Interspecific Differences in Canopy-Derived Water, Carbon, and Nitrogen in Upland Oak-Hickory Forest


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Stemflow, Throughfall, and Precipitation Measurements
2.4. Chemical Analysis
2.5. Statistical Analysis
3. Results
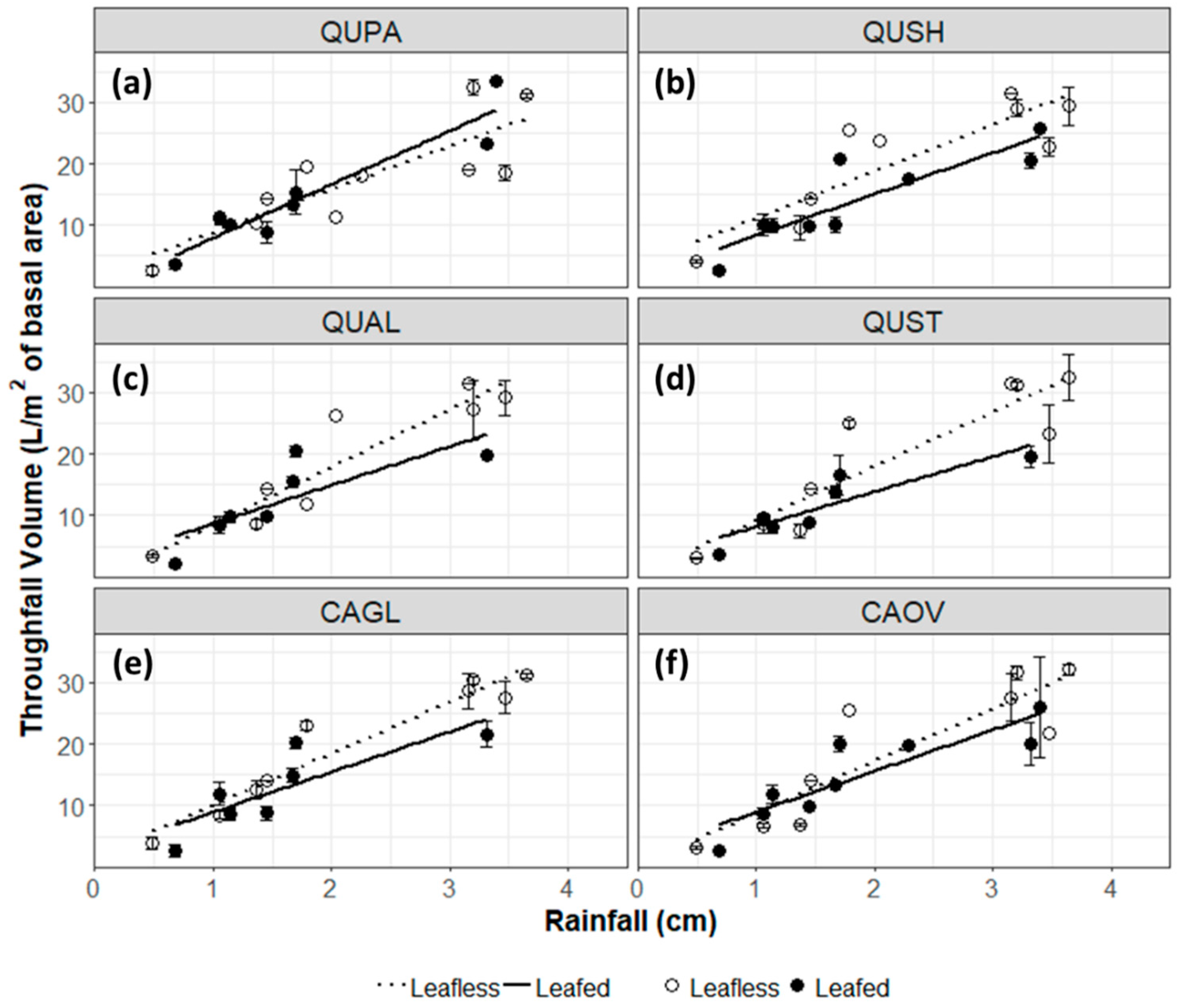
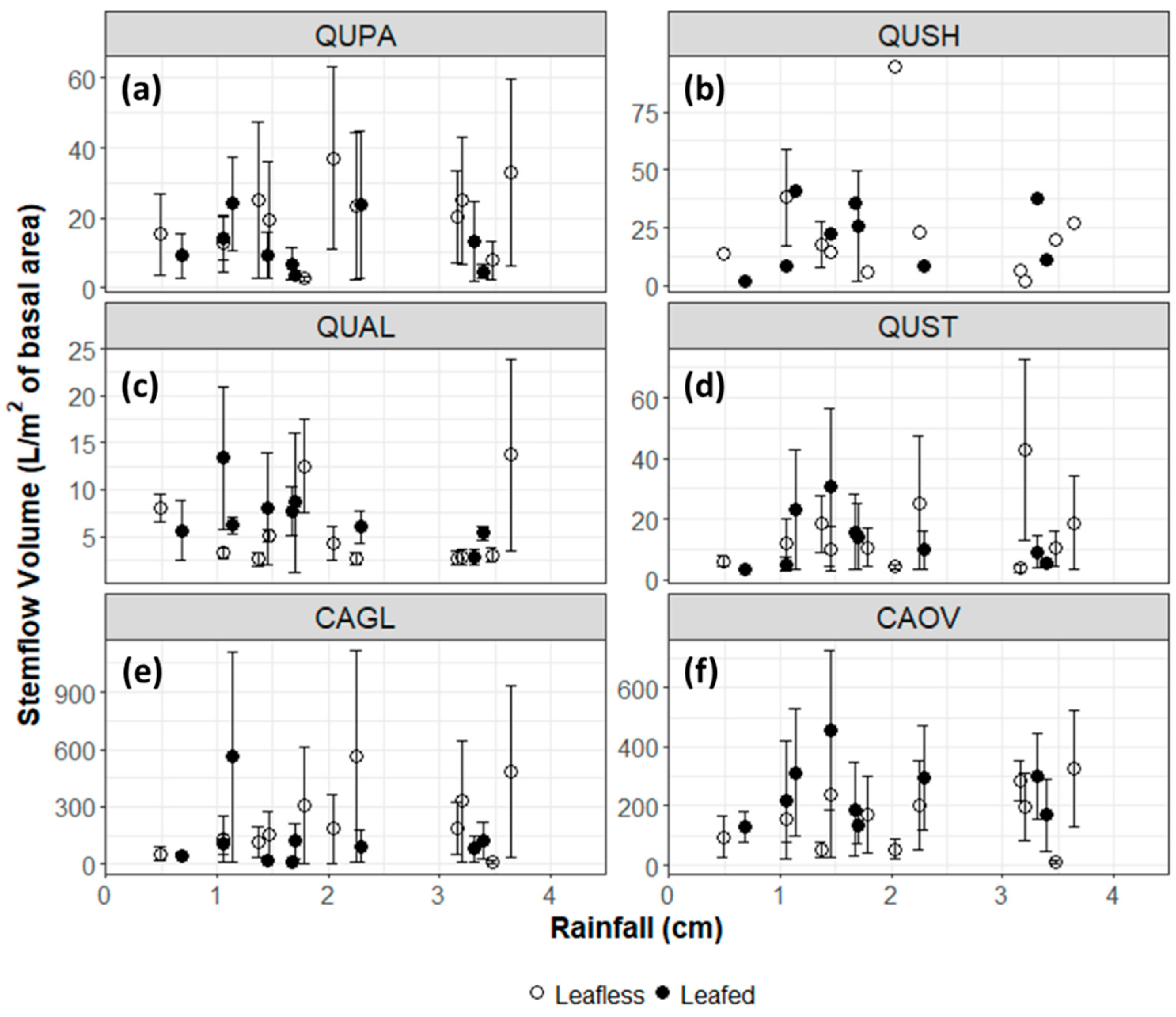
3.1. Throughfall and Stemflow Hydrology
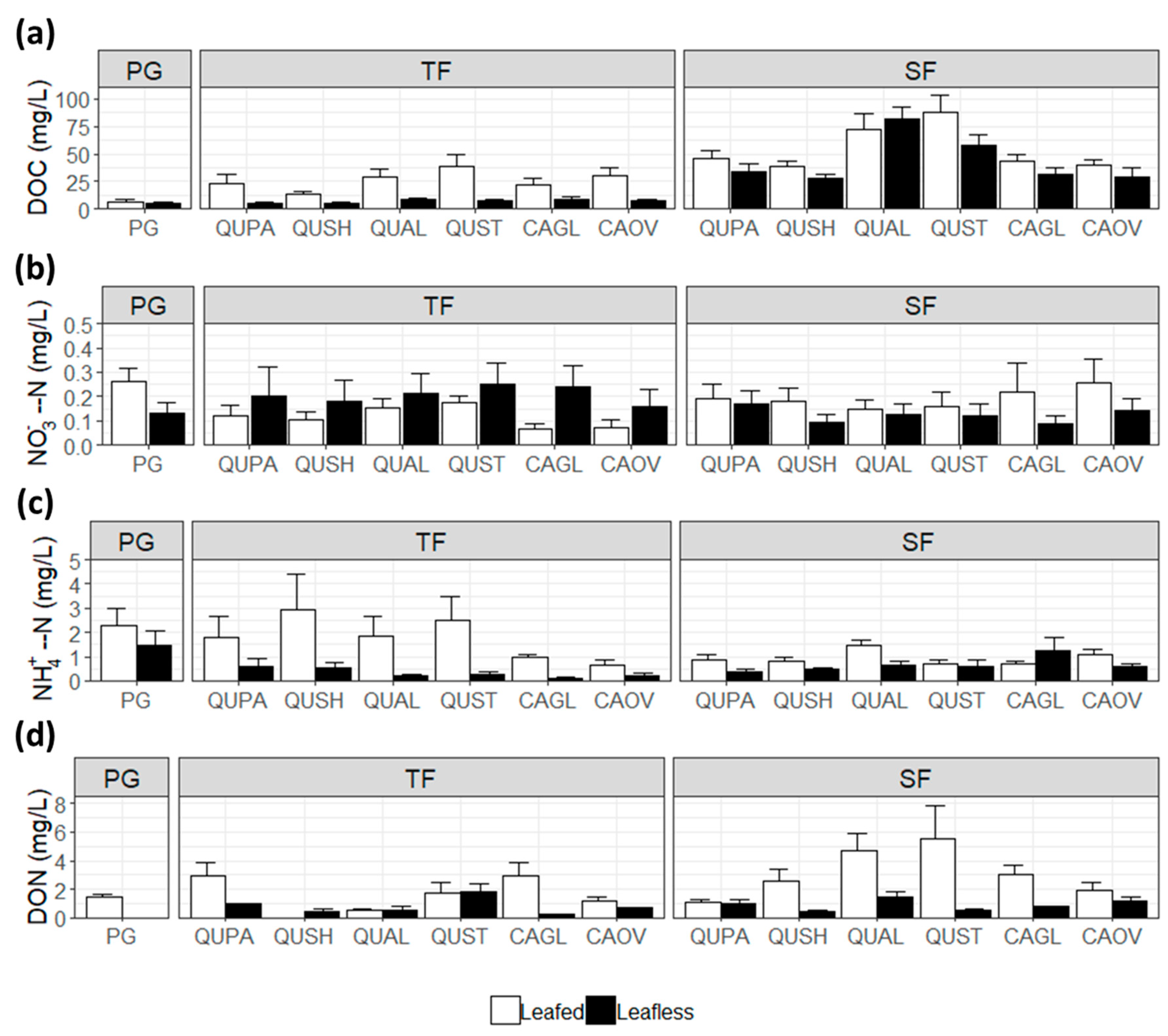
3.2. Throughfall and Stemflow DOC
3.3. Throughfall and Stemflow Inorganic Nitrogen
3.4. Throughfall and Stemflow Organic Nitrogen
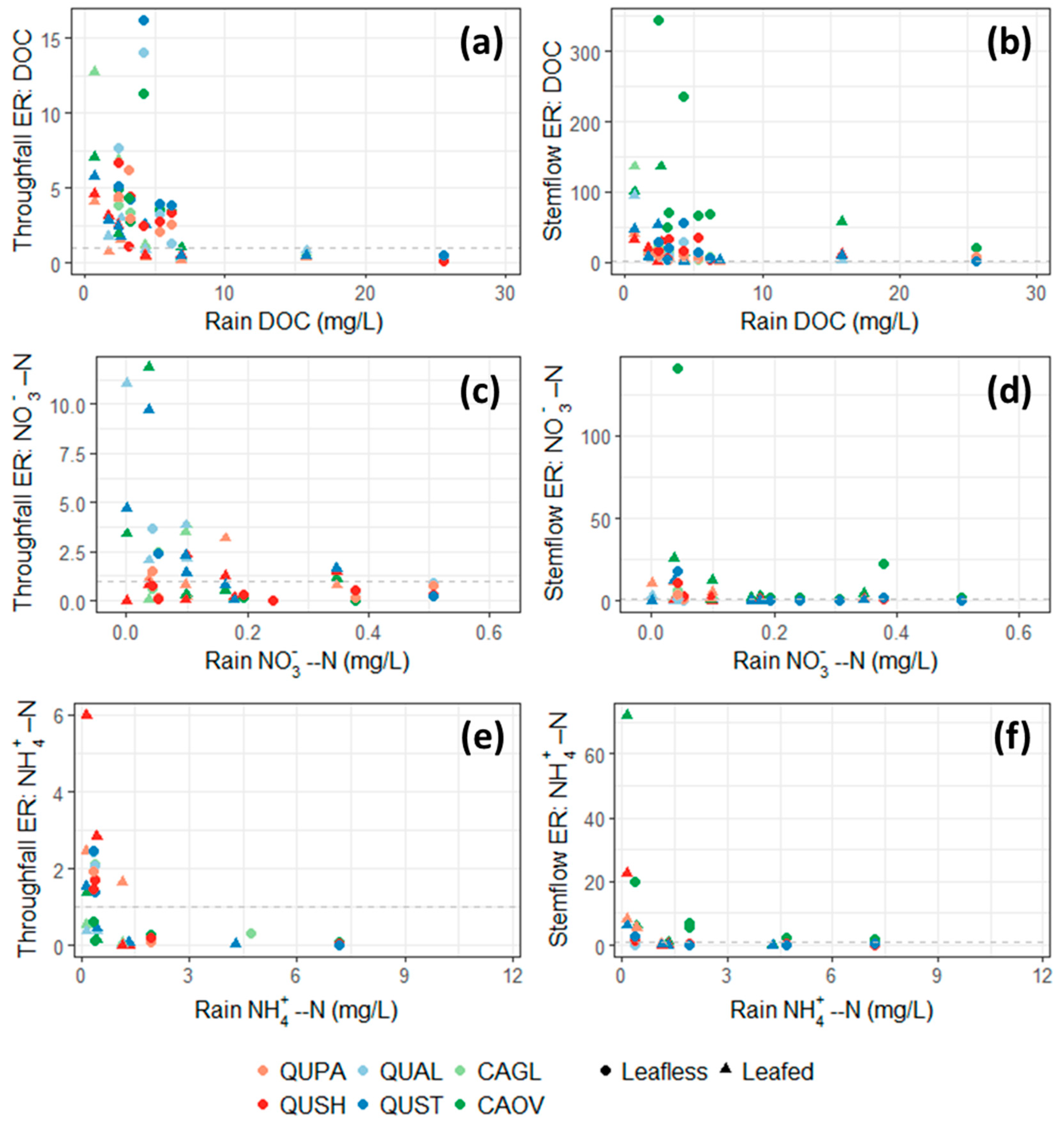
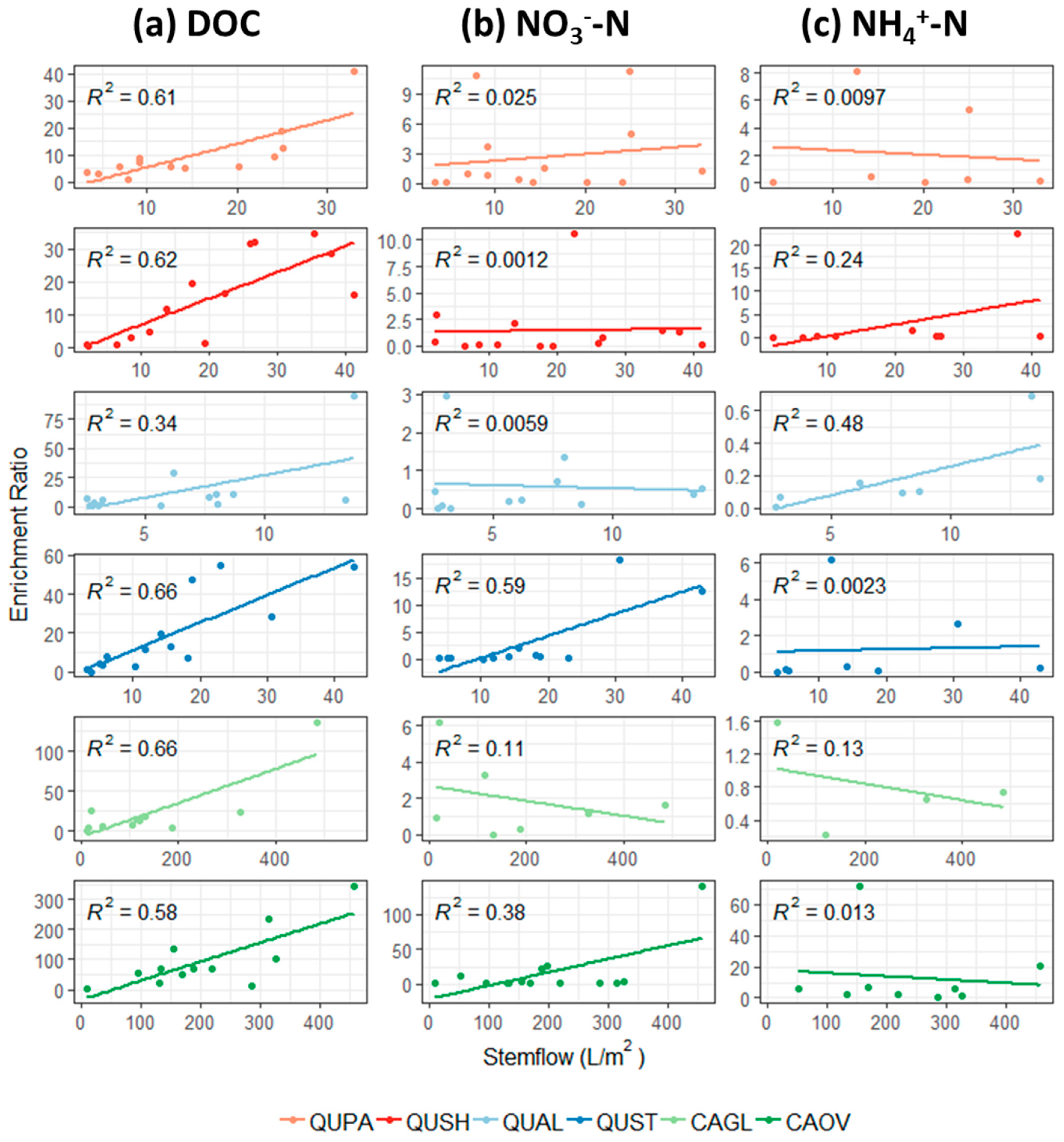
3.5. Throughfall and Stemflow DOC Enrichment
3.6. Throughfall and Stemflow Inorganic Nitrogen Enrichment
4. Discussion
4.1. Throughfall and Stemflow Hydrology
4.2. Throughfall and Stemflow Chemistry
4.2.1. Variations between Precipitation, Throughfall, and Stemflow
4.2.2. Interspecific Differences in Throughfall and Stemflow Chemistry
4.2.3. Seasonal Differences in Throughfall and Stemflow Chemistry
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Groot, R.S.; Wilson, M.A.; Boumans, R.M. A typology for the classification, description and valuation of ecosystem functions, goods and services. Ecol. Econ. 2002, 41, 393–408. [Google Scholar] [CrossRef] [Green Version]
- Alexander, H.D.; Arthur, M.A. Implications of a predicted shift from upland oaks to red maple on forest hydrology and nutrient availability. Can. J. For. Res. 2010, 40, 716–726. [Google Scholar] [CrossRef]
- Corti, G.; Agnelli, A.; Cocco, S.; Cardelli, V.; Masse, J.; Courchesne, F. Soil affects throughfall and stemflow under Turkey oak (Quercus cerris L.). Geoderma 2019, 333, 43–56. [Google Scholar] [CrossRef]
- Dibb, J.E.; Talbot, R.W.; Scheuer, E.; Seid, G.; DeBell, L.; Lefer, B.; Ridley, B. Stratospheric influence on the northern North American free troposphere during TOPSE: 7Be as a stratospheric tracer. J. Geophys. Res. Atmos. 2003, 108. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.H.; Barker, A.C. An analysis of throughfall and stemflow in mixed oak stands. Water Resour. Res. 1970, 6, 316–323. [Google Scholar] [CrossRef]
- Carlisle, A.; Brown, A.H.F.; White, E.J. The nutrient content of tree stem flow and ground flora litter and leachates in a Sessile oak (Quercus Petraea) woodland. J. Ecol. 1967, 55, 615–627. [Google Scholar] [CrossRef]
- Abrams, M.D. Where has all the white oak gone? BioScience 2003, 53, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Alexander, H.D.; Arthur, M.A. Increasing red maple leaf litter alters decomposition rates and nitrogen cycling in historically oak-dominated forests of the eastern U.S. Ecosystems 2014, 17, 1371–1383. [Google Scholar] [CrossRef]
- Hutchinson, T.F.; Long, R.P.; Ford, R.D.; Sutherland, E.K. Fire history and the establishment of oaks and maples in second-growth forests. Can. J. For. Res. 2008, 38, 1184–1198. [Google Scholar] [CrossRef] [Green Version]
- Waldrop, M.P.; Zak, D.R.; Sinsabaugh, R.L. Microbial community response to nitrogen deposition in northern forest ecosystems. Soil Biol. Biochem. 2004, 36, 1443–1451. [Google Scholar] [CrossRef]
- BassiriRad, H.; Lussenhop, J.F.; Sehtiya, H.L.; Borden, K.K. Nitrogen deposition potentially contributes to oak regeneration failure in the Midwestern temperate forests of the USA. Oecologia 2015, 177, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Hanberry, B.B.; Abrams, M.D. Does white-tailed deer density affect tree stocking in forests of the Eastern United States? Ecol. Process. 2019, 8, 30. [Google Scholar] [CrossRef] [Green Version]
- Cote, S.D.; Rooney, T.P.; Tremblay, J.P.; Dussault, C.; Waller, D.M. Ecological impacts of deer overabundance. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 113–147. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Liang, J.; Cumming, J.R.; Lee, E.; Welsh, A.B.; Watson, J.V.; Zhou, M. Fundamental shifts of central hardwood forests under climate change. Ecol. Model. 2016, 332, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Nowacki, G.J.; Abrams, M.D. The demise of fire and “mesophication” of forests in the eastern United States. BioScience 2008, 58, 123–138. [Google Scholar] [CrossRef]
- Olson, M.G.; Stevenson, A.P.; Knapp, B.O.; Kabrick, J.M.; Jensen, R.G. Is there evidence of mesophication of oak forests in the Missouri Ozarks? In Proceedings of the 19th Central Hardwood Forest Conference, Carbondale, IL, USA, 10–12 March 2014; Groninger, J.W., Holzmueller, E.J., Nielsen, C.K., Dey, D.C., Eds.; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2014; pp. 139–153. [Google Scholar]
- Chapman, J.I.; McEwan, R.W. Thirty years of compositional change in an old-growth temperate forest: The role of topographic gradients in oak-maple dynamics. PLoS ONE 2016, 11, e0160238. [Google Scholar] [CrossRef]
- Kreye, J.K.; Varner, J.M.; Hamby, G.W.; Kane, J.M. Mesophytic litter dampens flammability in fire-excluded pyrophytic oak-hickory woodlands. Ecosphere 2018, 9, e02078. [Google Scholar] [CrossRef]
- Palus, J.D.; Goebel, P.C.; Hix, D.M.; Matthews, S.N. Structural and compositional shifts in forests undergoing mesophication in the Wayne National Forest, southeastern Ohio. For. Ecol. Manag. 2018, 430, 413–420. [Google Scholar] [CrossRef]
- Abrams, M.D. Fire and the development of oak forests. BioScience 1992, 42, 346–353. [Google Scholar] [CrossRef]
- Adriaenssens, S.; Hansen, K.; Staelens, J.; Wuyts, K.; De Schrijver, A.; Baeten, L.; Boeckx, P.; Samson, R.; Verheyen, K. Throughfall deposition and canopy exchange processes along a vertical gradient within the canopy of beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst). Sci. Total Environ. 2012, 420, 168–182. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Levia, D.F.; Inamdar, S.P.; Lepori-Bui, M.; Mitchell, M.J. The effects of phenoseason and storm characteristics on throughfall solute washoff and leaching dynamics from a temperate deciduous forest canopy. Sci. Total Environ. 2012, 430, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Siegert, C.M.; Drotar, N.A.; Alexander, H.D. Spatial and temporal variability of throughfall among oak and co-occurring non-oak tree species in an upland hardwood forest. Geosciences 2019, 9, 405. [Google Scholar] [CrossRef] [Green Version]
- Levia, D.F.; Frost, E.E. Variability of throughfall volume and solute inputs in wooded ecosystems. Prog. Phys. Geogr. 2006, 30, 605–632. [Google Scholar] [CrossRef]
- Levia, D.F.; Germer, S. A review of stemflow generation dynamics and stemflow-environment interactions in forests and shrublands. Rev. Geophys. 2015, 53, 673–714. [Google Scholar] [CrossRef]
- Carlyle-Moses, D.E.; Flores Laureano, J.S.; Price, A.G. Throughfall and throughfall spatial variability in Madrean oak forest communities of northeastern Mexico. J. Hydrol. 2004, 297, 124–135. [Google Scholar] [CrossRef]
- Bhat, S.; Jacobs, J.M.; Bryant, M.L. The chemical composition of rainfall and throughfall in five forest communities: A case study in Fort Benning, Georgia. Water Air Soil Pollut. 2011, 218, 323–332. [Google Scholar] [CrossRef]
- Bryant, M.L.; Bhat, S.; Jacobs, J.M. Measurements and modeling of throughfall variability for five forest communities in the southeastern US. J. Hydrol. 2005, 312, 95–108. [Google Scholar] [CrossRef]
- André, F.; Jonard, M.; Jonard, F.; Ponette, Q. Spatial and temporal patterns of throughfall volume in a deciduous mixed-species stand. J. Hydrol. 2011, 400, 244–254. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Siegert, C.M.; Levia, D.F.; Scheick, C.E. Effects of wind-driven rainfall on stemflow generation between codominant tree species with differing crown characteristics. Agric. For. Meteorol. 2011, 151, 1277–1286. [Google Scholar] [CrossRef]
- Siegert, C.M.; Levia, D.F. Seasonal and meteorological effects on differential stemflow funneling ratios for two deciduous tree species. J. Hydrol. 2014, 519, 446–454. [Google Scholar] [CrossRef]
- Duchesne, L.; Ouimet, R.; Camiré, C.; Houle, D. Seasonal nutrient transfers by foliar resorption, leaching, and litter fall in a northern hardwood forest at Lake Clair Watershed, Quebec, Canada. Can. J. For. Res. 2001, 31, 333–344. [Google Scholar] [CrossRef]
- André, F.; Jonard, M.; Ponette, Q. Effects of biological and meteorological factors on stemflow chemistry within a temperate mixed oak–beech stand. Sci. Total Environ. 2008, 393, 72–83. [Google Scholar] [CrossRef]
- Wuyts, K.; Adriaenssens, S.; Staelens, J.; Wuytack, T.; Van Wittenberghe, S.; Boeckx, P.; Samson, R.; Verheyen, K. Contributing factors in foliar uptake of dissolved inorganic nitrogen at leaf level. Sci. Total Environ. 2015, 505, 992–1002. [Google Scholar] [CrossRef]
- André, F.; Jonard, M.; Ponette, Q. Spatial and temporal patterns of throughfall chemistry within a temperate mixed oak–beech stand. Sci. Total Environ. 2008, 397, 215–228. [Google Scholar] [CrossRef]
- Natural Resources Conservation Service Web Soil Survey. Available online: https://websoilsurvey.sc.egov.usda.gov/App/HomePage.htm (accessed on 15 October 2019).
- Arguez, A.; Durre, I.; Applequist, S.; Squires, M.; Vose, R.; Yin, X.; Bilotta, R. NOAA’s U.S. Climate Normals (1981-2010); National Centers for Environmental Information: Asheville, NC, USA, 2010.
- Keim, R.F.; Skaugset, A.E.; Weiler, M. Temporal persistence of spatial patterns in throughfall. J. Hydrol. 2005, 314, 263–274. [Google Scholar] [CrossRef]
- Robertson, S.M.C.; Hornung, M.; Kennedy, V.H. Water chemistry of throughfall and soil water under four tree species at Gisburn, northwest England, before and after felling. For. Ecol. and Manag. 2000, 129, 101–117. [Google Scholar] [CrossRef]
- Levia, D.F.; Herwitz, S.R. Physical properties of water in relation to stemflow leachate dynamics: Implications for nutrient cycling. Can. J. For. Res. 2000, 30, 662–666. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Marin, C.T.; Bouten, W.; Sevink, J. Gross rainfall and its partitioning into throughfall, stemflow and evaporation of intercepted water in four forest ecosystems in western Amazonia. J. Hydrol. 2000, 237, 40–57. [Google Scholar] [CrossRef]
- Andre, F.; Jonard, M.; Ponette, Q. Influence of species and rain event characteristics on stemflow volume in a temperate mixed oak-beech stand. Hydrol. Process. 2008, 22, 4455–4466. [Google Scholar] [CrossRef]
- Siegle-Gaither, M. Using Deuterium and Oxygen-18 Isotopes to Better Understand Stemflow Generation Mechanisms; Mississippi State University: Starkville, MS, USA, 2017. [Google Scholar]
- Crockford, R.H.; Richardson, D.P. Partitioning of rainfall into throughfall, stemflow and interception: Effect of forest type, ground cover and climate. Hydrol. Process. 2000, 14, 2903–2920. [Google Scholar] [CrossRef]
- Levia, D.F.; Frost, E.E. A review and evaluation of stemflow literature in the hydrologic and biogeochemical cycles of forested and agricultural ecosystems. J. Hydrol. 2003, 274, 1–29. [Google Scholar] [CrossRef]
- Thomas, R.; Bennett, N. Estimating bark thicknesses of common Appalachian hardwoods. In Proceedings of the 19th Central Hardwood Forest Conference, Carbondale, IL, USA, 10–12 March 2014; Groninger, J., Holzmueller, E., Nielsen, C., Dey, D., Eds.; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2014; p. 388. [Google Scholar]
- Hengst, G.E.; Dawson, J.O. Bark properties and fire resistance of selected tree species from the central hardwood region of North America. Can. J. For. Res. 1994, 24, 688–696. [Google Scholar] [CrossRef]
- Hammond, D.H.; Varner, J.M.; Kush, J.S.; Fan, Z. Contrasting sapling bark allocation of five southeastern USA hardwood tree species in a fire prone ecosystem. Ecosphere 2015, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Link, T.E.; Unsworth, M.; Marks, D. The dynamics of rainfall interception by a seasonal temperate rainforest. Agric. For. Meteorol. 2004, 124, 171–191. [Google Scholar] [CrossRef] [Green Version]
- Pypker, T.G.; Unsworth, M.H.; Bond, B.J. The role of epiphytes in rainfall interception by forests in the Pacific Northwest. II. Field measurements at the branch and canopy scale. Can. J. For. Res. 2006, 36, 819–832. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-P.; Wang, Z.-N.; Berndtsson, R.; Zhang, Y.-F.; Pan, Y.-X. Desert shrub stemflow and its significance in soil moisture replenishment. Hydrol. Earth Syst. Sci. 2011, 15, 561–567. [Google Scholar] [CrossRef] [Green Version]
- Silva, I.C.; Okumura, T. Throughfall, stemflow and interception loss in a mixed white oak forest (Quercus serrata Thunb.). J. For. Res. 1996, 1, 123–129. [Google Scholar] [CrossRef]
- Staelens, J.; De Schrijver, A.; Verheyen, K. Seasonal variation in throughfall and stemflow chemistry beneath a European beech (Fagus sylvatica) tree in relation to canopy phenology. Can. J. For. Res. 2007, 37, 1359–1372. [Google Scholar] [CrossRef]
- Hörmann, G.; Branding, A.; Clemen, T.; Herbst, M.; Hinrichs, A.; Thamm, F. Calculation and simulation of wind controlled canopy interception of a beech forest in Northern Germany. Agric. For. Meteorol. 1996, 79, 131–148. [Google Scholar] [CrossRef]
- Verstraeten, A.; Neirynck, J.; Genouw, G.; Cools, N.; Roskams, P.; Hens, M. Impact of declining atmospheric deposition on forest soil solution chemistry in Flanders, Belgium. Atmos. Environ. 2012, 62, 50–63. [Google Scholar] [CrossRef]
- Staelens, J.; De Schrijver, A.; Verheyen, K.; Verhoest, N.E. Spatial variability and temporal stability of throughfall water under a dominant beech (Fagus sylvatica L.) tree in relationship to canopy cover. J. Hydrol. 2006, 330, 651–662. [Google Scholar] [CrossRef]
- Staelens, J.; De Schrijver, A.; Verheyen, K.; Verhoest, N.E. Rainfall partitioning into throughfall, stemflow, and interception within a single beech (Fagus sylvatica L.) canopy: Influence of foliation, rain event characteristics, and meteorology. Hydrol. Process. 2008, 22, 33–45. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Pan, Y.; Hu, R. Relative contribution of biotic and abiotic factors to stemflow production and funneling efficiency: A long-term field study on a xerophytic shrub species in Tengger Desert of northern China. Agric. For. Meteorol. 2020, 280, 107781. [Google Scholar] [CrossRef]
- Skau, C.M. Interception, throughfall, and stemflow in Utah and Alligator Juniper cover types of northern Arizona. For. Sci. 1964, 10, 283–287. [Google Scholar]
- Siegert, C.M.; Levia, D.F.; Leathers, D.J.; Van Stan, J.T.; Mitchell, M.J. Do storm synoptic patterns affect biogeochemical fluxes from temperate deciduous forest canopies? Biogeochemistry 2017, 132, 273–292. [Google Scholar] [CrossRef]
- Potter, C.S.; Ragsdale, H.L.; Swank, W.T. Atmospheric deposition and foliar leaching in a regenerating southern Appalachian forest canopy. J. Ecol. 1991, 79, 97–115. [Google Scholar] [CrossRef]
- Tukey, H.B. The leaching of substances from plants. Annu. Rev. Plant Physiol. 1970, 21, 305–324. [Google Scholar] [CrossRef]
- Talkner, U.; Krämer, I.; Hölscher, D.; Beese, F.O. Deposition and canopy exchange processes in central-German beech forests differing in tree species diversity. Plant Soil 2010, 336, 405–420. [Google Scholar] [CrossRef] [Green Version]
- Lovett, G.M.; Nolan, S.S.; Driscoll, C.T.; Fahey, T.J. Factors regulating throughfall flux in a New Hampshire forested landscape. Can. J. For. Res. 1996, 26, 2134–2144. [Google Scholar] [CrossRef]
- Whelan, M.J.; Sanger, L.J.; Baker, M.; Anderson, J.M. Spatial patterns of throughfall and mineral ion deposition in a lowland Norway spruce (Picea abies) plantation at the plot scale. Atmos. Environ. 1998, 32, 3493–3501. [Google Scholar] [CrossRef]
- Staelens, J.; De Schrijver, A.; Verheyen, K.; Verhoest, N.E. Spatial variability and temporal stability of throughfall deposition under beech (Fagus sylvatica L.) in relationship to canopy structure. Environ. Pollut. 2006, 142, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Lovett, G.M.; Lindberg, S.E. Dry deposition and canopy exchange in a mixed oak forest as determined by analysis of throughfall. J. Appl. Ecol. 1984, 21, 1013–1027. [Google Scholar] [CrossRef]
- Hamburg, S.P.; Lin, T.-C. Throughfall chemistry of an ecotonal forest on the edge of the Great Plains. Can. J. For. Res. 1998, 28, 1456–1463. [Google Scholar] [CrossRef]
- Cape, J.N.; Sheppard, L.J.; Crossley, A.; van Dijk, N.; Tang, Y.S. Experimental field estimation of organic nitrogen formation in tree canopies. Environ. Pollut. 2010, 158, 2926–2933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, L.; Zhao, C.; Xu, W.; Xie, Z. Hydrochemical fluxes in bulk precipitation, throughfall, and stemflow in a mixed evergreen and deciduous broadleaved forest. Forests 2019, 10, 507. [Google Scholar] [CrossRef] [Green Version]
- Guerrieri, R.; Vanguelova, E.I.; Michalski, G.; Heaton, T.H.E.; Mencuccini, M. Isotopic evidence for the occurrence of biological nitrification and nitrogen deposition processing in forest canopies. Glob. Chang. Biol. 2015, 21, 4613–4626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrieri, R.; Lecha, L.; Mattana, S.; Cáliz, J.; Casamayor, E.O.; Barceló, A.; Michalski, G.; Peñuelas, J.; Avila, A.; Mencuccini, M. Partitioning between atmospheric deposition and canopy microbial nitrification into throughfall nitrate fluxes in a Mediterranean forest. J. Ecol. 2019, 00, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Le Mellec, A.; Meesenburg, H.; Michalzik, B. The importance of canopy-derived dissolved and particulate organic matter (DOM and POM)—Comparing throughfall solution from broadleaved and coniferous forests. Ann. For. Sci. 2010, 67, 411. [Google Scholar] [CrossRef]
- Verstraeten, A.; Verschelde, P.; De Vos, B.; Neirynck, J.; Cools, N.; Roskams, P.; Hens, M.; Louette, G.; Sleutel, S.; De Neve, S. Increasing trends of dissolved organic nitrogen (DON) in temperate forests under recovery from acidification in Flanders, Belgium. Sci. Total Environ. 2016, 553, 107–119. [Google Scholar] [CrossRef]
- Fan, H.B.; Hong, W. Estimation of dry deposition and canopy exchange in Chinese fir plantations. For. Ecol. Manag. 2001, 147, 99–107. [Google Scholar] [CrossRef]
- Moreno, G.; Gallardo, J.F.; Bussotti, F. Canopy modification of atmospheric deposition in oligotrophic Quercus pyrenaica forests of an unpolluted region (central-western Spain). For. Ecol. Manag. 2001, 149, 47–60. [Google Scholar] [CrossRef]
- Mahendrappa, M.K. Chemical composition of stemflow from some eastern Canadian tree species. Can. J. For. Res. 1974, 4, 1–7. [Google Scholar] [CrossRef]
- Chang, S.C.; Matzner, E. The effect of beech stemflow on spatial patterns of soil solution chemistry and seepage fluxes in a mixed beech/oak stand. Hydrol. Process. 2000, 14, 135–144. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Pypker, T.G. A review and evaluation of forest canopy epiphyte roles in the partitioning and chemical alteration of precipitation. Sci. Total Environ. 2015, 536, 813–824. [Google Scholar] [CrossRef]
- Levia, D.F.; Van Stan, J.T.; Siegert, C.M.; Inamdar, S.P.; Mitchell, M.J.; Mage, S.M.; McHale, P.J. Atmospheric deposition and corresponding variability of stemflow chemistry across temporal scales in a mid-Atlantic broadleaved deciduous forest. Atmos. Environ. 2011, 45, 3046–3054. [Google Scholar] [CrossRef]
- Fahey, T.J.; Williams, C.J.; Rooney-Varga, J.N.; Cleveland, C.C.; Postek, K.M.; Smith, S.D.; Bouldin, D.R. Nitrogen deposition in and around an intensive agricultural district in central New York. J. Environ. Qual. 1999, 28, 1585–1600. [Google Scholar] [CrossRef]
- Herrmann, M.; Pust, J.; Pott, R. The chemical composition of throughfall beneath oak, birch and pine canopies in Northwest Germany. Plant Ecol. 2006, 184, 273–285. [Google Scholar] [CrossRef]
- Qualls, R.G.; Haines, B.L. Geochemistry of dissolved organic nutrients in water percolating through a forest ecosystem. Soil Sci. Soc. Am. J. 1991, 55, 1112–1123. [Google Scholar] [CrossRef] [Green Version]
- Augusto, L.; Dupouey, J.; Ranger, J. Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann. For. Sci. 2003, 60, 823–831. [Google Scholar] [CrossRef]
- Pitman, R.M.; Vanguelova, E.I.; Benham, S.E. The effects of phytophagous insects on water and soil nutrient concentrations and fluxes through forest stands of the Level II monitoring network in the UK. Sci. Total Environ. 2010, 409, 169–181. [Google Scholar] [CrossRef]
- Cardelús, C.L.; Mack, M.C.; Woods, C.; DeMarco, J.; Treseder, K.K. The influence of tree species on canopy soil nutrient status in a tropical lowland wet forest in Costa Rica. Plant Soil 2009, 318, 47–61. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Van Stan, J.H.; Levia, D.F. Meteorological influences on stemflow generation across diameter size classes of two morphologically distinct deciduous species. Int. J. Biometeorol. 2014, 58, 2059–2069. [Google Scholar] [CrossRef] [PubMed]
- Stubbins, A.; Silva, L.M.; Dittmar, T.; Van Stan, J.T. Molecular and optical properties of tree-derived dissolved organic matter in throughfall and stemflow from live oaks and eastern red cedar. Front. Earth Sci. 2017, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Fox, J.E.D.; Xu, Z. Nutrient fluxes in bulk precipitation, throughfall and stemflow in montane subtropical moist forest on Ailao Mountains in Yunnan, south-west China. J. Trop. Ecol. 2002, 18, 527–548. [Google Scholar] [CrossRef]
- Lovett, G.M.; Lindberg, S.E. Dry deposition of nitrate to a deciduous forest. Biogeochemistry 1986, 2, 137–148. [Google Scholar] [CrossRef]
- Fowler, D.; Cape, J.N.; Unsworth, M.H.; Mayer, H.; Crowther, J.M.; Jarvis, P.G.; Gardiner, B.; Shuttleworth, W.J. Deposition of atmospheric pollutants on forests. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1989, 324, 247–265. [Google Scholar] [CrossRef]
- Morris, D.M.; Gordon, A.G.; Gordon, A.M. Patterns of canopy interception and throughfall along a topographic sequence for black spruce dominated forest ecosystems in northwestern Ontario. Can. J. For. Res. 2003, 33, 1046–1060. [Google Scholar] [CrossRef]
- Puckett, L.J. Estimates of ion sources in deciduous and coniferous throughfall. Atmos. Environ. 1990, 24, 545–555. [Google Scholar] [CrossRef]
- Hafner, S.D.; Groffman, P.M. Soil nitrogen cycling under litter and coarse woody debris in a mixed forest in New York State. Soil Biol. Biochem. 2005, 37, 2159–2162. [Google Scholar] [CrossRef]
- Neary, A.J.; Gizyn, W.I. Throughfall and stemflow chemistry under deciduous and coniferous forest canopies in South Central Ontario. Can. J. For. Res. 1994, 24, 1089–1100. [Google Scholar] [CrossRef]
- Foster, N.W.; Nicolson, J.A. Acid deposition and nutrient leaching from deciduous vegetation and podzolic soils at the Turkey Lakes watershed. Can. J. Fish. Aquat. Sci. 1988, 45, s96–s100. [Google Scholar] [CrossRef]
- Jo, I.; Fei, S.; Oswalt, C.M.; Domke, G.M.; Phillips, R.P. Shifts in dominant tree mycorrhizal associations in response to anthropogenic impacts. Sci. Adv. 2019, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species | Species Code | DBH (cm) | Crown Area (m2) | Tree Height (m) | Bark Thickness (cm) | |
|---|---|---|---|---|---|---|
| Shumard oak | Quercus shumardii | QUSH | 65.4 | 47.6 | 40.7 | 0.94 |
| Cherrybark oak | Quercus pagoda | QUPA | 68.6 | 179.1 | 48.0 | 1.07 |
| Post oak | Quercus stellata | QUST | 59.1 | 147.4 | 25.2 | 1.35 |
| White oak | Quercus alba | QUAL | 66.6 | 132.6 | 45.1 | 1.67 |
| Pignut hickory | Carya glabra | CAGL | 43.9 | 76.6 | 23.1 | 1.13 |
| Shagbark hickory | Carya ovata | CAOV | 35.7 | 44.4 | 21.8 | 0.64 |
| Leafed | Leafless | |||||
|---|---|---|---|---|---|---|
| DOC | NO3− | NH4+ | DOC | NO3− | NH4+ | |
| THROUGHFALL | ||||||
| Cherrybark Oak | 3.66 | 0.59 | 0.93 | 1.68 | 0.88 | 0.57 |
| Shumard Oak | 3.00 | 0.35 | 0.85 | 2.19 | 0.88 | 0.59 |
| White Oak | 5.17 | 1.27 | 1.52 | 1.55 | 3.33 | 0.10 |
| Post Oak | 5.65 | 1.32 | 1.29 | 2.35 | 2.94 | 0.14 |
| Pignut Hickory | 2.87 | 1.52 | 1.22 | 4.12 | 33.82 | 0.03 |
| Shagbark Hickory | 4.36 | 1.29 | 0.28 | 2.10 | 2.50 | 0.04 |
| Average | 4.18 | 0.89 | 1.00 | 2.31 | 1.99 | 0.75 |
| STEMFLOW | ||||||
| Cherrybark Oak | 6.07 | 0.81 | 0.26 | 14.03 | 4.32 | 2.78 |
| Shumard Oak | 15.32 | 2.20 | 0.49 | 13.55 | 0.66 | 5.72 |
| White Oak | 10.74 | 0.50 | 0.26 | 16.18 | 0.67 | 0.08 |
| Post Oak | 17.87 | 3.48 | 0.76 | 18.70 | 2.27 | 1.61 |
| Pignut Hickory | 10.96 | 3.52 | 0.90 | 32.67 | 1.27 | 0.70 |
| Shagbark Hickory | 122.12 | 24.56 | 7.35 | 61.06 | 7.26 | 19.86 |
| Average | 32.02 | 6.40 | 2.07 | 24.33 | 2.88 | 5.65 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Limpert, K.; Siegert, C. Interspecific Differences in Canopy-Derived Water, Carbon, and Nitrogen in Upland Oak-Hickory Forest. Forests 2019, 10, 1121. https://doi.org/10.3390/f10121121
Limpert K, Siegert C. Interspecific Differences in Canopy-Derived Water, Carbon, and Nitrogen in Upland Oak-Hickory Forest. Forests. 2019; 10(12):1121. https://doi.org/10.3390/f10121121
Chicago/Turabian StyleLimpert, Katy, and Courtney Siegert. 2019. "Interspecific Differences in Canopy-Derived Water, Carbon, and Nitrogen in Upland Oak-Hickory Forest" Forests 10, no. 12: 1121. https://doi.org/10.3390/f10121121
APA StyleLimpert, K., & Siegert, C. (2019). Interspecific Differences in Canopy-Derived Water, Carbon, and Nitrogen in Upland Oak-Hickory Forest. Forests, 10(12), 1121. https://doi.org/10.3390/f10121121