Validation and Application of European Beech Phenological Metrics Derived from MODIS Data along an Altitudinal Gradient



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Stands and In Situ Phenological Observations
- The representation of beech in a 250 × 250 m pixel (corresponding to MO09GQ resolution) was derived from the classification of the tree species composition in Slovakia [20] from Landsat satellite images in 30 m spatial resolution using the ArcGIS Aggregate function. The pixels with presence of beech 60% and higher were included in the mask.
- In order to eliminate a possible tree classification error from Landsat, the Forestry Information System database was used. Here, the actual data of tree species composition are available for each forest compartment (generally, compartment area varies from 1 to 15 ha). The presence of beech from the database was assigned to the pixels selected in the first step. Pixels with beech 60% and higher remained in the beech mask.
- Due to the possible contamination of DN values with non-forest land cover classes (meadows, fields, water areas, etc.), pixels at the edges of the forest were removed from the derived beech mask.
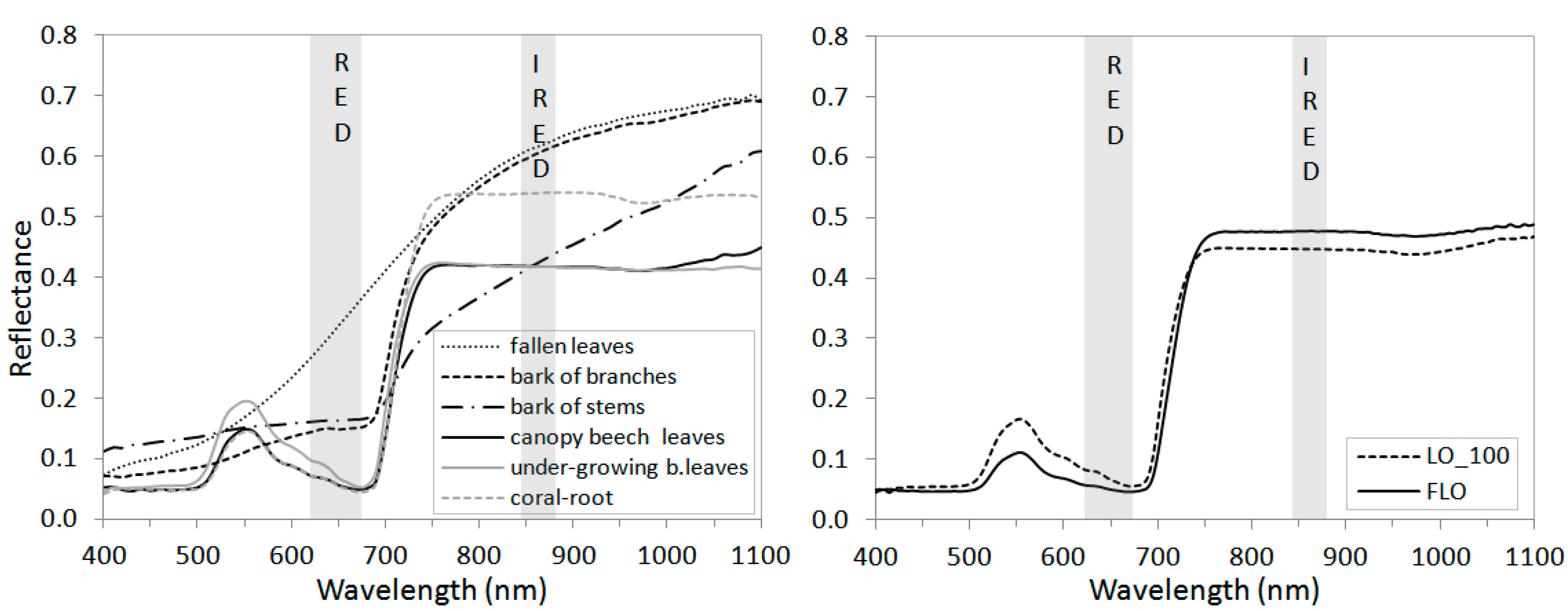
2.2. Validation Supporting Laboratory Spectral Analyses
2.3. Deriving MODIS NDVI Phenological Metrics
- S1—the acceleration of leafing in forest stand.
- S2—the leafing in forest stand reaches the half-maximum.
- S3—the deceleration of leafing in forest stand.
- A1—the acceleration of leaf coloring in forest stand.
- A2—the leaf coloring in forest stand reaches the half-maximum.
- A3—the deceleration of leaf coloring in forest stand.
2.4. Altitudinal Study
3. Results
3.1. What Is Hidden behind the NDVI Value of Beech-Dominated Stands?
3.2. Assigning the Phenological Metrics to the In Situ Observed Phenological Phases
3.2.1. Spring Phenological Phases
- In situ observed phenological phases such as bud swelling and bud bursting were not possible to observe in the satellite data because the increasing NDVIMOD at the time of their occurrence was caused by other forest components, and not by the canopy beech leaves. Therefore, phenological metric S1 was not paired with any in situ observed phenological phases.
- Phenological metric S2 was assigned to the beginning of leaf onset (LO_10). The paired sample t-test revealed non-significant differences between S2 and LO_10 (t = 0.03 < T0.01(90) = 1.96). The correlation coefficient indicated very strong correlation between LO_10 and S2 (Figure 6). The average NDVIMOD of the S2 was 0.67.
- Phenological metric S3 was assigned to the phenological phase LO_50, and although the r = 0.84 (Figure 6) suggested a very strong correlation between S3 and LO_50, the t-test revealed significant differences with a RMSE of 6.24. The phenological metric S3 was overestimated. The phenological data from the SHMI, which were the base data for the validation, did not contain information on the phenological phase LO_100, which, according to our detailed phenological monitoring, occurred on average four days after LO_50. No significant differences were revealed between S3 and LO_100 (t = 0.10 < T0.01(9) = 3.25) in our U1–U5 stands, although the correlation coefficient was smaller than the one between S3 and LO_50 (Figure 6). A control test by adding four days (average difference between LO_50 and LO_100) to the days of LO_50 from the SHMI observations was performed. The differences between the estimated LO_100 and S3 were non-significant (t = 0.54 < T0.01(90) = 1.96) with the RMSE of 4.57. The average NDVIMOD of S3 was 0.81.
3.2.2. Autumn Phenological Phases
- Phenological metric A1 occurred between the two autumn phenological phases: beginning of leaf coloring (LC_10) and general leaf coloring (LC_50), on average eight days after LC_10. Thus, we paired A1 with LC_10 and LC_50 from all twelve study stands and single years, but the differences in both cases were significant. Then, using data from detailed phenological monitoring on the U1–U5 stands, the pairs of A1 with LC_20, LC_30, and LC_40 were tested, but the differences in all of the tested pairs were significant.
- The paired test of the onset days of A2 and LC_50 revealed an average time lag of 13 days for the A2 phenological metric. Based on that, the pairs of A2 with LC_60, LC_70, LC_80, LC_90, and LC_100 recorded in the U1–U5 stands were tested. Only the paired t-test with LC_80 revealed non-significant differences (t = 1.51 < T0.01(9) = 3.25) between the in situ LC_80 and satellite data-based A2.
- A3 occurred after the end of leaf coloring (LC_100) when only the understory vegetation was green, but the beech trees had no green leaves. Therefore, this phenological metric was not paired with any autumn in situ observed phenological phase.
- The correlation between the onset days of the autumn phenological phases and phenological metrics was weak, with the correlation coefficient not exceeding 0.30 (Figure 7).
3.3. Phenological Metrics along the Altitudinal Gradient
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ciais, P.; Sabine, C.; Bala, G.; Bopp, L.; Brovkin, V.; Canadell, J.; Chhabra, A.; DeFries, R.; Galloway, J.; Heimann, M.; et al. Carbon and Other Biogeochemical Cycles. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 465–570. [Google Scholar] [CrossRef]
- Badeck, F.W.; Bondeau, A.; Böttcher, K.; Doktor, D.; Lucht, W.; Schaber, J.; Sitch, S. Responses of spring phenology to climate change. New Phytol. 2004, 162, 295–309. [Google Scholar] [CrossRef] [Green Version]
- Polgar, C.; Gallinat, A.; Primack, R.B. Drivers of leaf-out phenology and their implications for species invasions: Insights from Thoreau’s Concord. New Phytol. 2014, 202, 106–115. [Google Scholar] [CrossRef]
- Guyon, D.; Guillot, M.; Vitasse, Y.; Cardot, H.; Hagolle, O.; Delzon, S.; Wigneron, J.P. Monitoring elevation variations in leaf phenology of deciduous broadleaf forests from SPOT/VEGETATION time-series. Remote Sens. Environ. 2011, 115, 615–627. [Google Scholar] [CrossRef]
- Mátyás, C.; Berki, I.; Czúcz, B.; Gálos, B.; Móricz, N.; Rastovits, E. Future of Beech in Southeast Europe from the Perspective of Evolutionary Ecology. Acta Silv. Lignaria Hung. 2010, 6, 91–110. [Google Scholar]
- Becker, M. Taxonomie et characteres botaniques. In Le Hetre; Teissier du Cros, E., Ed.; INRA: Paris, France, 1981; pp. 35–49. ISBN 978-2-7592-0512-7. [Google Scholar]
- Myneni, R.B.; Hoffman, S.; Knyazikhin, Y.; Privette, J.L.; Glassy, J.; Tian, Y.; Wang, Y.; Song, X.; Zhang, Y.; Smith, G.R.; et al. Global products of vegetation leaf area and fraction absorbed PAR from year one of MODIS data. Remote Sens. Environ. 2002, 83, 214–231. [Google Scholar] [CrossRef] [Green Version]
- Fisher, J.I.; Mustard, J.F.; Vadeboncoeur, M.A. Green leaf phenology at Landsat resolution: Scaling from the field to the satelite. Remote Sens. Environ. 2006, 100, 265–279. [Google Scholar] [CrossRef]
- Fisher, J.I.; Mustard, J.F. Cross-scalar satellite phenology from ground, Landsat and MODIS data. Remote Sens. Environ. 2007, 109, 261–273. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Atzberger, C.; Høgda, K.A.; Johansen, B.; Skidmore, A.K. Improved monitoring of vegetation dynamics at very high latitudes: A new method using MODIS NDVI. Remote Sens. Environ. 2006, 100, 321–334. [Google Scholar] [CrossRef]
- Jönsson, A.M.; Hellström, M.; Bärring, L.; Jönsson, P. Annual changes in MODIS vegetation indices of Swedish coniferous forests in relation to snow dynamics and tree phenology. Remote Sens. Environ. 2010, 114, 2719–2730. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Heute, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- White, K.; Pontius, J.; Schaberg, P. Remote sensing of spring phenology in northeastern forests: A comparison of methods, field metrics and sources of uncertainty. Remote Sens. Environ. 2014, 148, 97–107. [Google Scholar] [CrossRef]
- Liang, L.; Schwartz, M.D.; Fei, S. Validating satellite phenology through intensive ground observation and landscape scaling in a mixed seasonal forest. Remote Sens. Environ. 2011, 115, 143–157. [Google Scholar] [CrossRef]
- Chen, X.; Luo, X.; Xu, L. Comparison of spatial patterns of satellite-derived and ground-based phenology for the deciduous broadleaf forest of China. Remote Sens. Lett. 2013, 4, 532–541. [Google Scholar] [CrossRef]
- Reed, B.C.; Brown, J.F.; VanderZee, R.; Merchant, J.W.; Ohlen, D.O. Measuring phenological variability from satellite imagery. Int. Assoc. Veg. Sci. 1994, 5, 703–714. [Google Scholar] [CrossRef]
- Testa, S.; Soudani, K.; Boschetti, L.; Borgogno Mondino, E. MODIS-derived EVI, NDVI and WDRVI time series to estimate phenological metrics in French deciduous forests. Int. J. Appl. Earth Obs. Geoinf. 2018, 64, 132–144. [Google Scholar] [CrossRef]
- Škvarerninová, J.; Snopková, Z. The Development of Phenological Stages of European Beach (Fagus sylvatica L.) in Slovakia During the Period of 1996–2010. In Proceedings of the Bioclimate: Source and Limit of Social Development, Topoľčianky, Slovakia, 6–9 September 2011; pp. 87–90. [Google Scholar]
- Škvareninová, J. Vplyv Zmeny Klimatických Podmienok na Fenologickú Odozvu Ekosystémov; Vydavateľstvo Technickej Univerzity vo Zvolene: Zvolen, Slovakia, 2013; 132p, ISBN 978-80-228-2598-6. [Google Scholar]
- Bucha, T. Classification of tree species composition in Slovakia from satellite images as a part of monitoring forest ecosystems biodiversity. Acta Inst. For. Zvolen 1999, 9, 65–84. [Google Scholar]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Berlin, Germany; Braunschweig, Germany, 2001; 158p. [Google Scholar]
- Bruns, E.; Chmielewski, F.M.; van Vliet, A.J.H. The Global Phenological Monitoring Concept. In Phenology: An Integrative Environmental Science; Schwartz, M., Ed.; Springer: Dordrecht, The Netherlands, 2003; pp. 93–104. ISBN 978-94-007-0632-3. [Google Scholar]
- Chmielewski, F.M.; Rötzer, T. Phenological trends in Europe in relation to climatic changes. Agrometeorol. Schr. 2000, 7, 1–15. [Google Scholar]
- Škvareninová, J.; Tuhárska, M.; Škvarenina, J.; Babálová, D.; Slobodníková, L.; Slobodník, B.; Středová, H.; Minďaš, J. Effects of light pollution on tree phenology in the urban environment. Morav. Geogr. Rep. 2017, 25, 282–290. [Google Scholar] [CrossRef] [Green Version]
- Babálová, D.; Škvareninová, J.; Fazekaš, J.; Vyskot, I. The dynamics of the phenological development of four woody species in south-west and central Slovakia. Sustainability 2018, 10, 1497. [Google Scholar] [CrossRef]
- Lukasova, V.; Lang, M.; Skvarenina, J. Seasonal changes in NDVI in relation to phenological phases, LAI and PAI of beech forests. Balt. For. 2014, 20, 248–262. [Google Scholar]
- Land Processes Distributed Active Archive Center. Available online: https://lpdaac.usgs.gov/dataset_discovery/modis/modis_products_table/mod09gq_v006 (accessed on 13 March 2018).
- Ju, J.; Roy, D.P.; Shuai, Y.; Schaaf, C. Development of an approach for generation of temporally complete daily nadir MODIS reflectance time series. Remote Sens. Environ. 2011, 114, 1–20. [Google Scholar] [CrossRef]
- Franch, B.; Vermote, E.F.; Sobrino, J.A.; Fédèle, E. Analysis of directional effect on atmospheric correction. Remote Sens. Environ. 2013, 128, 276–288. [Google Scholar] [CrossRef]
- Townshend, J.R.G.; Huang, S.N.; Kalluri, V.; Defries, R.S.; Liang, S. Beware of the per-pixel characterization of land cover. Int. J. Remote Sens. 2000, 21, 839–843. [Google Scholar] [CrossRef]
- Bucha, T.; Koreň, M. Phenology of the beech forests in the Western Carpathians from MODIS for 2000–2015. iForest Biogeosci. For. 2017, 10, 537–546. [Google Scholar] [CrossRef]
- Evans, J.D. Straightforward Statistics for the Behavioral Sciences; Brooks/Cole Publishing: Pacific Grove, CA, USA, 1996; 600p. [Google Scholar]
- Kramer, K. Phenotypic plasticity of the phenology of seven European tree species in relation to climatic warming. Plant Cell Environ. 1995, 18, 93–104. [Google Scholar] [CrossRef]
- Matsumoto, K.; Ohta, T.; Irasawa, M.; Nakamura, T. Climate change and extension of the Ginkgo biloba L. growing season in Japan. Glob. Chang. Biol. 2003, 9, 1634–1642. [Google Scholar] [CrossRef]
- Estrella, N.; Menzel, A. Response of leaf colouring in four deciduous tree species to climate and weather in Germany. Clim. Res. 2006, 32, 253–267. [Google Scholar] [CrossRef]
- Wang, T.; Ottle, C.; Peng, S.; Janssens, I.A.; Lin, X.; Poulter, B.; Yue, C.; Ciais, P. The influence of local spring temperature variation on temperature sensitivity of spring phenology. Global Chang. Biol. 2014, 20, 1473–1480. [Google Scholar] [CrossRef]
- Koike, T. Autumn coloring, photosynthetic performance and leaf development of deciduous broad-leaved trees in relation to forest succession. Tree Physiol. 1990, 7, 21–32. [Google Scholar] [CrossRef]
- Lee, D.W.; O’Keefe, J.; Holbrook, N.M.; Feild, T.S. Pigment dynamics and autumn leaf senescence in a New England deciduous forest, eastern USA. Ecol. Res. 2003, 18, 677–694. [Google Scholar] [CrossRef]
- Menzel, A. Phenology: Its importance to the global change community. Clim. Chang. 2002, 54, 379–385. [Google Scholar] [CrossRef]
- Broich, M.; Huete, A.; Tulbure, M.G.; Ma, X.; Xin, Q.; Paget, M.; Restrepo-Coupe, N.; Davies, K.; Devadas, R.; Held, A. Land surface phenological response to decadal climate variability across Australia using satellite remote sensing. BioGeoSci. Discuss. 2014, 11, 7685–7719. [Google Scholar] [CrossRef]
- Dittmar, C.; Elling, W. Phenological phases of common beech (Fagus sylvatica L.) and their dependence on region and altitude in Southern Germany. Eur. J. Forest Res. 2006, 125, 181–188. [Google Scholar] [CrossRef]
- Čufar, K.; Luis, M.D.; Saz, M.A.; Črepinšek, Z.; Kajfež-Bogataj, L. Temporal shifts in leaf phenology of beech (Fagus sylvatica) depend on elevation. Trees 2012, 26, 1091–1100. [Google Scholar] [CrossRef]
- Vitasse, Y.; Delzon, S.; Dufrene, E.; Pontiller, J.Y.; Louvet, J.M.; Kremer, A.; Michalet, R. Leaf phenology sensitivity to temperature in European trees: Do within-species populations exhibit similar responses? Agric. Forest Meteorol. 2009, 149, 735–744. [Google Scholar] [CrossRef]
- Dunn, A.H.; Beurs, K.M. Land surface phenology of North American mountain environments using moderate resolution imaging spectroradiometer data. Remote Sens. Environ. 2011, 115, 1220–1233. [Google Scholar] [CrossRef]
- Ahrens, C.D.; Henson, R. Meteorology today: An Introduction to Weather, Climate, and the Environment, 11th ed.; Cengage Learning: Boston, MA, USA, 2015; 656p, ISBN 978-1-305-26500-4. [Google Scholar]
- Climatic Conditions of Slovak Republic. Available online: http://www.shmu.sk/sk/?page=1064 (accessed on 12 March 2018).
- Pfanz, H.; Aschan, G. The Existence of Bark and Stem Photosynthesis in Woody Plants and Its Significance for the Overall Carbon Gain. An Eco-Physiological and Ecological Approach. In Progress in Botany; Springer: Berlin, Germany, 2001; Volume 62, pp. 477–510. [Google Scholar] [CrossRef]
- Richardson, A.D.; O’Keefe, J. Phenological differences between understory and overstory: A case study using the long-therm Harvard forest records. In Phenology of Ecosystem Processes; Noormets, A., Ed.; Springer: New York, NY, USA, 2009; pp. 88–117. [Google Scholar] [CrossRef]
- Ahl, D.E.; Gower, S.T.; Burrows, S.N.; Shabanov, N.V.; Myneni, R.B.; Knyazikhin, Y. Monitoring spring canopy phenology of a deciduous broadleaf forest using MODIS. Remote Sens. Environ. 2006, 104, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Nagai, S.; Nasahara, K.N.; Muraoka, H.; Akiyama, T.; Tsuchida, S. Field experiments to test the use of the normalized-difference vegetation index for phenology detection. Agric. Forest Meteorol. 2010, 150, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Tang, J.; Mustard, J.F. Beyond leaf color: Comparing camera-based phenological metrics with leaf biochemical, biophysical, and spectral properties throughout the growing season of a temperate deciduous forest. J. Geophys. Res. Biogeosci. 2014, 119, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Soudani, K.; Hmimina, G.; Delpierre, N.; Pontailler, J.Y.; Aubinet, M.; Bonal, D.; Caquet, B.; de Grandcourt, A.; Burban, B.; Flechard, C.; et al. Ground-based Network of NDVI measurements for tracking temporal dynamics of canopy structure and vegetation phenology in different biomes. Remote Sens. Environ. 2012, 123, 234–245. [Google Scholar] [CrossRef]
- Hmimina, G.; Dufrêne, E.; Pontailler, J.Y.; Delpierre, N.; Aubinet, M.; Caquet, B.; de Grandcourt, A.; Burban, B.; Flechard, C.; Granier, A.; et al. Evaluation of the potential of MODIS satellite data to predict vegetation phenology in different biomes: An investigation using ground-based NDVI measurements. Remote Sens. Environ. 2013, 132, 145–158. [Google Scholar] [CrossRef]
- García-Plazaola, J.I.; Becerril, J.M. Seasonal changes in photosynthetic pigments and antioxidants in beech (Fagus sylvatica) in a Mediterranean climate: Implications for tree decline diagnosis. Aust. J. Plant Physiol. 2001, 28, 225–232. [Google Scholar] [CrossRef]
- Soudani, K.; Maire, G.; Dufrêne, E.; François, C.H.; Delpierre, N.; Ulrich, E.; Cecchini, S. Evaluation of the onset of green-up in temperate deciduous broadleaf forests derived from Moderate Resolution Imaging Spectroradiometer (MODIS) data. Remote Sens. Environ. 2008, 112, 2643–2655. [Google Scholar] [CrossRef]
- Chalupa, V. Počátek, trvání a ukončení vegetační činnosti u lesních dřevin. Práce VÚLHM 1969, 37, 41–68. [Google Scholar]
- Toda, M.; Richardson, A.D. Estimation of plant area index and phenological transition dates from digital repeat photography and radiometric approaches in a hardwood forest in the Northeastern United States. Agric. Forest Meteorol. 2018, 249, 457–466. [Google Scholar] [CrossRef]
- Klosterman, S.; Richardson, A.D. Observing spring and fall phenology in a deciduous forest with aerial drone imagery. Sensors 2017, 17, 2852. [Google Scholar] [CrossRef]
- Nagai, S.; Inoue, T.; Ohtsuka, T.; Kobayashi, H.; Kurumado, K.; Muraoka, H.; Nasahara, K.N. Relationship between spatio-temporal characteristics of leaf-fall phenology and seasonal variations in near surface- and satellite-observed vegetation indices in cool-temperate deciduous broad-leaved forest in Japan. Int. J. Remote Sens. 2014, 35, 3520–3536. [Google Scholar] [CrossRef]
- Liu, L.; Liang, L.; Schwartz, M.D.; Donnelly, A.; Wang, Z.; Schaaf, C.B. Evaluating the potential of MODIS satellite data to track temporal dynamics of autumn phenology in a temperate mixed forest. Remote Sens. Environ. 2015, 160, 156–165. [Google Scholar] [CrossRef]
- Hwang, T.; Song, C.; Vose, J.M. Topography-mediated controls on local vegetation phenology estimated from MODIS vegetation index. Landsc. Ecol. 2011, 26, 541–556. [Google Scholar] [CrossRef]
- Dittmar, C.; Fricke, W.; Elling, W. Impact of late frost events on radial growth of common beech (Fagus sylvatica L.) in Southern Germany. Eur. J. Forest Res. 2006, 125, 249–259. [Google Scholar] [CrossRef]
- Kreyling, J.; Thiel, D.; Nagy, L.; Jentsch, A.; Huber, G.; Konnert, M.; Beierkuhnlein, C. Late frost sensitivity of juvenile Fagus sylvatica L. differs between southern Germany and Bulgaria and depends on preceding air temperature. Eur. J. Forest Res. 2012, 131, 717–725. [Google Scholar] [CrossRef]
- Kindermann, J.; Wurth, G.; Kohlmaier, G.H.; Badeck, F.W. Interannual variation of carbon exchange fluxes in terrestrial ecosystems. Global Biogeochem. Cycles 1996, 10, 737–755. [Google Scholar] [CrossRef]
- Gomory, D.; Paule, L. Trade-of between height growth and spring flushing in common beech (Fagus sylvatica L.). Ann. Forest Sci. 2011, 68, 975–984. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Number | Identifier | Altitude | Aspect | Presence of Beech (%) | Climatic Region | Climate Normal (1981–2010) | |
|---|---|---|---|---|---|---|---|
| T 7 (°C) | P 8 (mm) | ||||||
| 1 | ZS | 304 | NW | 60 | W7 1 | 10.1 | 675 |
| 2 | MY | 457 | W | 65 | M3 2 | 9 | 729 |
| 3 | U1 | 490 | W | 85 | M6 3 | 8.6 | 691 |
| 4 | U5 | 512 | N | 70 | M3 | 8.3 | 700 |
| 5 | U2 | 522 | W | 65 | M6 | 8.3 | 700 |
| 6 | ZV | 566 | SW | 65 | M6 | 8 | 745 |
| 7 | KC | 570 | SW | 90 | M3 | 8.1 | 644 |
| 8 | MU | 579 | N | 100 | M7 4 | 7.9 | 689 |
| 9 | U4 | 607 | E | 100 | M6 | 7.8 | 760 |
| 10 | U3 | 615 | W | 65 | M6 | 7.8 | 760 |
| 11 | CS | 1003 | N | 90 | C1 5 | 5.2 | 857 |
| 12 | PO | 1051 | SE | 80 | C2 6 | 5.7 | 863 |
| Phenological Phase | BBCH Codes | Definition |
|---|---|---|
| Beginning of Bud Bursting (BB_10) | BBCH07 | When bud scales opened and green top of leaf was sticking out |
| Beginning of Leaf Onset (LO_10) | BBCH11 | When 10% of leaves have final shape, but not final size and color |
| General Leaf Onset (LO_50) | BBCH13 | When 50% of leaves have final shape, but not final size and color |
| Beginning of Leaf Coloring (LC_10) | - | When 10% of leaves changed their color from green to yellow, red or brown |
| General Leaf Coloring (LC_50) | BBCH94 | When 50% of leaves changed their color from green to yellow, red or brown |
| Beginning of Leaf Fall (LF_10) | BBCH93 | When 10% of leaves fell down from trees to the ground |
| End of Leaf Fall (LF_100) | BBCH97 | When 100% of leaves fell down from trees to the ground |
| Time of Sampling–DOY | 97 | 110 | 127 | 159 |
|---|---|---|---|---|
| Date | 7.4 | 20.4 | 7.5 | 8.6 |
| Canopy beech leaves | - | - | 0.71 ± 0.023 (10) | 0.79 ± 0.015 (8) |
| Under-growing beech leaves | - | - | 0.66 ± 0.011 (15) | - |
| Fallen leaves | 0.34 ± 0.067 (8) | - | - | |
| Bark of branches | - | - | - | 0.63 ± 0.028 (5) |
| Bark of stems | - | - | - | 0.46 ± 0.042 (14) |
| Coral-root | - | 0.71 ± 0.012 (4) | - | - |
| Dogs mercury | - | 0.70 ± 0.003 (3) | - | - |
| NDVIMOD | 0.48 ± 0.062 (7) | 0.58 ± 0.017 (10) | 0.87 ± 0.010 (10) | 0.91 ± 0.008 (5) |
| Altitude | Altitudinal Gradient of Large-Scale Phenology | Difference on Altitudinal Gradient | ||||||
|---|---|---|---|---|---|---|---|---|
| S2 | S3 | A2 | GSL | S2 1 | S3 1 | A2 2 | GSL 3 | |
| 156 | 109 | 118 | 296 | 186 | 0 | - | −3 | −2 |
| 200 | 108 | 118 | 296 | 187 | 0 | 0 | −2 | −1 |
| 300 | 108 | 119 | 298 | 188 | - | 1 | −1 | - |
| 400 | 109 | 120 | 299 | 187 | 0 | 2 | 0 | 0 |
| 500 | 110 | 122 | 299 | 186 | 1 | 3 | - | −2 |
| 600 | 111 | 124 | 298 | 183 | 3 | 5 | 0 | −4 |
| 700 | 113 | 126 | 297 | 179 | 5 | 8 | −1 | −8 |
| 800 | 116 | 129 | 296 | 174 | 8 | 10 | −3 | −13 |
| 900 | 120 | 132 | 293 | 168 | 11 | 13 | −5 | −20 |
| 1000 | 124 | 135 | 291 | 161 | 15 | 17 | −8 | −27 |
| 1100 | 128 | 139 | 287 | 152 | 20 | 21 | −12 | −36 |
| 1200 | 133 | 143 | 283 | 142 | 25 | 25 | −16 | −45 |
| 1300 | 139 | 148 | 279 | 131 | 31 | 30 | −20 | −57 |
| 1331 | 141 | 150 | 277 | 127 | 33 | 31 | −22 | −60 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lukasová, V.; Bucha, T.; Škvareninová, J.; Škvarenina, J. Validation and Application of European Beech Phenological Metrics Derived from MODIS Data along an Altitudinal Gradient. Forests 2019, 10, 60. https://doi.org/10.3390/f10010060
Lukasová V, Bucha T, Škvareninová J, Škvarenina J. Validation and Application of European Beech Phenological Metrics Derived from MODIS Data along an Altitudinal Gradient. Forests. 2019; 10(1):60. https://doi.org/10.3390/f10010060
Chicago/Turabian StyleLukasová, Veronika, Tomáš Bucha, Jana Škvareninová, and Jaroslav Škvarenina. 2019. "Validation and Application of European Beech Phenological Metrics Derived from MODIS Data along an Altitudinal Gradient" Forests 10, no. 1: 60. https://doi.org/10.3390/f10010060
APA StyleLukasová, V., Bucha, T., Škvareninová, J., & Škvarenina, J. (2019). Validation and Application of European Beech Phenological Metrics Derived from MODIS Data along an Altitudinal Gradient. Forests, 10(1), 60. https://doi.org/10.3390/f10010060