Context-Dependence of Urban Forest Vegetation Invasion Level and Alien Species’ Ecological Success


Abstract
:1. Introduction
2. Materials and Methods
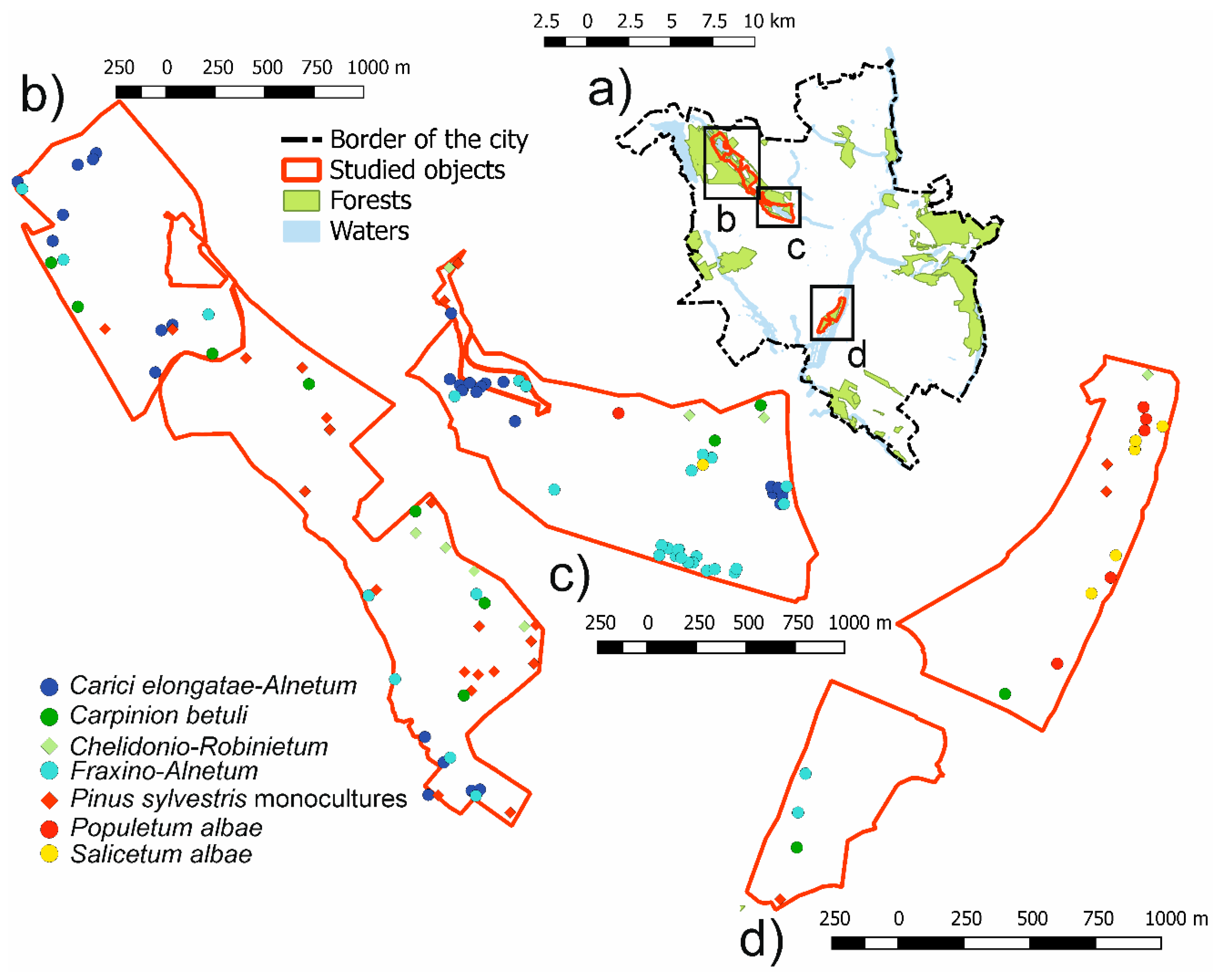
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
3. Results
3.1. Predictors of Alien Species Richness
3.2. Predictors of Alien Species Cover
3.3. Differences among Vegetation Types
4. Discussion
4.1. Patterns of Alien Species Invasions in Urban Forests
4.2. Resource Availability and Habitat Filtering
4.3. Rich Does Not Get Richer?
4.4. Impact of Disturbance
4.5. Differences among Vegetation Types
4.6. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of Earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.L.; Bacher, S.; Essl, F.; Hulme, P.E.; Jeschke, J.M.; Kühn, I.; Kumschick, S.; Nentwig, W.; Pergl, J.; Pyšek, P.; et al. Framework and guidelines for implementing the proposed IUCN Environmental Impact Classification for Alien Taxa (EICAT). Divers. Distrib. 2015, 21, 1360–1363. [Google Scholar] [CrossRef] [Green Version]
- Kowarik, I. Novel urban ecosystems, biodiversity, and conservation. Environ. Pollut. 2011, 159, 1974–1983. [Google Scholar] [CrossRef] [PubMed]
- Lososová, Z.; Chytrý, M.; Danihelka, J.; Tichý, L.; Ricotta, C. Biotic homogenization of urban floras by alien species: The role of species turnover and richness differences. J. Veg. Sci. 2016, 27, 452–459. [Google Scholar] [CrossRef]
- Chocholoušková, Z.; Pyšek, P. Changes in composition and structure of urban flora over 120 years: A case study of the city of Plzeň. Flora 2003, 198, 366–376. [Google Scholar] [CrossRef]
- Potgieter, L.J.; Gaertner, M.; Kueffer, C.; Larson, B.M.H.; Livingstone, S.W.; O’Farrell, P.J.; Richardson, D.M. Alien plants as mediators of ecosystem services and disservices in urban systems: A global review. Biol. Invasions 2017, 19, 3571–3588. [Google Scholar] [CrossRef]
- Godefroid, S.; Koedam, N. How important are large vs. small forest remnants for the conservation of the woodland flora in an urban context? Glob. Ecol. Biogeogr. 2003, 12, 287–298. [Google Scholar] [CrossRef]
- Kowarik, I. On the role of alien species in urban flora and vegetation. In Plant Invasions: General Aspects and Special Problems; Pyšek, P., Prach, K., Rejmánek, M., Wade, M., Eds.; SPB Academic Publishing: Amsterdam, Netherlands, 1995; pp. 85–103. [Google Scholar]
- Dyderski, M.K.; Gdula, A.K.; Jagodziński, A.M. “The rich get richer” concept in riparian woody species—A case study of the Warta River Valley (Poznań, Poland). Urban For. Urban Green. 2015, 14, 107–114. [Google Scholar] [CrossRef]
- Knapp, S.; Kühn, I.; Stolle, J.; Klotz, S. Changes in the functional composition of a Central European urban flora over three centuries. Perspect. Plant Ecol. Evol. Syst. 2010, 12, 235–244. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Tyborski, J.; Jagodziński, A.M. The utility of ancient forest indicator species in urban environments: A case study from Poznań, Poland. Urban For. Urban Green. 2017, 27, 76–83. [Google Scholar] [CrossRef]
- Pergl, J.; Sádlo, J.; Petřík, P.; Danihelka, J.; Chrtek, J., Jr.; Hejda, M.; Moravcova, L.; Perglová, I.; Štajerová, K.; Pyšek, P. Dark side of the fence: Ornamental plants as a source of wild-growing flora in the Czech Republic. Preslia 2016, 88, 163–184. [Google Scholar]
- Haeuser, E.; Dawson, W.; Thuiller, W.; Dullinger, S.; Block, S.; Bossdorf, O.; Carboni, M.; Conti, L.; Dullinger, I.; Essl, F.; et al. European ornamental garden flora as an invasion debt under climate change. J. Appl. Ecol. 2018, 55, 2386–2395. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Carlton, J.T. A compendium of esssential concepts and terminology in invasion ecology. In Fifty Years of Invasion Ecology: The Legacy of Charles Elton; Richardson, D.M., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2011; pp. 409–420. [Google Scholar]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Wilson, J.R.U.; Richardson, D.M. A proposed unified framework for biological invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeschke, J.M. General hypotheses in invasion ecology. Divers. Distrib. 2014, 20, 1129–1134. [Google Scholar] [CrossRef]
- Davis, M.A.; Thompson, K.; Philip Grime, J. Invasibility: The local mechanism driving community assembly and species diversity. Ecography 2005, 28, 696–704. [Google Scholar] [CrossRef]
- Ricciardi, A.; Blackburn, T.M.; Carlton, J.T.; Dick, J.T.A.; Hulme, P.E.; Iacarella, J.C.; Jeschke, J.M.; Liebhold, A.M.; Lockwood, J.L.; MacIsaac, H.J.; et al. Invasion Science: A Horizon Scan of Emerging Challenges and Opportunities. Trends Ecol. Evol. 2017, 32, 464–474. [Google Scholar] [CrossRef] [Green Version]
- Pairon, M.; Jonard, M.; Jacquemart, A.-L. Modeling seed dispersal of black cherry, an invasive forest tree: How microsatellites may help? Can. J. For. Res. 2006, 36, 1385–1394. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Dyderski, M.K.; Rawlik, M.; Banaszczak, P. Plantation of coniferous trees modifies risk and size of Padus serotina (Ehrh.) Borkh. Invasion—Evidence from a Rogów Arboretum case study. For. Ecol. Manag. 2015, 357, 84–94. [Google Scholar] [CrossRef]
- Vanhellemont, M.; Verheyen, K.; Keersmaeker, L.; Vandekerkhove, K.; Hermy, M. Does Prunus serotina act as an aggressive invader in areas with a low propagule pressure? Biol. Invasions 2009, 11, 1451–1462. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Jagodziński, A.M. Drivers of invasive tree and shrub natural regeneration in temperate forests. Biol. Invasions 2018, 20, 2363–2379. [Google Scholar] [CrossRef] [Green Version]
- Alpert, P.; Bone, E.; Holzapfel, C. Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants. Perspect. Plant Ecol. Evol. Syst. 2000, 3, 52–66. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P. Plant invasions: Merging the concepts of species invasiveness and community invasibility. Prog. Phys. Geogr. 2006, 30, 409–431. [Google Scholar] [CrossRef]
- Chytrý, M.; Jarošík, V.; Pyšek, P.; Hájek, O.; Knollová, I.; Tichý, L.; Danihelka, J. Separating Habitat Invasibility by Alien Plants from the Actual Level of Invasion. Ecology 2008, 89, 1541–1553. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Chytrỳ, M.; Jarošík, V. Habitats and land-use as determinants of plant invasions in the temperate zone of Europe. In Bioinvasions and Globalization: Ecology, Economics, Management and Policy; Perrings, C., Mooney, H., Williamson, M., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 66–79. [Google Scholar]
- Annighöfer, P.; Mölder, I.; Zerbe, S.; Kawaletz, H.; Terwei, A.; Ammer, C. Biomass functions for the two alien tree species Prunus serotina Ehrh. and Robinia pseudoacacia L. in floodplain forests of Northern Italy. Eur. J. For. Res. 2012, 131, 1619–1635. [Google Scholar] [CrossRef]
- Franzese, J.; Urrutia, J.; García, R.A.; Taylor, K.; Pauchard, A. Pine invasion impacts on plant diversity in Patagonia: Invader size and invaded habitat matter. Biol. Invasions 2017, 19, 1015–1027. [Google Scholar] [CrossRef]
- Staska, B.; Essl, F.; Samimi, C. Density and age of invasive Robinia pseudoacacia modulate its impact on floodplain forests. Basic Appl. Ecol. 2014, 15, 551–558. [Google Scholar] [CrossRef]
- Riepšas, E.; Straigyté, L. Invasiveness and Ecological Effects of Red Oak (Quercus rubra L.) in Lithuanian Forests. Balt. For. 2008, 14, 122–130. [Google Scholar]
- Wagner, V.; Chytrý, M.; Jiménez-Alfaro, B.; Pergl, J.; Hennekens, S.; Biurrun, I.; Knollová, I.; Berg, C.; Vassilev, K.; Rodwell, J.S.; et al. Alien plant invasions in European woodlands. Divers. Distrib. 2017, 23, 969–981. [Google Scholar] [CrossRef] [Green Version]
- Chmura, D. Penetration and naturalisation of invasive alien plant species (neophytes) in woodlands of the Silesian Upland (southern Poland). Nat. Conserv. 2004, 60, 3–11. [Google Scholar]
- Catford, J.A.; Downes, B.J.; Gippel, C.J.; Vesk, P.A. Flow regulation reduces native plant cover and facilitates exotic invasion in riparian wetlands. J. Appl. Ecol. 2011, 48, 432–442. [Google Scholar] [CrossRef] [Green Version]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; University of Chicago Press: Chicago, IL, USA, 1958. [Google Scholar]
- Catford, J.A.; Jansson, R.; Nilsson, C. Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers. Distrib. 2009, 15, 22–40. [Google Scholar] [CrossRef] [Green Version]
- Knight, K.S.; Oleksyn, J.; Jagodziński, A.M.; Reich, P.B.; Kasprowicz, M. Overstorey tree species regulate colonization by native and exotic plants: A source of positive relationships between understorey diversity and invasibility. Divers. Distrib. 2008, 14, 666–675. [Google Scholar] [CrossRef]
- Baker, H.G. The Evolution of Weeds. Ann. Rev. Ecol. Syst. 1974, 5, 1–24. [Google Scholar] [CrossRef]
- Pyšek, P.; Manceur, A.M.; Alba, C.; McGregor, K.F.; Pergl, J.; Štajerová, K.; Chytrý, M.; Danihelka, J.; Kartesz, J.; Klimešová, J.; et al. Naturalization of central European plants in North America: Species traits, habitats, propagule pressure, residence time. Ecology 2015, 96, 762–774. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Hulme, P.E.; Pyšek, P.; Jarošík, V.; Pergl, J.; Schaffner, U.; Vilà, M. Bias and error in understanding plant invasion impacts. Trends Ecol. Evol. 2013, 28, 212–218. [Google Scholar] [CrossRef]
- González-Moreno, P.; Diez, J.M.; Ibáñez, I.; Font, X.; Vilà, M. Plant invasions are context-dependent: Multiscale effects of climate, human activity and habitat. Divers. Distrib. 2014, 20, 720–731. [Google Scholar] [CrossRef]
- Jarošík, V.; Pyšek, P.; Kadlec, T. Alien plants in urban nature reserves: From red-list species to future invaders? NeoBiota 2011, 10, 27–46. [Google Scholar]
- Iannone, B.V.; Potter, K.M.; Hamil, K.-A.D.; Huang, W.; Zhang, H.; Guo, Q.; Oswalt, C.M.; Woodall, C.W.; Fei, S. Evidence of biotic resistance to invasions in forests of the Eastern USA. Landsc. Ecol. 2016, 31, 85–99. [Google Scholar] [CrossRef]
- Gaertner, M.; Wilson, J.R.U.; Cadotte, M.W.; MacIvor, J.S.; Zenni, R.D.; Richardson, D.M. Non-native species in urban environments: Patterns, processes, impacts and challenges. Biol. Invasions 2017, 19, 3461–3469. [Google Scholar] [CrossRef]
- Simberloff, D.; Von Holle, B. Positive interactions of nonindigenous species: Invasional meltdown? Biol. Invasions 1999, 1, 21–32. [Google Scholar] [CrossRef]
- Stohlgren, T.J.; Jarnevich, C.; Chong, G.W.; Evangelista, P.H.; Pyšek, P.; Kaplan, Z.; Richardson, D.M. Scale and plant invasions: A theory of biotic acceptance. Preslia 2006, 78, 405–426. [Google Scholar]
- Darwin, C. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859; ISBN 978-83-235-0531-0. [Google Scholar]
- Melbourne, B.A.; Cornell, H.V.; Davies, K.F.; Dugaw, C.J.; Elmendorf, S.; Freestone, A.L.; Hall, R.J.; Harrison, S.; Hastings, A.; Holland, M.; et al. Invasion in a heterogeneous world: Resistance, coexistence or hostile takeover? Ecol. Lett. 2007, 10, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Hierro, J.L.; Maron, J.L.; Callaway, R.M. A biogeographical approach to plant invasions: The importance of studying exotics in their introduced and native range. J. Ecol. 2005, 93, 5–15. [Google Scholar] [CrossRef]
- Sher, A.A.; Hyatt, L.A. The Disturbed Resource-Flux Invasion Matrix: A New Framework for Patterns of Plant Invasion. Biol. Invasions 1999, 1, 107–114. [Google Scholar] [CrossRef]
- Davis, M.A.; Grime, J.P.; Thompson, K. Fluctuating resources in plant communities: A general theory of invasibility. J. Ecol. 2000, 88, 528–534. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Huenneke, L.F. Disturbance, Diversity, and Invasion: Implications for Conservation. Conserv. Biol. 1992, 6, 324–337. [Google Scholar] [CrossRef]
- Jackowiak, B. Poznań. In Plants and Habitats of European Cities; Müller, N., Kelcey, J.G., Eds.; Springer: New York, NY, USA, 2011; pp. 363–405. ISBN 978-0-387-89683-0. [Google Scholar]
- Statistical Office in Poznań. Statistical Yearbook of Poznań City; Statistical Office in Poznań: Poznań, Poland, 2013.
- Dyderski, M.K.; Wrońska-Pilarek, D.; Jagodziński, A.M. Ecological lands for conservation of vascular plant diversity in the urban environment. Urban Ecosyst. 2016, 20, 639–650. [Google Scholar] [CrossRef] [Green Version]
- Dyderski, M.K.; Wrońska-Pilarek, D. Szata roślinna nowo powstałego użytku ekologicznego “Strzeszyn” w Poznaniu i stan jej zachowania. Nauk. Przyr. Technol. 2015, 9, 39. [Google Scholar]
- Dyderski, M.K.; Gdula, A.K.; Wrońska-Pilarek, D. Roślinność nowo utworzonych użytków ekologicznych Bogdanka I i Bogdanka II w Poznaniu. Acta Scientiarum Polonorum Silvarum Colendarum Ratio et Industria Lignaria 2014, 13, 22–37. [Google Scholar]
- Dyderski, M.K.; Wrońska-Pilarek, D. Szata roślinna nowo powstałych użytków ekologicznych “Dębina I” i “Dębina II” w Poznaniu. Nauk. Przyr. Technol. 2015, 9, 46. [Google Scholar]
- Dyderski, M.K.; Gdula, A.K.; Wrońska-Pilarek, D. Aktualny stan szaty roślinnej terenów wokół jeziora Rusałka w Poznaniu w warunkach antropopresji. Acta Scientiarum Polonorum Silvarum Colendarum Ratio et Industria Lignaria 2016, 15, 229–246. [Google Scholar] [CrossRef]
- Kącki, Z.; Śliwiński, M. The Polish Vegetation Database: Structure, resources and development. Acta Soc. Bot. Pol. 2012, 81, 75–79. [Google Scholar] [CrossRef]
- Tremblay, N.O.; Larocque, G.R. Seasonal dynamics of understory vegetation in four eastern Canadian forest types. Int. J. Plant Sci. 2001, 162, 271–286. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Dyderski, M.K.; Rawlik, K.; Kątna, B. Seasonal variability of biomass, total leaf area and specific leaf area of forest understory herbs reflects their life strategies. For. Ecol. Manag. 2016, 374, 71–81. [Google Scholar] [CrossRef]
- Czapiewska, N.; Paź, S.; Dyderski, M.K.; Jagodziński, A.M. Ciągłość kompozycji florystycznej pomiędzy dwoma zbiorowiskami roślinnymi—Carici elongatae-Alnetum oraz Fraxino-Alnetum. Leśn. Pr. Badaw. 2017, 78, 285–296. [Google Scholar]
- Holeksa, J.; Woźniak, G. Biased vegetation patterns and detection of vegetation changes using phytosociological databases. A case study in the forests of the Babia Góra National Park (the West Carpathians, Poland). Phytocoenologia 2005, 35, 1–18. [Google Scholar] [CrossRef]
- Lepš, J.; Hadincová, V. How reliable are our vegetation analyses? J. Veg. Sci. 1992, 3, 119–124. [Google Scholar] [CrossRef]
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Flowering Plants and Pteridophytes of Poland. A Checklist; W. Szafer Institute of Botany, PAN: Kraków, Poland, 2003. [Google Scholar]
- Tokarska-Guzik, B.; Dajdok, Z.; Zając, M.; Zając, A.; Urbisz, A.; Danielewicz, W.; Hołdyński, C. Rośliny Obcego Pochodzenia w Polsce ze Szczególnym Uwzględnieniem Gatunków Inwazyjnych; Generalna Dyrekcja Ochrony Środowiska: Warszawa, Poland, 2012; ISBN 978-83-62940-33-2.
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity (FD) from Multiple Traits, and Other tools for Functional Ecology; 2014. Available online: https://cran.r-project.org/web/packages/FD/index.html (accessed on 28 December 2018).
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas Mit Den Alpen in Ökologischer, Dynamischer und Historischer Sicht; UTB: Stuttgart, Germany, 2010. [Google Scholar]
- Klotz, S.; Kühn, I.; Durka, W. BIOLFLOR—Eine Datenbank zu Biologisch-Ökologischen Merkmalen der Gefäßpflanzen in Deutschland. Schriftenreihe für Vegetationskunde; Schriftenreihe für Vegetationskunde; Bundesamt für Naturschutz: Bonn, Germany, 2002. [Google Scholar]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; Van Groenendael, J.M.; Klimeš, L.; Klimešová, J.; et al. The LEDA Traitbase: A database of life-history traits of the Northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Czortek, P.; Delimat, A.; Dyderski, M.K.; Zięba, A.; Jagodziński, A.M.; Jaroszewicz, B. Climate change, tourism and historical grazing influence the distribution of Carex lachenalii Schkuhr—A rare arctic-alpine species in the Tatra Mts. Sci. Total Environ. 2018, 618, 1628–1637. [Google Scholar] [CrossRef] [PubMed]
- Herben, T.; Chytrý, M.; Klimešová, J. A quest for species-level indicator values for disturbance. J. Veg. Sci. 2016, 27, 628–636. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Biecek, P. DALEX: Descriptive mAchine Learning EXplanations. 2018. Available online: https://cran.r-project.org/web/packages/DALEX/index.html (accessed on 28 December 2018).
- Biecek, P. CeterisParibus: Ceteris Paribus Plots (What-If Plots) for a Single Observation. 2018. Available online: https://cran.r-project.org/web/packages/ceterisParibus/index.html (accessed on 28 December 2018).
- Rigby, R.A.; Stasinopoulos, D.M. Generalized additive models for location, scale and shape. J. R. Stat. Soc. C (Appl. Stat.) 2005, 54, 507–554. [Google Scholar] [CrossRef]
- Sitzia, T.; Campagnaro, T.; Weir, R.G. Novel woodland patches in a small historical Mediterranean city: Padova, Northern Italy. Urban Ecosyst. 2016, 19, 475–487. [Google Scholar] [CrossRef]
- Marozas, V.; Cekstere, G.; Laivins, M.; Straigyte, L. Comparison of neophyte communities of Robinia pseudoacacia L. and Acer negundo L. in the eastern Baltic Sea region cities of Riga and Kaunas. Urban For. Urban Green. 2015, 14, 826–834. [Google Scholar] [CrossRef]
- Pyšek, P. Alien and native species in Central European urban floras: A quantitative comparison. J. Biogeogr. 1998, 25, 155–163. [Google Scholar] [CrossRef]
- Kowarik, I.; Lippe, M.; Cierjacks, A. Prevalence of alien versus native species of woody plants in Berlin differs between habitats and at different scales. Preslia 2013, 85, 113–132. [Google Scholar]
- Zerbe, S.; Wirth, P. Non-indigenous plant species and their ecological range in Central European pine (Pinus sylvestris L.) forests. Ann. For. Sci. 2006, 63, 189–203. [Google Scholar] [CrossRef]
- Lambdon, P.W.; Pyšek, P.; Basnou, C.; Hejda, M.; Arianoutsou, M.; Essl, F.; Jarošík, V.; Pergl, J.; Winter, M.; Anastasiu, P.; et al. Alien flora of Europe: Species diversity, temporal trends, geographical patterns and research needs. Preslia 2008, 80, 101–149. [Google Scholar]
- Pierce, S.; Brusa, G.; Vagge, I.; Cerabolini, B.E.L. Allocating CSR plant functional types: The use of leaf economics and size traits to classify woody and herbaceous vascular plants. Funct. Ecol. 2013, 27, 1002–1010. [Google Scholar] [CrossRef]
- Pyšek, P.; Křivánek, M.; Jarošík, V. Planting intensity, residence time, and species traits determine invasion success of alien woody species. Ecology 2009, 90, 2734–2744. [Google Scholar] [CrossRef]
- Dylewski, Ł.; Maćkowiak, Ł.; Myczko, Ł. Physical defence of the wild cucumber Echinocystis lobata in an invasive range changing seed removal by rodents. Plant Ecol. 2018, 219, 863–873. [Google Scholar] [CrossRef]
- Chmura, D. Biology and Ecology of an Invasion of Impatiens Parviflora DC in Natural and Semi-Natural Habitats; Wydawnictwo ATH: Bielsko-Biała, Poland, 2014. [Google Scholar]
- Godefroid, S.; Koedam, N. Comparative ecology and coexistence of introduced and native congeneric forest herbs: Impatiens parviflora and I. noli-tangere. Plant Ecol. Evol. 2010, 143, 119–127. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; Pauchard, A.; Lenoir, J.; Nuñez, M.A.; Geron, C.; Ven, A.; Bravo-Monasterio, P.; Teneb, E.; Nijs, I.; Milbau, A. Disturbance is the key to plant invasions in cold environments. Proc. Natl. Acad. Sci. USA 2016, 113, 14061–14066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyderski, M.K.; Czapiewska, N.; Zajdler, M.; Tyborski, J.; Jagodziński, A.M. Functional diversity, succession, and human-mediated disturbances in raised bog vegetation. Sci. Total Environ. 2016, 562, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Pauchard, A.; Kueffer, C.; Dietz, H.; Daehler, C.C.; Alexander, J.; Edwards, P.J.; Arévalo, J.R.; Cavieres, L.A.; Guisan, A.; Haider, S.; et al. Ain’t no mountain high enough: Plant invasions reaching new elevations. Front. Ecol. Environ. 2009, 7, 479–486. [Google Scholar] [CrossRef]
- Parker, J.D.; Richie, L.J.; Lind, E.M.; Maloney, K.O. Land use history alters the relationship between native and exotic plants: The rich don’t always get richer. Biol. Invasions 2010, 12, 1557–1571. [Google Scholar] [CrossRef]
- Herben, T.; Mandák, B.; Bímová, K.; Münzbergová, Z. Invasibility and species richness of a community: A neutral model and a survey of published data. Ecology 2004, 85, 3223–3233. [Google Scholar] [CrossRef]
- Davies, K.F.; Chesson, P.; Harrison, S.; Inouye, B.D.; Melbourne, B.A.; Rice, K.J. Spatial heterogeneity explains the scale dependence of the native-exotic diversity relationship. Ecology 2005, 86, 1602–1610. [Google Scholar] [CrossRef]
- Fox, J.F. Intermediate-Disturbance Hypothesis. Science 1979, 204, 1344–1345. [Google Scholar] [CrossRef] [Green Version]
- Chmura, D.; Sierka, E. The invasibility of deciduous forest communities after disturbance: A case study of Carex brizoides and Impatiens parviflora invasion. For. Ecol. Manag. 2007, 242, 487–495. [Google Scholar] [CrossRef]
- Chabrerie, O.; Verheyen, K.; Saguez, R.; Decocq, G. Disentangling relationships between habitat conditions, disturbance history, plant diversity, and American black cherry (Prunus serotina Ehrh.) invasion in a European temperate forest. Divers. Distrib. 2008, 14, 204–212. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Jagodziński, A.M. Low impact of disturbance on ecological success of invasive tree and shrub species in temperate forests. Plant Ecol. 2018, 219, 1369–1380. [Google Scholar] [CrossRef]
- Gunderson, L.H. Ecological Resilience—In Theory and Application. Ann. Rev. Ecol. Syst. 2000, 31, 425–439. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Dyderski, M.K.; Horodecki, P.; Rawlik, K. Limited dispersal prevents Quercus rubra invasion in a 14-species common garden experiment. Divers. Distrib. 2018, 24, 403–414. [Google Scholar] [CrossRef]
- Medvecká, J.; Kliment, J.; Majekova, J.; Halada, L.; Zaliberova, M.; Gojdičová, E.; Ferakova, V.; Jarolimek, I. Inventory of the alien flora of Slovakia. Preslia 2012, 84, 257–309. [Google Scholar]
- Pyšek, P.; Danihelka, J.; Sádlo, J.; Chrtek, J.; Chytrỳ, M.; Jarošík, V.; Kaplan, Z.; Krahulec, F.; Moravcová, L.; Pergl, J.; et al. Catalogue of alien plants of the Czech Republic: Checklist update, taxonomic diversity and invasion patterns. Preslia 2012, 84, 155–255. [Google Scholar]
- Wulf, M. Preference of plant species for woodlands with differing habitat continuities. Flora 2003, 198, 444–460. [Google Scholar] [CrossRef]
- Niinemets, Ü. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol. Res. 2010, 25, 693–714. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J.; Modrzynski, J.; Mrozinski, P.; Hobbie, S.E.; Eissenstat, D.M.; Chorover, J.; Chadwick, O.A.; Hale, C.M.; Tjoelker, M.G. Linking litter calcium, earthworms and soil properties: A common garden test with 14 tree species. Ecol. Lett. 2005, 8, 811–818. [Google Scholar] [CrossRef]
- Horodecki, P.; Jagodziński, A.M. Tree species effects on litter decomposition in pure stands on afforested post-mining sites. For. Ecol. Manag. 2017, 406, 1–11. [Google Scholar] [CrossRef]
- Dickie, I.A.; Oleksyn, J.; Reich, P.B.; Karolewski, P.; Zytkowiak, R.; Jagodziński, A.M.; Turzanska, E. Soil modification by different tree species influences the extent of seedling ectomycorrhizal infection. Mycorrhiza 2006, 16, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Mueller, K.E.; Eisenhauer, N.; Reich, P.B.; Hobbie, S.E.; Chadwick, O.A.; Chorover, J.; Dobies, T.; Hale, C.M.; Jagodziński, A.M.; Kałucka, I.; et al. Light, earthworms, and soil resources as predictors of diversity of 10 soil invertebrate groups across monocultures of 14 tree species. Soil Biol. Biochem. 2016, 92, 184–198. [Google Scholar] [CrossRef] [Green Version]
- Barbier, S.; Gosselin, F.; Balandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved—A critical review for temperate and boreal forests. For. Ecol. Manag. 2008, 254, 1–15. [Google Scholar] [CrossRef]
- Augusto, L.; Dupouey, J.-L.; Ranger, J. Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann. For. Sci. 2003, 60, 823–831. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Parameter Tested (Table 2) | Hypothesis | Explanation | Reference |
|---|---|---|---|
| Rich.nat | Biotic resistance | High species richness of native species prevents invasion of alien species, i.e., decreases level of invasion | [24,35,44] |
| Rich.nat | Biotic acceptance | High native species richness indicates habitats suitable to be colonized by both native and alien species, i.e., prone to invasion | [10,47] |
| FRic, FDis, EIV-N, EIV-L, EIV-M | Habitat filtering | Invaders may be successful only if their traits are adapted to existing environmental filters, i.e., high level of environmental filtering decreases community level of invasion | [48,49] |
| FRic, FDis | Empty niche | Due to limited pool of native species, plant communities are unsaturated and provide opportunities for functionally distinct alien species, i.e., increases level of invasion | [50] |
| FDis | Environmental heterogeneity | Higher variability of niches in plant communities provide opportunities for colonization by more alien species, i.e., increases level of invasion | [49,50] |
| EIV-N, EIV-L, EIV-M | Increased resources availability | Increase of resource availability increases level of invasion | [25,51,52] |
| Dist.freq, Dist.sev | Disturbance | Disturbance events increase resource availability and reduce competition for alien species, i.e., increase level of invasion | [51,53] |
| Vegetation Type | No. of Plots | Min Plot Area (m2) | Mean Plot Area (m2) | SE | Max Plot Area (m2) |
|---|---|---|---|---|---|
| Carici elongatae-Alnetum | 31 | 100 | 109.7 | 5.4 | 200 |
| Carpinion betuli | 10 | 100 | 140.0 | 16.3 | 200 |
| Chelidonio-Robinietum | 8 | 100 | 106.3 | 6.3 | 150 |
| Fraxino-Alentum | 35 | 100 | 115.7 | 5.7 | 200 |
| Pinus monocultures | 24 | 100 | 100.0 | 0.0 | 100 |
| Populetum albae | 6 | 80 | 95.0 | 3.4 | 100 |
| Salicetum albae | 6 | 80 | 110.0 | 10.3 | 150 |
| Parameter | Abbreviation | Details | Min | Max | Mean | SE |
|---|---|---|---|---|---|---|
| Plot area | Plot.area | Measurement of plot area (m2) | 80.0 | 200.0 | 111.1 | 2.8 |
| Alien species richness * | Rich.alien | Number of alien species | 0.0 | 5.0 | 2.1 | 0.1 |
| Cover of alien species * | Cov.alien | Sum of cover scores for each alien species | 0.0 | 77.0 | 11.5 | 1.0 |
| Disturbance frequency index | Dist.freq | Species cover weighted mean of Herben et al. [76] indicators | −0.899 | 0.137 | −0.591 | 0.024 |
| Disturbance severity index | Dist.sev | Species cover weighted mean of Herben et al. [67] indicators | 0.083 | 0.500 | 0.258 | 0.008 |
| Ecological indicator value—light | EIV-L | Species cover weighted mean of Ellenberg and Leuschner [71] indicators | 4.000 | 6.898 | 5.422 | 0.066 |
| Ecological indicator value—moisture | EIV-M | Species cover weighted mean of Ellenberg and Leuschner [71] indicators | 3.950 | 9.545 | 6.122 | 0.111 |
| Ecological indicator value—soil fertility | EIV-N | Species cover weighted mean of Ellenberg and Leuschner [71] indicators | 1.495 | 8.638 | 6.217 | 0.100 |
| Functional dispersion | FDis | Averaged distance to the centroid in functional traits hypervolume | 0.05955 | 0.24113 | 0.16500 | 0.00370 |
| Functional richness | FRic | The volume of plant community species convex hull in functional traits hypervolume | 0.00001 | 0.20894 | 0.02336 | 0.00278 |
| Native species richness | Rich.nat | Number of native species | 3.0 | 28.0 | 11.6 | 0.5 |
| Tree stand type | StType | Plant association name | (seven classes) | |||
| Vegetation Type | Mean Alien Species Richness | SE | Tukey Posteriori Test | Mean Alien Species Cover (%) | SE | Tukey Posteriori Test |
|---|---|---|---|---|---|---|
| Carici elongatae-Alnetum | 1.3 | 0.2 | a | 7.8 | 1.6 | a |
| Carpinion betuli | 2.5 | 0.3 | ab | 9.2 | 2.1 | c |
| Chelidonio-Robinietum | 3.6 | 0.4 | b | 14.8 | 2.6 | cd |
| Fraxino-Alentum | 2.2 | 0.2 | ab | 13.0 | 1.9 | e |
| Pinus monocultures | 2.3 | 0.2 | ab | 8.6 | 1.1 | b |
| Populetum albae | 2.5 | 0.5 | ab | 10.8 | 3.6 | abcd |
| Salicetum albae | 2.8 | 0.4 | ab | 34.5 | 9.8 | d |
| Species 1 | Life Form 2 | Invasion Status in Poland 3 [68] | Carici elongatae-Alnetum | Meadian Cover (%) | Carpinion betuli | Meadian Cover (%) | Chelidonio-Robinietum | Meadian Cover (%) | Fraxino-Alentum | Meadian Cover (%) | Pinus Monocultures | Meadian Cover (%) | Populetum albae | Meadian Cover (%) | Salicetum albae | Meadian Cover (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Acer negundo L. | P | inv | 0.0 | 0.0 | 10.0 | 2.0 | 25.0 | 1.5 | 5.7 | 2.0 | 4.2 | 2.0 | 16.7 | 2.0 | 66.7 | 3.0 |
| Amaranthus retroflexus L. | T | inv | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 2.0 |
| Bidens frondosa L. | T | inv | 9.7 | 1.0 | 0.0 | 0.0 | 0.0 | 0.0 | 11.4 | 2.0 | 0.0 | 0.0 | 16.7 | 2.0 | 66.7 | 10.5 |
| Chenopodium strictum Roth | T | nat | 0.0 | 0.0 | 0.0 | 0.0 | 12.5 | 1.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Cornus alba L. | P | cas | 3.2 | 3.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Cornus sericea L. | P | inv | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.7 | 3.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Echinocystis lobata (Michx.) Torr. & A. Gray | T, L | inv | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 3.0 | 0.0 | 0.0 |
| Fraxinus pensylvanica Marshall | P | inv | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 1.0 | 0.0 | 0.0 |
| Impatiens parviflora DC. | T | inv | 74.2 | 4.0 | 90.0 | 4.0 | 75.0 | 6.0 | 82.9 | 4.0 | 83.3 | 3.0 | 83.3 | 3.0 | 83.3 | 18.0 |
| Ligustrum vulgare L. * | P | nat | 6.5 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Lonicera tatarica L. | P | nat | 0.0 | 0.0 | 0.0 | 0.0 | 12.5 | 2.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Parthenocissus inserta (A. Kern.) R. M. Fritsch | P, L | inv | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.6 | 1.0 | 4.2 | 2.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Prunus domestica L. | P | nat | 6.5 | 1.0 | 40.0 | 1.0 | 25.0 | 3.5 | 5.7 | 1.5 | 12.5 | 1.0 | 16.7 | 4.0 | 16.7 | 18.0 |
| Prunus mahaleb L. | P | nat | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.2 | 1.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Prunus serotina Ehrh. | P | inv | 12.9 | 2.0 | 60.0 | 2.5 | 62.5 | 3.0 | 22.9 | 1.0 | 79.2 | 3.0 | 16.7 | 1.0 | 0.0 | 0.0 |
| Quercus rubra L. | P | inv | 0.0 | 0.0 | 0.0 | 0.0 | 37.5 | 1.0 | 0.0 | 0.0 | 8.3 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ribes alpinum L. * | P | nat | 0.0 | 0.0 | 20.0 | 3.5 | 50.0 | 8.0 | 2.9 | 8.0 | 16.7 | 8.0 | 16.7 | 8.0 | 0.0 | 0.0 |
| Ribes rubrum L. | P | nat | 12.9 | 2.5 | 10.0 | 2.0 | 0.0 | 0.0 | 42.9 | 4.0 | 0.0 | 0.0 | 33.3 | 6.0 | 16.7 | 18.0 |
| Ribes uva-crispa L. * | P | nat | 0.0 | 0.0 | 10.0 | 2.0 | 12.5 | 2.0 | 8.6 | 3.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 3.0 |
| Robinia pseudacacia L. | P | inv | 0.0 | 0.0 | 10.0 | 2.0 | 37.5 | 2.0 | 2.9 | 1.0 | 16.7 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 |
| Rumex thyrsiflorus Fingerh. | H | nat | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 2.0 | 0.0 | 0.0 |
| Salvia glutinosa L. * | H | cas | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.6 | 3.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Solidago canadensis L. | G, H | inv | 3.2 | 1.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.9 | 2.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Solidago gigantea Aiton | G, H | inv | 0.0 | 0.0 | 0.0 | 0.0 | 12.5 | 3.0 | 5.7 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dyderski, M.K.; Jagodziński, A.M. Context-Dependence of Urban Forest Vegetation Invasion Level and Alien Species’ Ecological Success. Forests 2019, 10, 26. https://doi.org/10.3390/f10010026
Dyderski MK, Jagodziński AM. Context-Dependence of Urban Forest Vegetation Invasion Level and Alien Species’ Ecological Success. Forests. 2019; 10(1):26. https://doi.org/10.3390/f10010026
Chicago/Turabian StyleDyderski, Marcin K., and Andrzej M. Jagodziński. 2019. "Context-Dependence of Urban Forest Vegetation Invasion Level and Alien Species’ Ecological Success" Forests 10, no. 1: 26. https://doi.org/10.3390/f10010026
APA StyleDyderski, M. K., & Jagodziński, A. M. (2019). Context-Dependence of Urban Forest Vegetation Invasion Level and Alien Species’ Ecological Success. Forests, 10(1), 26. https://doi.org/10.3390/f10010026