Relationships among Root–Shoot Ratio, Early Growth, and Health of Hybrid Poplar and Willow Clones Grown in Different Landfill Soils


Abstract
:1. Introduction
2. Materials and Methods
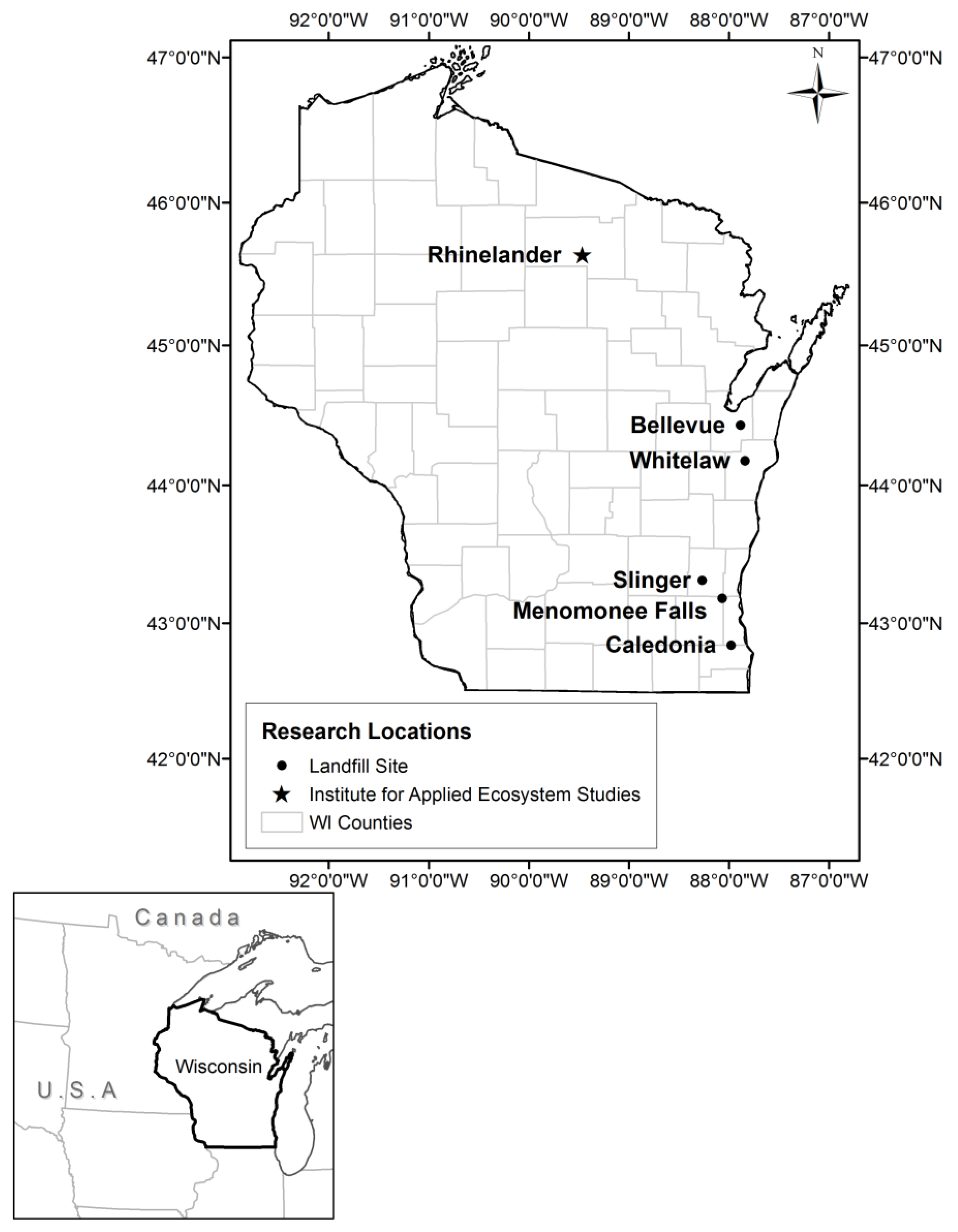
2.1. Soil Collection and Site Description
2.2. Soil Properties
2.3. Genotype Selection and Experimental Design
2.4. Weighted Summation Indices
2.5. Data Collection
2.6. Growth Index
2.7. Data Analysis
3. Results
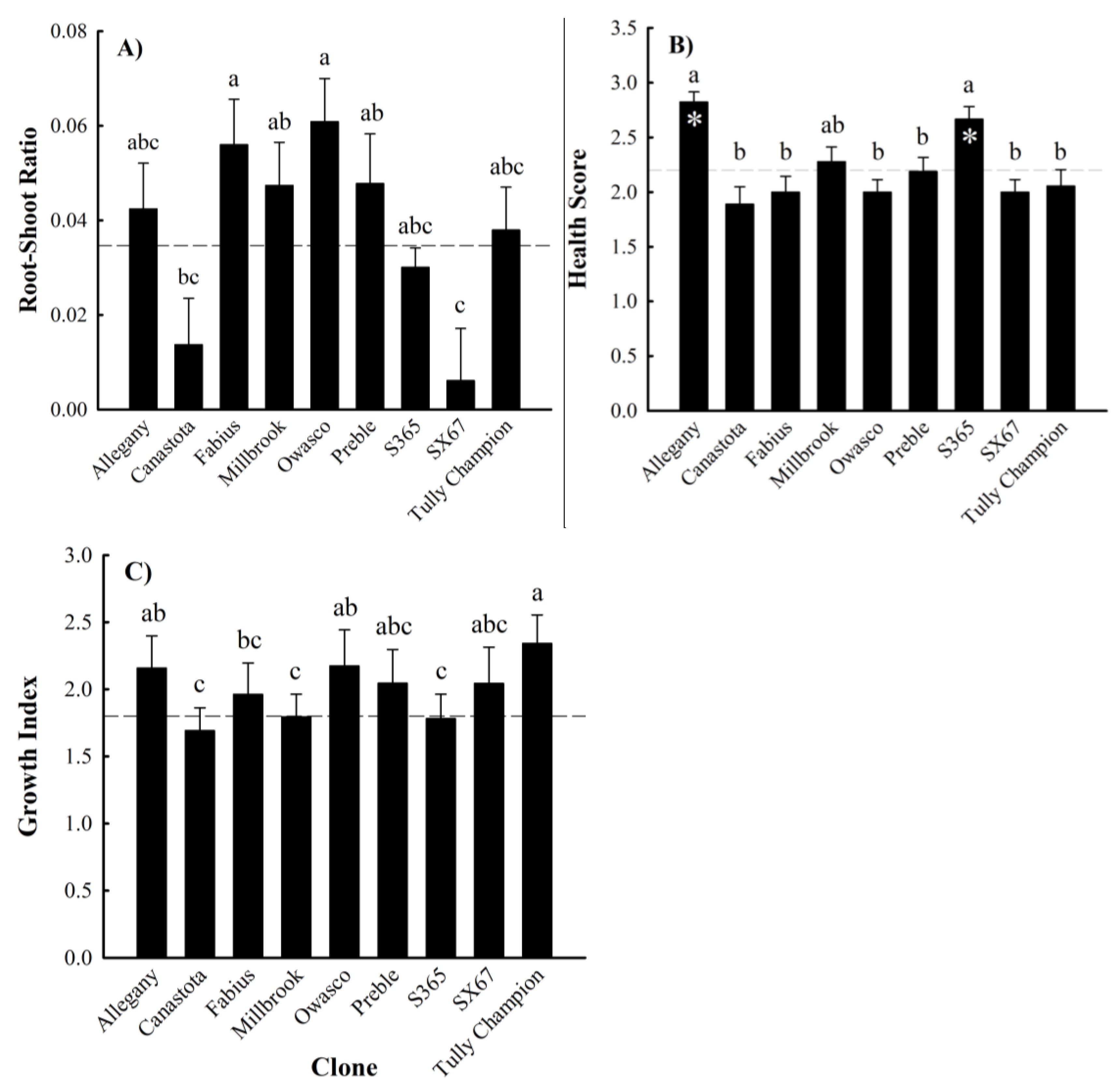
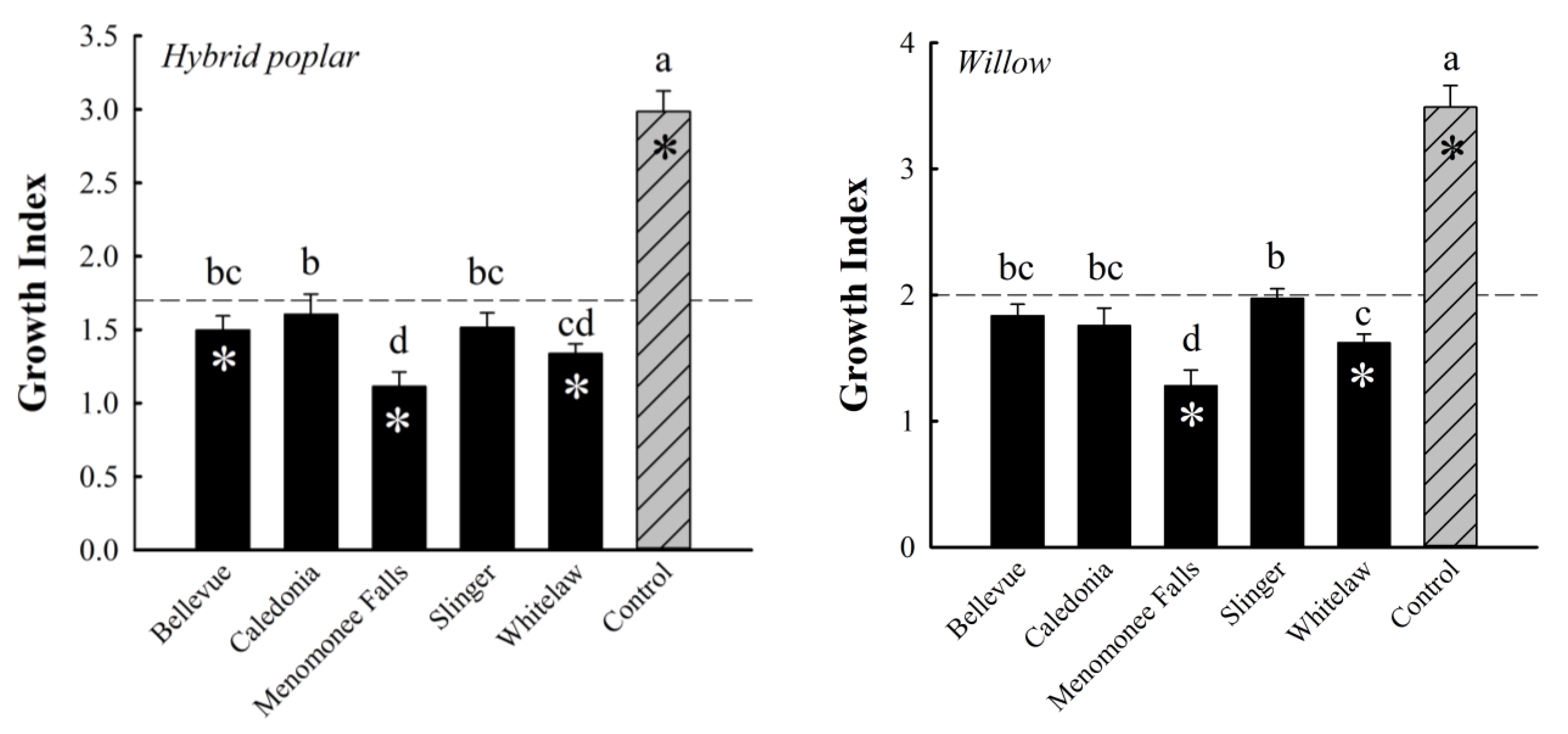
3.1. Analyses of Variance
3.2. Correlations
3.3. Analyses of Covariance
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schnoor, J.L.; Licht, L.A.; McCutcheon, S.C.; Wolfe, N.L.; Carreira, L.H. Phytoremediation of organic and nutrient contaminants. Environ. Sci. Technol. 1995, 29, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Suresh, B.; Ravishanker, G.A. Phytoremediation: A novel and promising approach for environmental clean-up. Crit. Rev. Biotechnol. 2004, 24, 97–124. [Google Scholar] [CrossRef]
- Robinson, B.H.; Green, S.R.; Mills, T.M.; Clothier, B.E.; van der Velde, M.; Laplane, R.; Fung, L.; Deurer, M.; Hurst, S.; Thayalakumaran, T.; et al. Phytoremediation: Using plants as biopumps to improve degraded environments. Aust. J. Soil Res. 2003, 41, 599–611. [Google Scholar] [CrossRef]
- Mirck, J.; Isebrands, J.G.; Verwijst, T.; Ledin, S. Development of short-rotation willow coppice systems for environmental purposes in Sweden. Biomass Bioenergy 2005, 28, 219–228. [Google Scholar] [CrossRef]
- Cunningham, S.D.; Shann, J.R.; Crowley, D.E.; Anderson, T.A. Phytoremediation of contaminated water and soil. In Phytoremediation of Soil and Water Contaminants; Kruger, E.L., Anderson, T.A., Coats, J.R., Eds.; American Chemical Society: Washington, DC, USA, 1997; pp. 2–17. ISBN 9780841235038. [Google Scholar]
- Cunningham, S.D.; Ow, D.W. Promises and prospects of phytoremediation. Plant Physiol. 1996, 110, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, J.A.; Zalesny, R.S., Jr.; Wiese, A.H.; Hall, R.B. Choosing tree genotypes for phytoremediation of landfill leachate using phyto-recurrent selection. Int. J. Phytorem. 2007, 9, 513–530. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, R.S., Jr.; Stanturf, J.A.; Gardiner, E.S.; Bañuelos, G.S.; Hallett, R.A.; Hass, A.; Stange, C.M.; Perdue, J.H.; Young, T.M.; Coyle, D.R.; et al. Environmental technologies of woody crop production systems. BioEnergy Res. 2016, 9, 492–506. [Google Scholar] [CrossRef]
- Zalesny, J.A.; Zalesny, R.S., Jr.; Coyle, D.R.; Hall, R.B.; Bauer, E.O. Clonal variation in morphology of Populus root systems following irrigation with landfill leachate or water during two years of establishment. BioEnergy Res. 2009, 2, 134–143. [Google Scholar] [CrossRef]
- Netzer, D.; Tolsted, D.; Ostry, M.E.; Isebrands, J.G.; Riemenschneider, D.; Ward, K. Growth, Yield, and Disease Resistance of 7- to 12-Year-Old Poplar Clones in the North Central United States; General Technical Report NC 229; U.S. Department of Agriculture, Forest Service, North Central Research Station: St. Paul, MN, USA, 2002; pp. 1–40. [Google Scholar]
- Zalesny, R.S., Jr.; Stanturf, J.A.; Gardiner, E.S.; Perdue, J.H.; Young, T.M.; Coyle, D.R.; Headlee, W.L.; Bañuelos, G.S.; Hass, A. Ecosystem services of woody crop production systems. BioEnergy Res. 2016, 9, 465–491. [Google Scholar] [CrossRef]
- Baldantoni, D.; Cicatelli, A.; Bellion, A.; Castiglione, S. Different behaviours in phytoremediation capacity of two heavy metal tolerant poplar clones in relation to iron and other trace elements. J. Environ. Manag. 2014, 146, 94–99. [Google Scholar] [CrossRef]
- Laureysens, I.; Blust, R.; De Temmerman, L.; Lemmens, C.; Ceulemans, R. Clonal variation in heavy metal accumulation and biomass production in a poplar coppice culture: I. Seasonal variation in leaf, wood and bark concentrations. Environ. Pollut. 2004, 131, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Perttu, K.; Kowalik, P. Salix vegetation filters for purification of waters and soils. Biomass Bioenergy 1997, 12, 9–19. [Google Scholar] [CrossRef]
- Dietz, A.C.; Schnoor, J.L. Advances in phytoremediation. Environ. Health Perspect. 2001, 109, 163–168. [Google Scholar] [CrossRef]
- Vervaeke, P.; Luyssaert, S.; Mertens, J.; Meers, E.; Tack, F.; Lust, N. Phytoremediation prospects of willow stands on contaminated sediment: A field trial. Environ. Pollut. 2003, 126, 275–282. [Google Scholar] [CrossRef]
- Holm, B.; Heinsoo, K. Municipal wastewater application to short rotation coppice of willows—Treatment efficiency and clone response in an Estonian case study. Biomass Bioenergy 2013, 57, 126–135. [Google Scholar] [CrossRef]
- Isebrands, J.G.; Aronsson, P.; Carlson, M.; Ceulemans, R.; Coleman, M.; Dickinson, N.; Dimitriou, J.; Doty, S.; Gardiner, E.; Heinsoo, K.; et al. Environmental applications of poplars and willows. In Poplars and Willows: Trees for Society and the Environment; Isebrands, J.G., Richardson, J., Eds.; Food and Agriculture Organization (FAO) of the United Nations: Rome, Italy; CAB International, Inc.: Boston, MA, USA, 2014; Chapter 6; pp. 258–336. ISBN 978-1-78064-108-9. [Google Scholar]
- Monk, C. Ecological importance of root/shoot Ratios. Bull. Torrey Bot. Club 1966, 93, 402–406. [Google Scholar] [CrossRef]
- Lloret, F.; Casanovas, C.; Penuelas, J. Seedling survival of Mediterranean shrubland species in relation to root: Shoot ratio, seed size and water and nitrogen use. Funct. Ecol. 1999, 13, 210–216. [Google Scholar] [CrossRef]
- Van der Werf, A.; Visser, A.J.; Schieving, F.; Lambers, H. Evidence for optimal partitioning of biomass and nitrogen at a range of nitrogen availabilities for a fast- and slow-growing species. Funct. Ecol. 1993, 7, 63–74. [Google Scholar] [CrossRef]
- Gedroc, J.J.; Mcconnaughay, K.D.; Coleman, J.S. Plasticity in root/shoot partitioning: Optimal, ontogenetic, or both? Funct. Ecol. 1996, 10, 44–50. [Google Scholar] [CrossRef]
- Friend, A.L.; Coleman, M.D.; Isebrands, J.G. Carbon allocation to root and shoot systems of woody plants. In Biology of Adventitious Root Formation; Davis, T.D., Haissig, B.E., Eds.; Plenum Press: New York, NY, USA, 1994; pp. 245–273. [Google Scholar]
- Adler, A.; Karacic, A.; Weih, M. Biomass allocation and nutrient use in fast growing woody and herbaceous perennials used for phytoremediation. Plant Soil 2008, 305, 189–206. [Google Scholar] [CrossRef]
- Weih, M.; Nordh, N. Characterising willows for biomass and phytoremediation: Growth, nitrogen and water use of 14 willow clones under different irrigation and fertilisation regimes. Biomass Bioenergy 2002, 23, 397–413. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Yin, T.M.; Difazio, S.P.; Tschaplinski, T.J.; Gunter, L.E.; Davis, M.F.; Tuskan, G.A. Phenotypic variation in growth and biomass distribution for two advanced-generation pedigrees of hybrid poplar. Can. J. For. Res. 2005, 35, 1779–1789. [Google Scholar] [CrossRef]
- Rytter, R. Biomass production and allocation, including fine-root turnover, and annual N uptake in lysimeter-grown basket willows. For. Ecol. Manag. 2001, 140, 177–192. [Google Scholar] [CrossRef]
- Barigah, T.; Saugier, B.; Mousseau, M.; Guittet, J.; Ceulemans, R. Photosynthesis, leaf area and productivity of 5 poplar clones during their establishment year. Ann. Sci. For. 1994, 51, 613–625. [Google Scholar] [CrossRef] [Green Version]
- Pallardy, S.; Gibbins, D.; Rhoads, J. Biomass production by two-year-old poplar clones on floodplain sites in the Lower Midwest, USA. Agroforest. Syst. 2003, 59, 21–26. [Google Scholar] [CrossRef]
- U.S. Climate Data. Available online: http://www.usclimatedata.com/ (accessed on 23 April 2017).
- Green, R. Sampling Design and Statistical Methods for Environmental Biologists; Wiley: New York, NY, USA, 1979; ISBN 9780471039013. [Google Scholar]
- Pezeshki, S.R.; Anderson, P.H.; Shields, F.D. Effects of soil moisture regimes on growth and survival of black willow (Salix nigra) posts (cuttings). Wetlands 1998, 18, 460–470. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Hall, R.B.; Zalesny, J.A.; McMahon, B.G.; Berguson, W.E.; Stanosz, G.R. Biomass and genotype × environment interactions of Populus energy crops in the Midwestern United States. BioEnergy Res. 2009, 2, 106–122. [Google Scholar] [CrossRef]
- Marmiroli, M.; Pietrini, F.; Maestri, E.; Zacchini, M.; Marmiroli, N.; Massacci, A. Growth, physiological and molecular traits in Salicacae trees investigated for phytoremediation of heavy metals and organics. Tree Physiol. 2011, 31, 1319–1334. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Treatment | ||||||
|---|---|---|---|---|---|---|
| Bellevue | Caledonia | Menomonee Falls | Slinger | Whitelaw | Control | |
| pH | 7.64 ± 0.02 | 7.20 ± 0.01 | 7.52 ± 0.02 | 7.37 ± 0.01 | 7.74 ± 0.02 | 4.49 ± 0.01 |
| Texture | Clay Loam | Clay | Clay Loam | Clay Loam | Sandy Loam | - |
| Percent | ||||||
| Clay | 38 | 48 | 28 | 28 | 7 | - |
| Sand | 37 | 21 | 33 | 25 | 55 | - |
| Silt | 35 | 31 | 39 | 47 | 38 | - |
| C | 0.024 ± 0.001 | 0.029 ± 0.004 | 0.064 ± 0.000 | 0.024 ± 0.001 | 0.041 ± 0.003 | 0.310 ± 0.005 |
| N | 0.000 ± 0.000 | 0.001 ± 0.000 | 0.001 ± 0.000 | 0.001 ± 0.000 | 0.001 ± 0.000 | 0.007 ± 0.000 |
| g kg−1 | ||||||
| Al | 41.82 ± 0.56 | 52.46 ± 4.21 | 31.68 ± 0.30 | 45.71 ± 0.32 | 35.43 ± 4.62 | 22.75 ± 0.67 |
| Ca | 43.19 ± 1.78 | 30.51 ± 8.27 | 97.14 ± 1.12 | 23.22 ± 0.24 | 62.74 ± 8.14 | 11.24 ± 0.20 |
| Fe | 22.19 ± 0.30 | 29.56 ± 2.21 | 20.34 ± 0.18 | 26.29 ± 0.27 | 21.14 ± 2.11 | 16.61 ± 0.38 |
| K | 20.81 ± 0.24 | 19.57 ± 1.27 | 11.58 ± 0.36 | 13.88 ± 0.20 | 14.78 ± 1.69 | 4.43 ± 0.40 |
| Na | 5.40 ± 0.05 | 4.45 ± 0.12 | 4.14 ± 0.03 | 6.21 ± 0.05 | 4.95 ± 0.16 | 2.71 ± 0.15 |
| Ti | 2.56 ± 0.02 | 3.39 ± 0.24 | 2.03 ± 0.01 | 3.23 ± 0.01 | 2.35 ± 0.24 | 2.48 ± 0.06 |
| Si | 255.47 ± 3.59 | 234.58 ± 6.19 | 163.75 ± 1.42 | 263.10 ± 1.00 | 209.66 ± 3.23 | 76.98 ± 2.51 |
| mg kg−1 | ||||||
| Mg | 17.68 ± 0.63 | 21.14 ± 4.06 | 47.25 ± 0.31 | 15.08 ± 0.17 | 37.21 ± 3.85 | 25.53 ± 0.65 |
| Mn | 396.80 ± 8.50 | 648.96 ± 66.74 | 512.31 ± 4.55 | 790.67 ± 5.27 | 392.88 ± 66.65 | 257.87 ± 5.76 |
| P | 435.47 ± 8.75 | 426.05 ± 4.79 | 357.47 ± 5.89 | 445.51 ± 3.14 | 381.91 ± 9.96 | 380.88 ± 12.91 |
| Sr | 114.21 ± 0.58 | 101.06 ± 1.55 | 100.34 ± 0.27 | 103.01 ± 0.43 | 101.50 ± 1.36 | 56.84 ± 1.60 |
| Zr | 16.58 ± 0.15 | 16.55 ± 0.51 | 11.92 ± 0.38 | 23.77 ± 0.26 | 13.30 ± 0.94 | 2.00 ± 0.13 |
| Genomic Group | Clone |
|---|---|
| Populusa | |
| P. deltoides × P. maximowiczii ‘DM’ | 313.55, DM111, NC14106 |
| P. deltoides × P. nigra ‘DN’ | 9732-32, 9732-36, 21700, 99038022, 99038026, BR 3960, DN5, DN34, DN177 |
| P. nigra × P. maximowiczii ‘NM’ | NM2, NM6 |
| (P. trichocarpa × P. deltoides) × P. deltoides ‘TDD’ | NC13820 |
| Salix b | |
| S. caprea hybrid ‘C’ | S365 |
| S. miyabeana ‘M’ | SX67 |
| S. purpurea ‘P’ | Allegany |
| S. purpurea × S. miyabeana ‘PM’ | Millbrook |
| S. sachalinensis × S. miyabeana ‘SM’ | Canastota |
| S. viminalis × S. miyabeana ‘VM’ | Fabius, Owasco, Tully Champion |
| S. viminalis × (S. sachalinensis × S. miyabeana) ‘VSM’ | Preble |
| Source of Variation | Root–Shoot Ratio | Tree Health | Growth Index |
|---|---|---|---|
| Populus | |||
| Soil treatment | 0.1427 | 0.2877 | <0.0001 |
| Clone | 0.0726 | <0.0001 | 0.0003 |
| Soil treatment × clone | 0.9778 | 0.0196 | 0.1750 |
| Salix | |||
| Soil treatment | 0.0697 | 0.1842 | <0.0001 |
| Clone | 0.0049 | <0.0001 | 0.0029 |
| Soil treatment × clone | 0.3145 | 0.9550 | 0.0755 |
| Performance Group | Genomic Group | Clone | Soil Treatment | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bellevue | Caledonia | Menomonee Falls | Slinger | Whitelaw | Control | |||||||||
| Generalist | DM | 313.55 | 2.7 ± 0.3 | a | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | 2.3 ± 0.3 | ab | 2.7 ± 0.3 | a |
| Generalist | DM | DM111 | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | ne | 2.3 ± 0.3 | ab | 2.0 ± 0.0 | bc | 2.3 ± 0.7 | ab | |
| Generalist | DM | NC14106 | 2.7 ± 0.3 | a | 3.0 ± 0.0 | a | ne | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | |
| Specialist | DN | 9732-32 | 2.5 ± 0.4 | ab | 2.0 ± 0.0 | bc | 2.0 ± 0.0 | bc | 2.7 ± 0.3 | a | 2.3 ± 0.3 | ab | 2.0 ± 0.0 | bc |
| Specialist | DN | 9732-36 | 3.0 ± 0.0 | a | 2.0 ± 0.0 | bc | 2.0 ± 0.0 | bc | 2.5 ± 0.4 | ab | 3.0 ± 0.0 | a | 1.7 ± 0.3 | c |
| Specialist | DN | 21700 | 2.0 ± 0.0 | bc | 2.0 ± 0.0 | bc | 3.0 ± 0.0 | a | 2.3 ± 0.3 | ab | 3.0 ± 0.0 | a | 2.0 ± 0.6 | bc |
| Specialist | DN | 99038022 | 2.0 ± 0.0 | bc | 2.7 ± 0.3 | a | 2.0 ± 0.0 | bc | 2.7 ± 0.3 | a | 2.0 ± 0.0 | bc | 2.0 ± 0.0 | bc |
| Generalist | DN | 99038026 | 2.0 ± 0.0 | bc | 2.5 ± 0.4 | ab | 2.0 ± 0.6 | bc | 1.7 ± 0.3 | c | 2.0 ± 0.0 | bc | 2.0 ± 0.0 | bc |
| Specialist | DN | BR 3960 | 2.7 ± 0.3 | a | 2.3 ± 0.3 | ab | 2.0 ± 0.0 | bc | 2.3 ± 0.3 | ab | 2.0 ± 0.0 | bc | 2.0 ± 0.6 | bc |
| Generalist | DN | DN5 | 3.0 ± 0.0 | a | 2.7 ± 0.3 | a | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | 2.5 ± 0.4 | ab | 3.0 ± 0.0 | a |
| Specialist | DN | DN34 | 3.0 ± 0.0 | a | 2.0 ± 0.0 | bc | 3.0 ± 0.0 | a | 2.7 ± 0.3 | a | 2.0 ± 0.0 | bc | 2.0 ± 0.0 | bc |
| Generalist | DN | DN177 | 2.7 ± 0.3 | a | 2.5 ± 0.4 | ab | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | 2.3 ± 0.3 | ab | 2.7 ± 0.3 | a |
| Generalist | NM | NM2 | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | 2.5 ± 0.4 | ab | 3.0 ± 0.0 | a |
| Generalist | NM | NM6 | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | 2.7 ± 0.3 | a | 3.0 ± 0.0 | a |
| Generalist | TDD | NC13820 | 2.5 ± 0.4 | ab | 3.0 ± 0.0 | a | 3.0 ± 0.0 | a | 2.7 ± 0.3 | a | 3.0 ± 0.0 | a | 2.7 ± 0.3 | a |
| Across | 2.7 ± 0.1 | 2.6 ± 0.1 | 2.6 ± 0.1 | 2.7 ± 0.1 | 2.4 ± 0.1 | 2.4 ± 0.1 | ||||||||
| Diameter | Health | Height | RSR | Total Biomass | Growth Index | |
|---|---|---|---|---|---|---|
| Diameter | −0.17 0.0120 | 0.91 <0.0001 | 0.45 <0.0001 | 0.89 <0.0001 | 0.94 <0.0001 | |
| Health | −0.18 0.0245 | −0.12 0.0687 | −0.08 0.2215 | −0.15 0.0213 | −0.18 0.0066 | |
| Height | 0.91 <0.0001 | −0.16 0.0439 | 0.36 <0.0001 | 0.85 <0.0001 | 0.95 <0.0001 | |
| RSR | 0.20 0.0143 | −0.09 0.2826 | 0.12 0.1381 | 0.56 <0.0001 | 0.50 <0.0001 | |
| Total biomass | 0.80 <0.0001 | −0.17 0.0390 | 0.75 <0.0001 | 0.22 0.0068 | 0.88 <0.0001 | |
| Growth index | 0.96 <0.0001 | −0.13 0.1155 | 0.94 <0.0001 | 0.14 0.0865 | 0.91 <0.0001 |
| Genus | Model | Diameter | Height | Total Biomass | Growth Index |
|---|---|---|---|---|---|
| Populus | T | <0.0001 | <0.0001 | 0.0451 | <0.0001 |
| RSR | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| T × RSR | <0.0001 | 0.0009 | <0.0001 | 0.0003 | |
| r2 | 0.76 | 0.64 | 0.81 | 0.66 | |
| C | 0.6784 | 0.0233 | 0.5592 | 0.0473 | |
| RSR | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| C × RSR | 0.0597 | 0.0770 | 0.0553 | 0.0630 | |
| r2 | 0.43 | 0.40 | 0.48 | 0.42 | |
| Salix | T | <0.0001 | <0.0001 | 0.0008 | <0.0001 |
| RSR | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| T × RSR | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| r2 | 0.69 | 0.57 | 0.76 | 0.72 | |
| C | 0.1344 | 0.0613 | 0.5170 | 0.2447 | |
| RSR | 0.0016 | 0.0103 | 0.0004 | 0.0159 | |
| C × RSR | 0.0341 | 0.0202 | 0.0572 | 0.0505 | |
| r2 | 0.19 | 0.21 | 0.20 | 0.16 |
| Genus | Slope (Treatment) | Diameter | Height | Total Biomass | Growth Index | ||||
|---|---|---|---|---|---|---|---|---|---|
| Est | p-Value | Est | p-Value | Est | p-Value | Est | p-Value | ||
| Populus | Menomonee Falls | 45.4 | 0.0006 | 709.9 | 0.0035 | 106.9 | 0.0696 | 19.9 | 0.0057 |
| Control | 57.1 | <0.0001 | 628.7 | <0.0001 | 419.8 | <0.0001 | 19.2 | <0.0001 | |
| Bellevue | 44.7 | <0.0001 | 631.3 | 0.0001 | 161.2 | <0.0001 | 23.0 | <0.0001 | |
| Slinger | 32.2 | 0.0056 | 497.2 | 0.0202 | 128.1 | 0.0143 | 16.7 | 0.0087 | |
| Caledonia | 73.6 | <0.0001 | 1011.7 | 0.0003 | 312.3 | <0.0001 | 33.9 | <0.0001 | |
| Whitelaw | 7.8 | 0.1527 | 105.9 | 0.2957 | 61.0 | 0.0141 | 4.0 | 0.1809 | |
| Salix | Menomonee Falls | 39.9 | <0.0001 | 992.3 | <0.0001 | 112.9 | 0.0029 | 19.1 | <0.0001 |
| Control | 30.3 | <0.0001 | 360.9 | 0.0573 | 353.6 | <0.0001 | 19.2 | <0.0001 | |
| Bellevue | −1.1 | 0.6915 | −115.4 | 0.1426 | 14.3 | 0.2771 | −1.7 | 0.2933 | |
| Slinger | 4.6 | 0.4018 | 86.7 | 0.5639 | 43.8 | 0.0828 | 1.5 | 0.6220 | |
| Caledonia | 25.3 | 0.0014 | 698.1 | 0.0013 | 98.7 | 0.0062 | 12.9 | 0.0034 | |
| Whitelaw | 11.0 | 0.0584 | 112.3 | 0.4797 | 48.9 | 0.0675 | 3.1 | 0.3404 | |
| Genus | Slope (Clone) | Diameter | Height | ||
|---|---|---|---|---|---|
| Est | p-Value | Est | p-Value | ||
| Salix | Allegany | 17.4 | 0.0597 | 497.6 | 0.0199 |
| Canastota | 12.8 | 0.3139 | 267.5 | 0.3615 | |
| Fabius | 0.5 | 0.9155 | −41.4 | 0.6776 | |
| Millbrook | 12.4 | 0.0833 | 212.3 | 0.1955 | |
| Owasco | 17.3 | 0.0045 | 307.8 | 0.0273 | |
| Preble | 12.9 | 0.1058 | 126.3 | 0.4906 | |
| S365 | 10.4 | 0.4382 | −382.9 | 0.2166 | |
| SX67 | 29.4 | 0.0212 | 663.7 | 0.0237 | |
| Tully Champion | 23.2 | 0.0079 | 376.9 | 0.0590 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rogers, E.R.; Zalesny, R.S., Jr.; Hallett, R.A.; Headlee, W.L.; Wiese, A.H. Relationships among Root–Shoot Ratio, Early Growth, and Health of Hybrid Poplar and Willow Clones Grown in Different Landfill Soils. Forests 2019, 10, 49. https://doi.org/10.3390/f10010049
Rogers ER, Zalesny RS Jr., Hallett RA, Headlee WL, Wiese AH. Relationships among Root–Shoot Ratio, Early Growth, and Health of Hybrid Poplar and Willow Clones Grown in Different Landfill Soils. Forests. 2019; 10(1):49. https://doi.org/10.3390/f10010049
Chicago/Turabian StyleRogers, Elizabeth R., Ronald S. Zalesny, Jr., Richard A. Hallett, William L. Headlee, and Adam H. Wiese. 2019. "Relationships among Root–Shoot Ratio, Early Growth, and Health of Hybrid Poplar and Willow Clones Grown in Different Landfill Soils" Forests 10, no. 1: 49. https://doi.org/10.3390/f10010049
APA StyleRogers, E. R., Zalesny, R. S., Jr., Hallett, R. A., Headlee, W. L., & Wiese, A. H. (2019). Relationships among Root–Shoot Ratio, Early Growth, and Health of Hybrid Poplar and Willow Clones Grown in Different Landfill Soils. Forests, 10(1), 49. https://doi.org/10.3390/f10010049