An Ant-Plant Mesocosm Experiment Reveals Dispersal Patterns of Myrmecochorous Plants

Abstract
:1. Introduction
2. Methods and Materials
2.1. Study Species
2.2. Mesocosms
2.3. Statistical Analyses
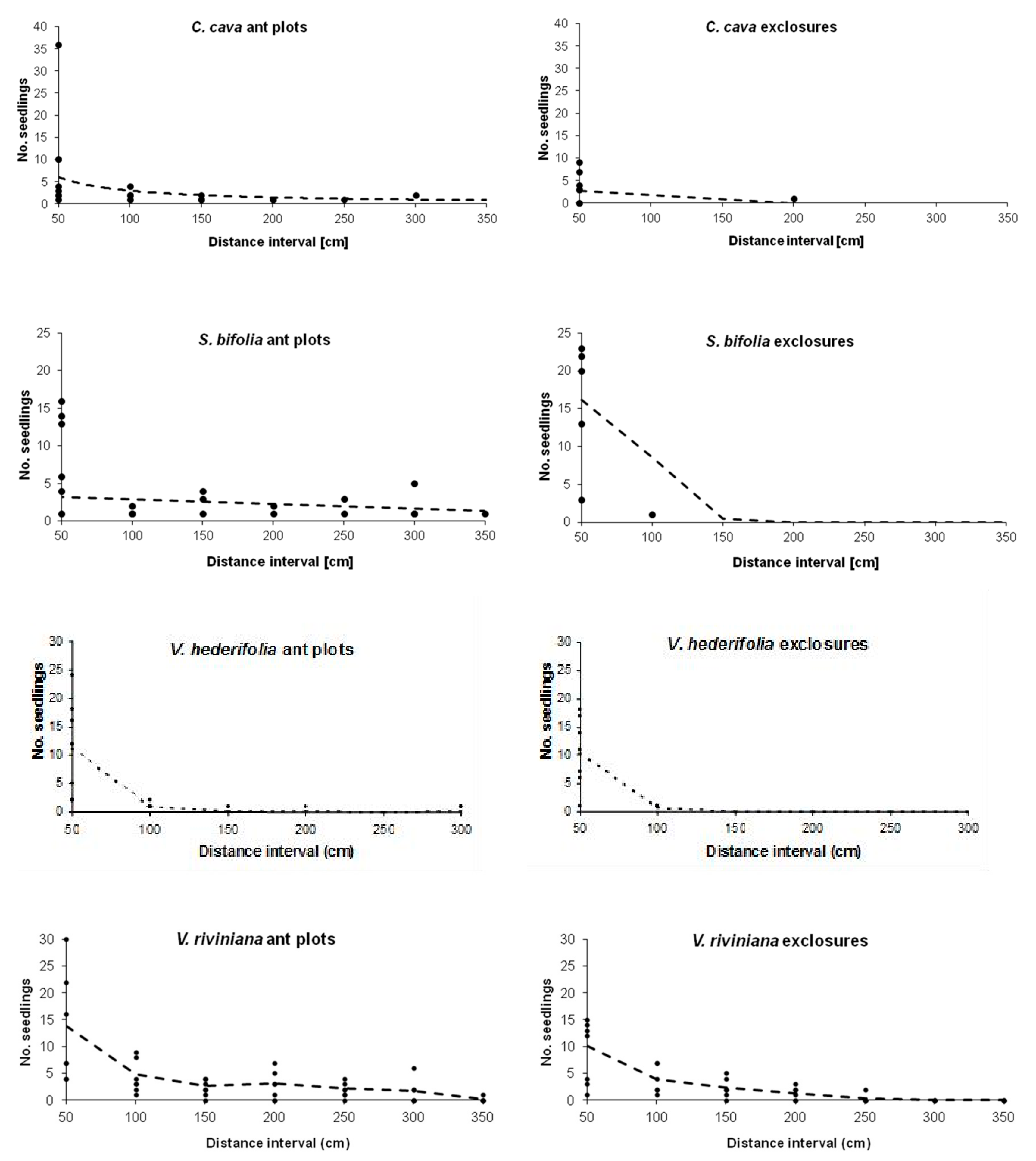
3. Results
3.1. Corydalis cava
3.2. Scilla bifolia
3.3. Veronica hederifolia
3.4. Viola riviniana
4. Discussion
4.1. Spatial Seed Dispersal Pattern and Distances
4.1.1. Corydalis cava and Scilla bifolia
4.1.2. Veronica hederifolia
4.1.3. Viola riviniana
4.2. Dispersal Distance
4.3. Directed Dispersal
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Giladi, I. Choosing benefits or partners: A review of the evidence for the evolution of myrmecochory. Oikos 2006, 112, 481–492. [Google Scholar] [CrossRef] [Green Version]
- Beattie, A.J. The Evolutionary Ecology of Ant-Plant Mutualisms; Cambridge University Press: Cambridge, UK, 1985. [Google Scholar]
- Morales, M.A.; Heithaus, E.R. Food from seed-dispersal mutualism shifts sex ratios in colonies of the ant Aphaenogaster rudis. Ecology 1998, 79, 734–739. [Google Scholar] [CrossRef] [Green Version]
- Gammans, N.; Bullock, J.; Schönrogge, K. Ant benefits in a seed dispersal mutualism. Oecologia 2005, 146, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Fokuhl, G.; Heinze, J.; Poschlod, P. Colony growth in Myrmica rubra with supplementation of myrmecochorous seeds. Ecol. Res. 2007, 22, 845–847. [Google Scholar] [CrossRef]
- Warren II, R.J.; Elliott, K.J.; Giladi, I.; King, J.R.; Bradford, M.A. Field experiments show contradictory short- and long-term myrmecochorous plant impacts on seed-dispersing ants. Ecol. Entomol. 2019, 44, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Vander Wall, S.B.; Longland, S.L. Diplochory: Are two seed dispersers better than one? Trends Ecol. Evol. 2004, 19, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Gomez, C.; Espadaler, X. An update of the world survey of myrmecochorous dispersal distances. Ecography 2013, 36, 1193–1201. [Google Scholar] [CrossRef]
- Chen, S.-C.; Tamme, R.; Thomson, F.J.; Moles, A.T. Seeds tend to disperse further in the tropics. Ecol. Lett. 2019, 22, 954–961. [Google Scholar] [CrossRef]
- Heinken, T. Migration of an annual myrmecochore: A four year experiment with Melampyrum pratense L. Plant Ecol. 2004, 170, 55–72. [Google Scholar] [CrossRef] [Green Version]
- Andersen, A.N. Dispersal distance as a benefit of myrmecochory. Oecologia 1988, 75, 507–511. [Google Scholar] [CrossRef]
- Higashi, S.; Tsuyuzaki, S.; Ohara, M.; Ito, F. Adaptive advantages of ant-dispersed seeds in the myrmecochorous plant Trillium tschonoskii (Liliaceae). Oikos 1989, 54, 389–394. [Google Scholar] [CrossRef]
- Oostermeijer, J.G.B. Myrmecochory in Polygala vulgaris L., Luzula campestris (L.) DC. and Viola curtisii Forster in a Dutch dune area. Oecologia 1989, 78, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Gibson, W. Selective advantages to hemi-parasitic annuals, genus Melampyrum, of a seed-disperal mutualism involving ants 1. Favorable nest sites. Oikos 1993, 67, 334–344. [Google Scholar] [CrossRef]
- Culver, D.C.; Beattie, A. The Fate of Viola Seeds Dispersed by Ants. Am. J. Bot. 1980, 67, 710–714. [Google Scholar] [CrossRef]
- Kjellsson, G. Seed fate in a population of Carex pilulifera L. Oecologia 1985, 67, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.A.; Matlack, G.R. Spatial structure develops early in forest herb populations, controlled by dispersal and life cycle. Oecologia 2019, 189, 951–970. [Google Scholar] [CrossRef]
- Boyd, R.S. Ecological benefits of myrmecochory for the endangered chaparral shrub Fremontodendron decumbens (Sterculiaceae). Am. J. Bot. 2001, 88, 234–241. [Google Scholar] [CrossRef] [Green Version]
- Gorb, S.N.; Gorb, E.V.; Punttila, P. Effects of redispersal of seeds by ants on the vegetation pattern in a deciduous forest: A case study. Acta Oecol. 2000, 21, 293–301. [Google Scholar] [CrossRef]
- Lehouck, V.; Bonte, D.; Dekoninck, W.; Maelfait, J.-P. The distribution of ant nests (Hymenoptera, Formicidae) in coastal grey dunes of Flanders (Belgium) and their relationship to myrmecochorous plants. Belg. J. Zool. 2004, 134, 89–96. [Google Scholar]
- Gorb, S.M.; Gorb, E.B. Removal rates of seeds of five myrmecochorous plants by the ant Formica polyctena (Hymenoptera: Formicidae). Oikos 1995, 73, 367–374. [Google Scholar] [CrossRef]
- Gorb, E.B.; Gorb, S.N. Effects of ant species composition on seed removal in deciduous forest in eastern Europe. Oikos 1999, 84, 110–118. [Google Scholar] [CrossRef]
- Poschlod, P.; Kleyer, M.; Jackel, A.-K.; Dannemann, A.; Tackenberg, O. BIOPOP—A database of plant traits and internet application for nature conservation. Folia Geobot. Et Phytotaxon. 2003, 38, 263–271. [Google Scholar] [CrossRef]
- Collingwood, C.A. The Formicidae (Hymenoptera) of Fennoscandia and Denmark. Fauna Entomol. Scand. 1979, 8, 1–174. [Google Scholar]
- Müller-Schneider, P. Verbreitungsbiologie Der Blütenpflanzen Graubündens; Geobotanisches Institut der ETH, Stiftung Rübel: Zürich, Switzerland, 1986; Volume 85, pp. 1–263. [Google Scholar]
- Clark, P.J.; Evans, F.C. Distance to nearest neighbor as a measure of spatial relationships in populations. Ecology 1954, 35, 445–453. [Google Scholar] [CrossRef]
- Condit, R.; Ashton, P.S.; Baker, P.; Bunyavejchewin, S.; Gunatilleke, S.; Gunatilleke, N.; Hubbell, S.P.; Foster, R.B.; Itoh, A.; LaFrankie, J.V.; et al. Spatial Patterns in the Distribution of Tropical Tree Species. Science 2000, 288, 1414–1418. [Google Scholar] [CrossRef] [PubMed]
- Perry, G.L.W.; Miller, B.P.; Enright, N.J. A comparison of methods for the statistical analysis of spatial point patterns in plant ecology. Plant Ecol. 2006, 187, 59–82. [Google Scholar] [CrossRef]
- Perry, G.L.W. SpPack: Spatial point pattern analysis in Excel using Visual Basic for Applications (VBA). Environ. Model. Softw. 2004, 19, 559–569. [Google Scholar] [CrossRef]
- Oberrath, R.; Böhning-Gaese, K. Phenological adaptation of ant-dispersed plants to seasonal variation in ant activity. Ecology 2002, 83, 1412–1420. [Google Scholar] [CrossRef]
- Cain, M.L.; Damman, H.; Muir, A. Seed dispersal and the holocene migration of woodland herbs. Ecol. Monogr. 1998, 68, 325–347. [Google Scholar] [CrossRef]
- Pakeman, R.J. Plant migration rates and seed dispersal mechanisms. J. Biogeogr. 2001, 28, 795–800. [Google Scholar] [CrossRef]
- Salisbury, E.J. Weeds and Aliens; Collins: London, UK, 1961. [Google Scholar]
- Rothmaler, W. Exkursionsflora Von Deutschland.—Spektrum, Akad; Verlag: Heidelberg, Germany, 1990. [Google Scholar]
- Chen, S.; Pahlevani, A.H.; Malíková, L.; Riina, R.; Thomson, F.J.; Giladi, I. Trade-off or coordination? Correlations between ballochorous and myrmecochorous phases of diplochory. Funct. Ecol. 2019, 33, 1469–1479. [Google Scholar] [CrossRef]
- Beattie, A.; Lyons, N. Seed dispersal in Viola (Violaceae): Adaptations and strategies. Am. J. Bot. 1975, 62, 714–722. [Google Scholar] [CrossRef]
- Zelikova, T.J.; Sanders, N.J.; Dunn, R.R. The mixed effects of experimental ant removal on seedling distribution, belowground invertebrates, and soil nutrients. Ecosphere 2011, 2, 1–14. [Google Scholar] [CrossRef]
- Kalisz, S.; Hanzawa, F.M.; Tonsor, S.J.; Thiede, D.A.; Voigt, S. Ant-mediated seed dispersal alters pattern of relatedness in a population of Trillium grandiflorum. Ecology 1999, 80, 2620–2634. [Google Scholar] [CrossRef] [Green Version]
- Horvitz, C.C.; Le Corff, J. Spatial scale and dispersion pattern of ant- and bird-dispersed herbs in two tropical lowland rain forests. Vegetatio 1993, 107, 351–362. [Google Scholar]
- Petersen, P.M.; Philipp, M. Implantation of forest plants in a wood on former arable land: A ten year experiment. Flora 2001, 196, 286–291. [Google Scholar] [CrossRef]
- Christian, C.E. Consequences of a biological invasion reveal the importance of mutualism for plant communities. Nature 2001, 413, 635–639. [Google Scholar] [CrossRef]
- Ness, J.H. Forest edges and fire ants alter the seed shadow of an ant-dispersed plant. Oecologia 2004, 138, 448–454. [Google Scholar] [CrossRef]
- Böhling, N. Dauerflächen-Untersuchungen in einem Eichen-Hainbuchenwald im Vorland der Schwäbischen Alb (Südwestdeutschland), 1978-2001: Der Niedergang von Scilla bifolia und die Invasion von Allium ursinum. Tuexenia 2003, 23, 131–161. [Google Scholar]
- Ohkawara, K.; Ohara, M.; Higashi, S. The evolution of ant-dispersal in a spring-ephemeral Corydalis ambigua (Papaveraceae): Timing of seed-fall and effects of ant and ground beetles. Ecography 1997, 20, 217–223. [Google Scholar] [CrossRef]
- Bekker, R.M.; Bakker, J.P.; Grandin, U.; Kalamees, R.; Milberg, P.; Poschlod, P.; Thompson, K.; Willems, J.H. Seed size, shape and vertical distribution in the soil: Indicators of seed longevity. Funct. Ecol. 1998, 12, 834–842. [Google Scholar] [CrossRef]
- Gorb, S.N.; Gorb, E.B. Dropping rates of elaiosome-bearing seeds during transport by ants (Formica polyctena Foerst.): Implications for distance dispersal. Acta Oecol. 1999, 20, 509–518. [Google Scholar] [CrossRef]
- Turnbull, C.L. The Dynamics of An Association between Viola nuttallii Pursh and Its Seed Dispersers, Myrmica discontinua Weber and Formica podzolica Francoeur. Ph.D. Thesis, Northwestern University, Evanston, IL, USA, 1985. [Google Scholar]
- Banschbach, V.S.; Herbers, J.M. Nest movements and population spatial structure of the forest ant Myrmica punctiventris (Hymenoptera: Formicidae). Ann. Entomol. Soc. Am. 1999, 92, 414–423. [Google Scholar] [CrossRef]
- Rice, B.; Westoby, M. Evidence against the hypothesis that ant-dispersed seeds reach nutrient-enriched microsites. Ecology 1986, 67, 1270–1274. [Google Scholar] [CrossRef]
- Fokuhl, G.; Heinze, J.; Poschlod, P. Myrmecochory by small ants - Beneficial effects through elaiosome nutrition and seed dispersal. Acta Oecol. 2012, 38, 71–76. [Google Scholar] [CrossRef]
- Türke, M.; Heinze, E.; Andreas, K.; Svendsen, S.M.; Gossner, M.M.; Weisser, W.W. Seed consumption and dispersal in ant-dispersed plants by slugs. Oecologia 2010, 163, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Türke, M.; Andreas, K.; Gossner, M.M.; Kowalski, E.; Lange, M.; Boch, S.; Socher, S.A.; Müller, J.; Prati, D.; Fischer, M.; et al. Are gastropods, rather than ants, important dispersers of seeds of myrmecochorous forest herbs? Am. Nat. 2012, 179, 124–131. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Life form | Life cycle | Time of seed set in experiment | Seed weight (mg)1 | Elaiosome structure |
|---|---|---|---|---|---|
| Hollow root, Corydalis cava (L.) Schweigg. & Körte | geophyte | perennial | May | 10.1 | soft appendage |
| Alpine Squill, Scilla bifolia L. | geophyte | perennial | May | 19.0 | soft appendage |
| Ivy-leaved Speed-well, Veronica hederifolia L. | therophyte | annual | May–June | 4.11 | thickened cell wall |
| Common Dog-violet, Viola riviniana Rchb. | hemicryptophyte | perennial | July–August | 1.43 | dry appendage |
| Species | %Seedlings | n seedlings | Mann-Whitney test | ||||
|---|---|---|---|---|---|---|---|
| Exclosures | Ant plots | Exclosures | Ant plots | n | U | p | |
| Corydalis cava a | 7.5% | 27.8% | 0.50 ± 0.27 | 2.63 ± 1.10 | 7 | 11.0 | 0.021 |
| Scilla bifolia a | 2.3% | 37.7% | 0.43 ± 0.30 | 3.86 ± 1.62 | 7 | 9.0 | 0.027 |
| Veronica hederifolia a | 5.6% | 10.0% | 10.57 ± 2.36 | 11.29 ± 3.24 | 7 | 23.5 | 0.898 |
| Viola riviniana a | 36.9% | 49.3% | 6.71 ± 6.26 | 12.71 ± 7.83 | 7 | 14.0 | 0.180 |
| Viola riviniana b | 1.0% | 12.5% | 0.29 ± 0.76 | 3.43 ± 2.82 | 7 | 5.5 | 0.009 |
| Plant species | Distance dispersal | Directed dispersal | Establishment success | Dispersal vectors |
|---|---|---|---|---|
| Corydalis cava | ++ | ○ | ○ | ants only |
| Scilla bifolia | ++ | + | ○ | ants only |
| Veronica hederifolia | + | ○ | ○ | ants and animals |
| Viola riviniana | ++ | ○ | ○ | autochory (ballistic dispersal) and ants |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fokuhl, G.; Heinze, J.; Poschlod, P. An Ant-Plant Mesocosm Experiment Reveals Dispersal Patterns of Myrmecochorous Plants. Forests 2019, 10, 1149. https://doi.org/10.3390/f10121149
Fokuhl G, Heinze J, Poschlod P. An Ant-Plant Mesocosm Experiment Reveals Dispersal Patterns of Myrmecochorous Plants. Forests. 2019; 10(12):1149. https://doi.org/10.3390/f10121149
Chicago/Turabian StyleFokuhl, Gerriet, Jürgen Heinze, and Peter Poschlod. 2019. "An Ant-Plant Mesocosm Experiment Reveals Dispersal Patterns of Myrmecochorous Plants" Forests 10, no. 12: 1149. https://doi.org/10.3390/f10121149
APA StyleFokuhl, G., Heinze, J., & Poschlod, P. (2019). An Ant-Plant Mesocosm Experiment Reveals Dispersal Patterns of Myrmecochorous Plants. Forests, 10(12), 1149. https://doi.org/10.3390/f10121149