMRS-RP-30; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2001; p. 11.
- Safranyik, L.; Carroll, A.L. The biology and epidemiology of the mountain pine beetle in lodgepole pine forests. In The Mountain Pine Beetle A Synthesis of Biology, Management, and Impacts on Lodgepole Pine; Safranyik, L., Wilson, B., Eds.; Natural Resources Canada, Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 2006; pp. 3–66. [Google Scholar]
- Schmitz, R.F.; McGregor, M.D.; Amman, G.D.; Oakes, R.D. Effect of partial cutting treatments of lodgepole pine stands on the abundance and behavior of flying mountain pine beetles. Can. J. For. Res. 1989, 19, 566–574. [Google Scholar] [CrossRef]
- Hindmarch, T.D.; Reid, M.L. Thinning of mature lodgepole pine stands increases scolytid bark beetle abundance and diversity. Can. J. For. Res. 2001, 31, 1502–1512. [Google Scholar] [CrossRef]
- Aukema, B.H.; Carroll, A.L.; Zhu, J.; Raffa, K.F.; Sickley, T.A.; Taylor, S.W. Landscape level analysis of mountain pine beetle in British Columbia, Canada: Spatiotemporal development and spatial synchrony within the present outbreak. Ecography 2006, 29, 427–441. [Google Scholar] [CrossRef]
- Aukema, B.H.; Carroll, A.L.; Zheng, Y.; Zhu, J.; Raffa, K.F.; Moore, D.R.; Stahl, K.; Taylor, S.W. Movement of outbreak populations of mountain pine beetle: Influences of spatiotemporal patterns and climate. Ecography 2008, 31, 348–358. [Google Scholar] [CrossRef]
- Chapman, T.B.; Veblen, T.T.; Schoennagel, T. Spatiotemporal patterns of mountain pine beetle activity in the southern Rocky Mountains. Ecology 2012, 93, 2175–2185. [Google Scholar] [CrossRef]
- Safranyik, L.; Silversides, R.; McMullen, L.H.; Linton, D.A. An empirical approach to modeling the local dispersal of the mountain pine beetle (Dendroctonus ponderosae Hopk.) (Col., Scolytidae) in relation to sources of attraction, wind direction, and speed. J. Appl. Entomol. 1989, 108, 498–511. [Google Scholar] [CrossRef]
- Safranyik, L.; Linton, D.A.; Silversides, R.; McMullen, L.H. Dispersal of released mountain pine beetles under the canopy of a mature lodgepole pine stand. J. Appl. Entomol. 1992, 113, 441–450. [Google Scholar] [CrossRef]
- Robertson, C.; Nelson, T.A.; Boots, B. Mountain pine beetle dispersal: The spatial–temporal interaction of infestations. For. Sci. 2007, 53, 395–405. [Google Scholar]
- Jackson, P.L.; Straussfogel, D.; Lindgren, B.S.; Mitchell, S.; Murphy, B. Radar observation of aerial capture of mountain pine beetle, Dendroctonus ponderosae Hopk. (Coleoptera: Scolytidae) in flight above the forest canopy. Can. J. For. Res. 2008, 38, 2313–2327. [Google Scholar] [CrossRef]
- Hynum, B.G.; Berryman, A.A. Dendroctunus ponderosae (Coleoptera: Scolytidae): Pre-aggregation landing and gallery initiation on lodgepole pine. Can. Entomol. 1980, 112, 185–191. [Google Scholar] [CrossRef]
- Campbell, S.A.; Borden, J.H. Integration of visual and olfactory cues of hosts and non-hosts by three bark beetles (Coleoptera: Scolytidae). Ecol. Entomol. 2006, 31, 437–449. [Google Scholar] [CrossRef]
- Pureswaran, D.S.; Borden, J.H. Primary attraction and kairomonal host discrimination in three species of Dendroctonus (Coleoptera: Scolytidae). Agric. For. Entomol. 2005, 7, 219–230. [Google Scholar] [CrossRef]
- Raffa, K.F.; Aukema, B.H.; Erbilgin, N.; Klepzig, K.D.; Wallin, K.F. Interactions among conifer terpenoids and bark beetles across multiple levels of scale: An attempt to understand links between population patterns and physiological processes. Recent Adv. Phytochem 2005, 39, 80–118. [Google Scholar]
- Raffa, K.F.; Berryman, A.A. Gustatory cues in the orientation of Dendroctonus ponderosae Coleoptera: Scolytidae) to host trees. Can. Entomol. 1982, 114, 97–104. [Google Scholar] [CrossRef]
- Edburg, S.L.; Allwine, G.; Lamb, B.; Stock, D.; Thistle, H.; Peterson, H.; Strom, B. A simple model to predict scalar dispersion within successively thinned loblolly pine canopy. J. Appl. Meteorol. Clim. 2010, 49, 1913–1926. [Google Scholar] [CrossRef]
- Fettig, C.J.; Klepzig, K.D.; Billings, R.F.; Munson, A.S.; Nebeker, T.E.; Negrón, J.F.; Nowak, J.T. The effectiveness of vegetation management practices for prevention and control of bark beetle infestations in coniferous forests of the western and southern United States. For. Ecol. Manag. 2007, 238, 24–53. [Google Scholar] [CrossRef]
- Raffa, K.F.; Berryman, A.A. The role of host plant resistance in the colonization behavior and ecology of bark beetles (Coleoptera: Scolytidae). Ecol. Monogr. 1983, 53, 27–49. [Google Scholar] [CrossRef]
- Raffa, K.F. Mixed messages across multiple trophic levels: The ecology of bark beetle chemical communication systems. Chemoecology 2001, 11, 49–65. [Google Scholar] [CrossRef]
- Berryman, A.A. Resistance to conifers to invasion by bark beetle-fungal associations. BioScience 1972, 22, 598–602. [Google Scholar] [CrossRef]
- Raffa, K.F.; Powell, E.N.; Townsend, P.A. Temperature-driven range expansion of an irruptive insect heightened by weakly coevolved plant defenses. Proc. Natl. Acad. Sci. USA. 2013, 110, 2193–2198. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, V.R.; Krokene, P.; Christiansen, E.; Krekling, T. Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytol. 2005, 167, 353–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolb, T.E.; Fettig, C.J.; Ayres, M.P.; Bentz, B.J.; Hicke, J.A.; Mathiasen, R.; Stewart, J.E.; Weed, A.S. Observed and anticipated impacts of drought on forest insects and diseases in the United States. For. Ecol. Manag. 2016, 380, 321–334. [Google Scholar] [CrossRef]
- Wood, D.L. The role of pheromones, kairomones, and allomones in the host selection and colonization behavior of bark beetles. Ann. Rev. Entomol. 1982, 27, 411–446. [Google Scholar] [CrossRef]
- Holtta, T.; Kurppa, M.; Nikinmaa, E. Scaling of xylem and phloem transport capacity and resource usage with tree size. Front. Plant Sci. 2013, 4, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Dannoura, M.; Epron, D.; Desalme, D.; Massonnet, C.; Tsuji, S.; Plain, C.; Priault, P.; Gérant, D. The impact of prolonged drought on phloem anatomy and phloem transport in young beech trees. Tree Physiol. 2018. [CrossRef] [PubMed]
- Sevanto, S. Drought impacts on phloem transport. Curr. Opin. Plant Biol. 2018, 43, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Cerezke, H.F. Egg gallery, brood production, and adult characteristics of mountain pine beetle, Dendroctonus ponderosae Hopkins (Coleoptera: Scolytidae), in three pine hosts. Can. Entomol. 1995, 127, 955–965. [Google Scholar] [CrossRef]
- Dean, D.K. Host utilization by the mountain pine beetle, Dendroctonus ponderosae Hopkins (Coleoptera: Scolytidae), in mixed stands of limber pine, Pinus flexilis James, and lodgepole pine, Pinus contorta latifolia Engelmann. Master’s Thesis, University of Wyoming, Laramie, WY, USA, 2007. [Google Scholar]
- Cole, W.E.; Cahill, D.B. Cutting strategies can reduce probabilities of mountain pine beetle epidemics in lodgepole pine. J. For. 1976, 74, 294–297. [Google Scholar]
- McGregor, M.D.; Amman, G.D.; Schmitz, R.F.; Oakes, R.D. Partial cutting lodgepole pine stands to reduce losses to the mountain pine beetle. Can. J. For. Res. 1987, 17, 1234–1239. [Google Scholar] [CrossRef]
- Mitchell, R.G.; Preisler, H.K. Analysis of spatial patterns of lodgepole pine attacked by outbreak populations of the mountain pine beetle. For. Sci. 1991, 37, 1390–1408. [Google Scholar]
- Negrón, J.F.; Allen, K.; Cook, B.; Withrow, J.R. Susceptibility of ponderosa pine, Pinus ponderosa (Dougl. ex Laws.), to mountain pine beetle, Dendroctonus ponderosae Hopkins, attack in uneven-aged stands in the Black Hills of South Dakota and Wyoming, USA. For. Ecol. Manag. 2008, 254, 327–334. [Google Scholar] [CrossRef]
- Powell, J.A.; Logan, J.A. Insect seasonality: Circle map analysis of temperature-driven life cycles. Theor. Popul. Biol. 2005, 67, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Amman, G.D. Mountain pine beetle (Coleptera: Scolytidae) mortality in three types of infestations. Environ. Entomol. 1984, 13, 184–191. [Google Scholar] [CrossRef]
- Mercado, J.E.; Hofstetter, R.W.; Reboletti, D.M.; Negrón, J.F. Phoretic symbionts of the mountain pine beetle (Dendroctonus ponderosae Hopkins). For. Sci. 2014, 60, 512–526. [Google Scholar] [CrossRef]
- Adams, A.S.; Aylward, F.O.; Adams, S.M.; Erbilgin, N.; Aukema, B.H.; Currie, C.R.; Suen, G.; Raffa, K.F. Mountain pine beetles colonizing historical and naïve host trees are associated with a bacterial community highly enriched in genes contributing to terpene metabolism. Appl. Environ. Micr. 2013, 79, 3468–3475. [Google Scholar] [CrossRef]
- Hofstetter, R.W.; Chen, Z.; Gaylord, M.L.; McMillin, J.D.; Wagner, M.R. Synergistic effects of a-pinene and exo-brevicomin on pine bark beetles and associated insects in Arizona. J. Appl. Entomol. 2008, 132, 387–397. [Google Scholar] [CrossRef]
- Klutsch, J.G.; Battaglia, M.A.; West, D.R.; Costello, S.L.; Negrón, J.F. Evaluating potential fire behavior in lodgepole pine-dominated forests after a mountain pine beetle epidemic in north-central Colorado. West. Appl. For. 2011, 26, 101–109. [Google Scholar]
- Kulakowski, D.; Jarvis, D.; Veblen, T.T.; Smith, J. Stand-replacing fires reduce susceptibility of lodgepole pine to mountain pine beetle outbreaks in Colorado. J. Biogeogr. 2012, 39, 2052–2060. [Google Scholar] [CrossRef]



| GSL | Mean (SE) Number of Beetles | Year | Mean (SE) Number of Beetles |
|---|---|---|---|
| 120 | 453.7 (139.2) a | 2005 | 127.0 (45.5) b |
| 80 | 235.4 (79.9) a | 2006 | 550.4 (140.5) a |
| 40 | 92.8 (28.0) b | 2007 | 104.5 (21.4) b |
| GSL 40 | GSL 120 | |
|---|---|---|
| Attacks per m2 | 101.1 (6.8) a | 73.6 (5.3) b |
| Emerging beetles per m2 | 638.4 (92.1) a | 556.8 (68.2) a |
| dbh (cm) | 28.1 (0.5) a | 26.2 (0.5) b |
| Percent phloem consumption 1 | 49.0 (4.7) a | 41.6 (2.4) a |
| GSL | Diameter at Breast Height (cm) | ||||
|---|---|---|---|---|---|
| 15 cm | 20 cm | 25 cm | 30 cm | 35 cm | |
| 40 vs. 80 | 0.003 * | 0.063 | 0.774 | <0.0001 * | <0.0001 * |
| 40 vs. 100 | 0.006 * | 0.542 | 0.004 * | <0.0001 * | <0.0001 * |
| 40 vs. 120 | 0.007 * | 0.483 | 0.012 * | <0.0001 * | <0.0001 * |
| 80 vs. 100 | 0.869 | 0.020 * | 0.004 * | 0.119 | 0.294 |
| 80 vs. 120 | 0.699 | 0.028 * | 0.013 * | 0.264 | 0.534 |
| 100 vs. 120 | 0.977 | 0.987 | 0.907 | 0.911 | 0.914 |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Negrón, J.F. Biological Aspects of Mountain Pine Beetle in Lodgepole Pine Stands of Different Densities in Colorado, USA. Forests 2019, 10, 18. https://doi.org/10.3390/f10010018
Negrón JF. Biological Aspects of Mountain Pine Beetle in Lodgepole Pine Stands of Different Densities in Colorado, USA. Forests. 2019; 10(1):18. https://doi.org/10.3390/f10010018
Chicago/Turabian StyleNegrón, José F. 2019. "Biological Aspects of Mountain Pine Beetle in Lodgepole Pine Stands of Different Densities in Colorado, USA" Forests 10, no. 1: 18. https://doi.org/10.3390/f10010018
APA StyleNegrón, J. F. (2019). Biological Aspects of Mountain Pine Beetle in Lodgepole Pine Stands of Different Densities in Colorado, USA. Forests, 10(1), 18. https://doi.org/10.3390/f10010018