Biomass Accumulation and Carbon Sequestration in an Age-Sequence of Mongolian Pine Plantations in Horqin Sandy Land, China


Abstract
:1. Introduction
2. Materials and Methods
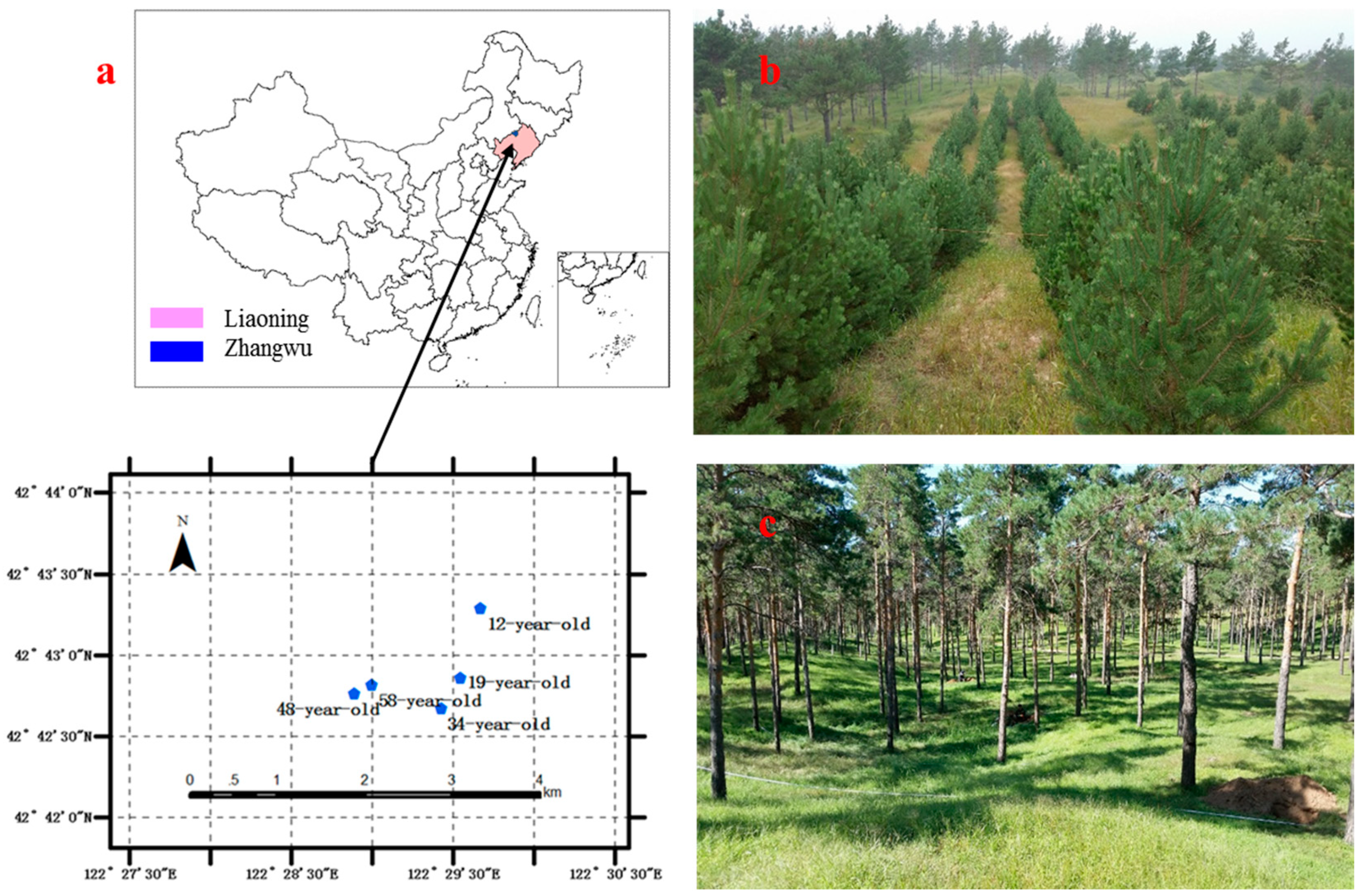
2.1. Site Description
2.2. Forest Inventory and Measurements
2.3. Soil Measurements and Sampling
2.4. Destructive Tree Sampling
2.5. Allometric Biomass Equations for Tree Components
2.6. Biomass and Carbon Density
2.7. Statistical Analysis
3. Results
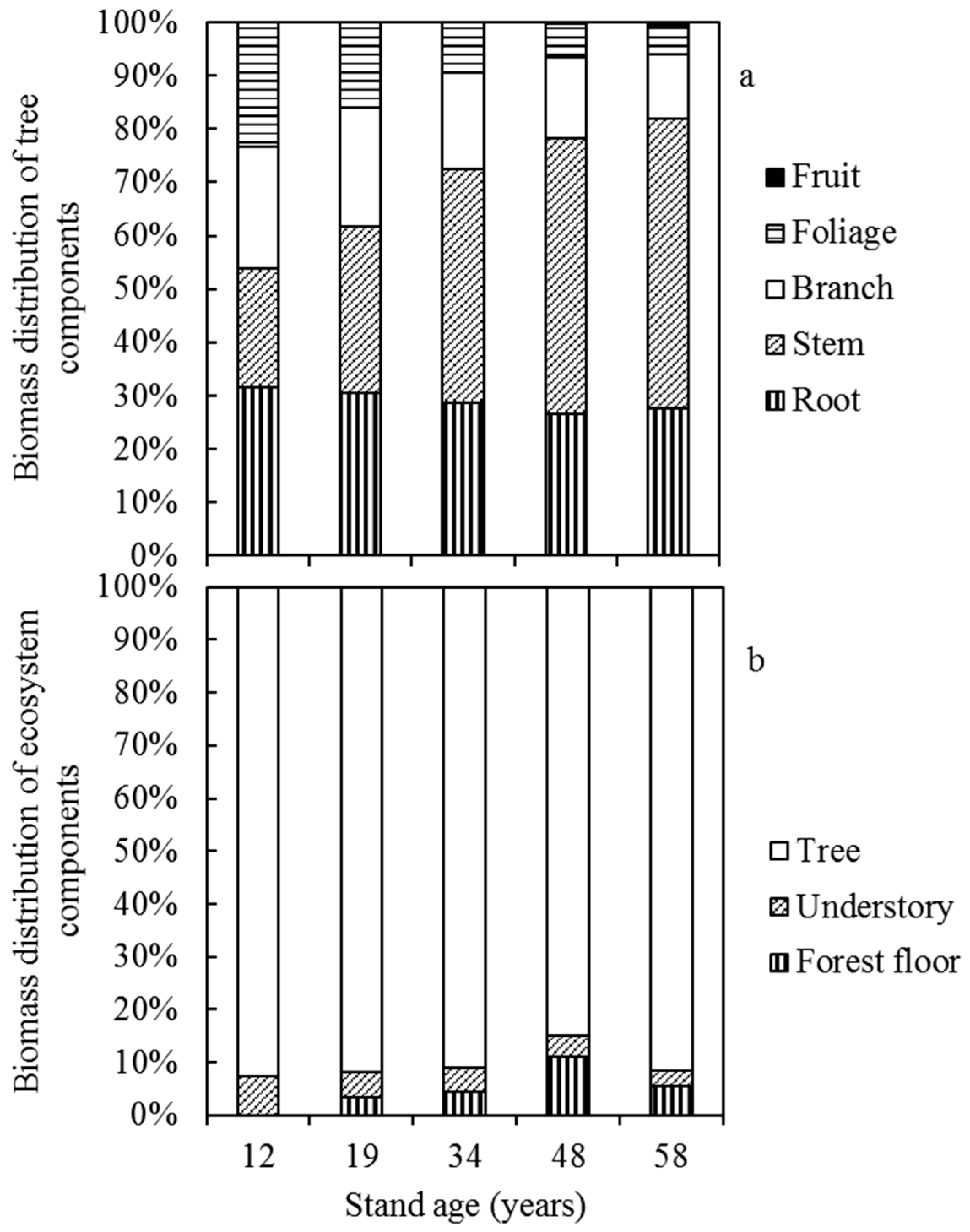
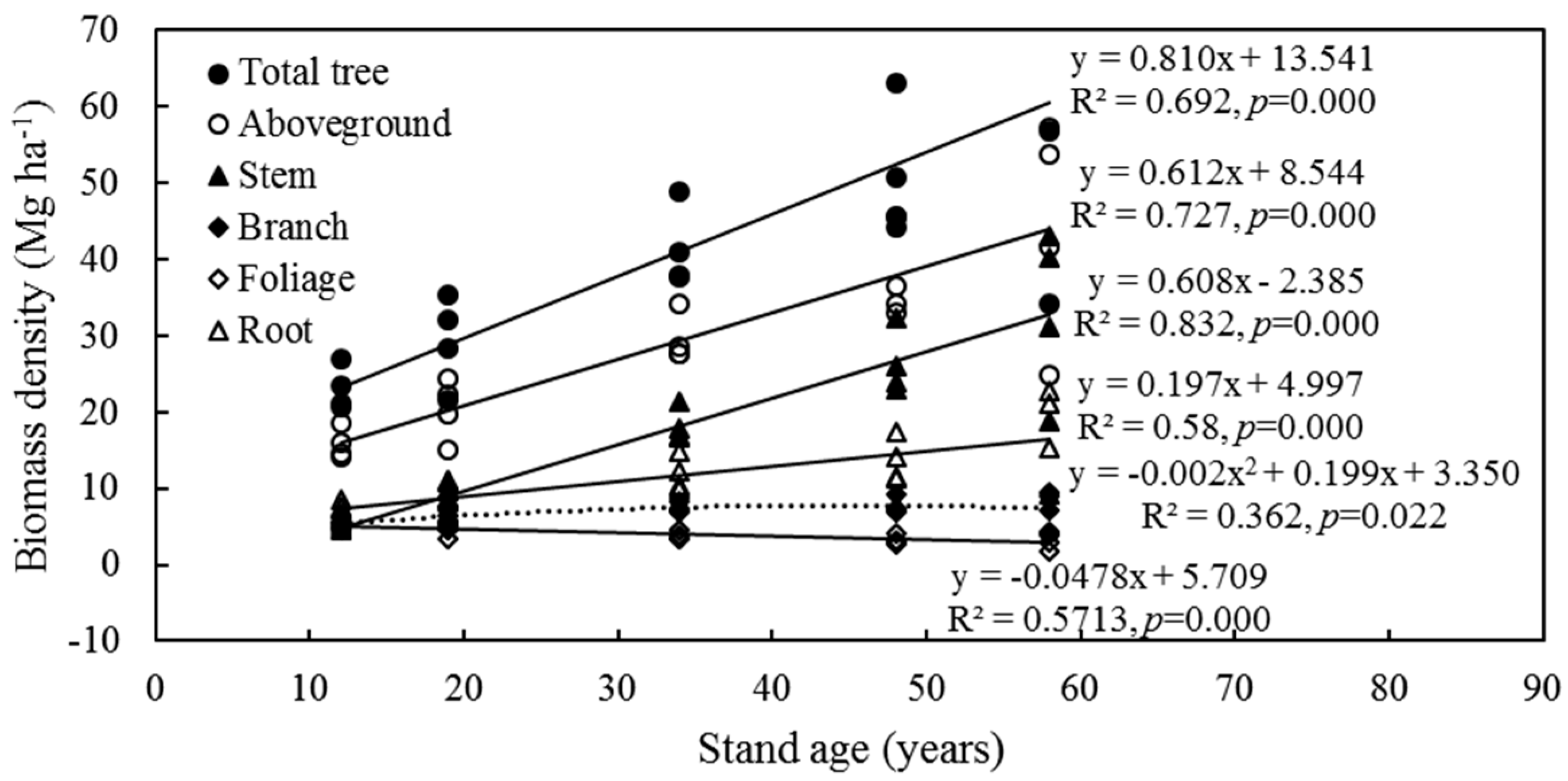
3.1. Tree Biomass and Its Distribution
3.2. Biomass of Understory and Forest Floor
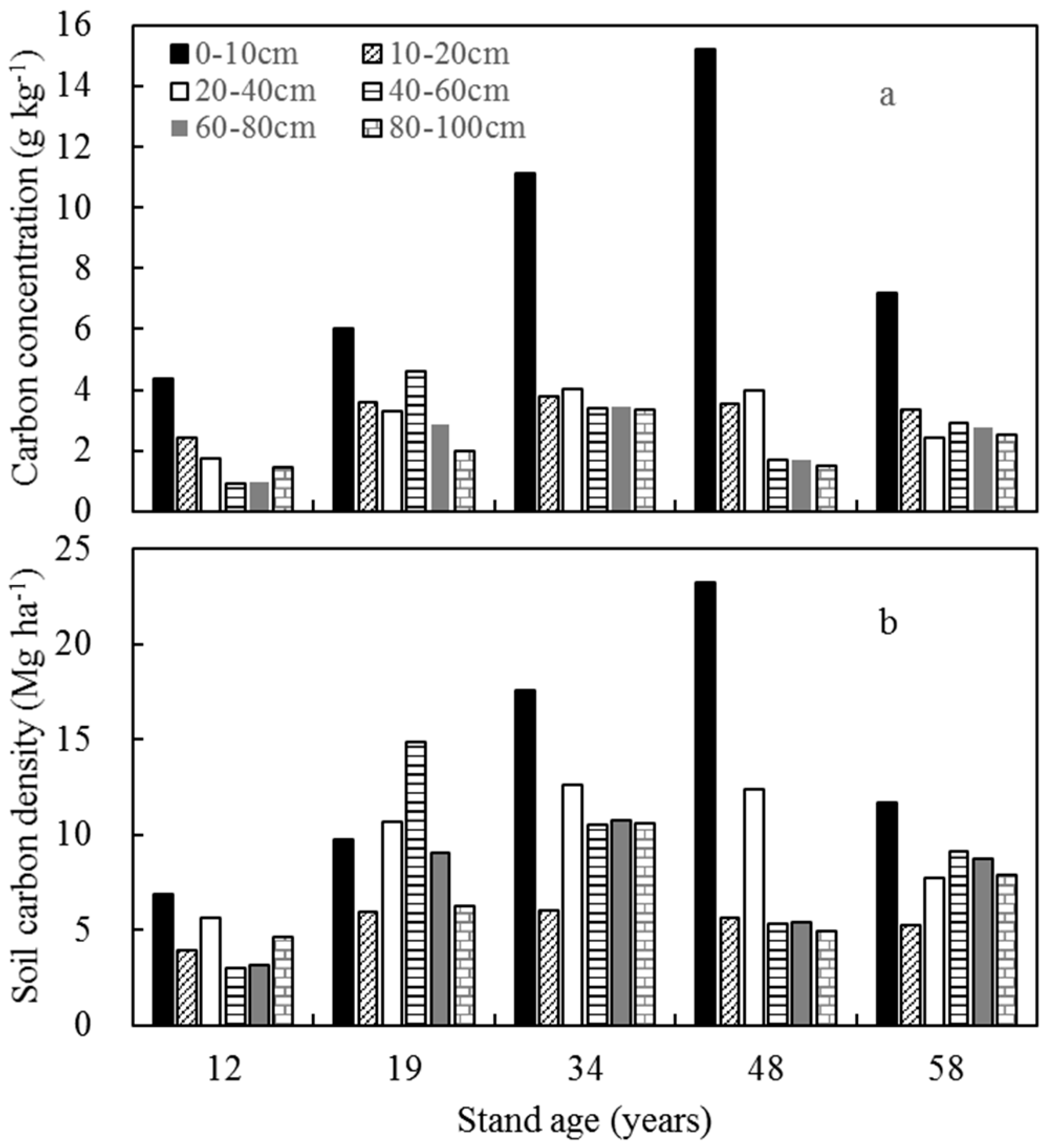
3.3. Carbon Concentration
3.4. Soil Carbon Density and Ecosystem Carbon Pools
4. Discussion
4.1. Allometric Biomass Equation for Tree Components
4.2. Biomass Distribution
4.3. Changes in Soil Carbon Density
4.4. Carbon Density and Distribution for Ecosystem Components
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Dixon, R.K.; Brown, S.; Houghton, R.A.; Solomon, A.M.; Trexler, M.C.; Wisniewski, J. Carbon pools and flux of global forest ecosystem. Science 1994, 263, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Schimel, D.S.; House, J.I.; Hibbard, K.A.; Bousquet, P.; Ciais, P.; Peylin, P.; Braswell, B.H.; Apps, M.J.; Baker, D.; Bondeau, A.; et al. Recent patterns and mechanisms of carbon exchange by terrestrial ecosystems. Nature 2001, 414, 169–172. [Google Scholar] [CrossRef] [PubMed]
- FAO. Global Forest Resources Assessment 2015. How are the World’S Forests Changing? 2nd ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016; pp. 1–54. [Google Scholar]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- State Forestry Administration of China (SFA). 2014 China Forestry Development Report; Chinese Forestry Publisher: Beijing, China, 2014. [Google Scholar]
- State Forestry Administration of China (SFA). Bulletion of the fifth Desertification and Sandification State of China. State Forestry Administration, China. December 2015. Available online: http://cfdb.forestry.gov.cn/lysjk/indexJump.do?url=view/moudle/searchData/showDetail&keyid=100702 (accessed on 16 January 2019).
- Xu, X.L.; Li, K.R. Biomass carbon sequestration by planted forests in China. Chin. Geogr. Sci. 2010, 20, 289–297. [Google Scholar] [CrossRef]
- Xu, B.; Guo, Z.; Piao, S.; Fang, J. Biomass carbon stocks in China’s forests between 2000 and 2050: A prediction based on forest biomass-age relationships. Sci. China Life Sci. 2010, 53, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, A.; Mäkipää, R.; Heikkinen, J.; Sievänen, R.; Liski, J. Biomass expansion factors (BEFs) for Scots pine, Norway spruce and birch according to stand age for boreal forests. For. Ecol. Manag. 2004, 188, 211–224. [Google Scholar] [CrossRef] [Green Version]
- Andrzej, M.J.; Marcin, K.D.; Kamil, G.; Pawel, H.; Agnieszka, C.; Sylwia, W.; Karol, M. How do tree stand parameters affect young Scots pine biomass?-Allometric equations and biomass conversion and expansion factors. For. Ecol. Manag. 2018, 409, 74–83. [Google Scholar]
- Jagodziński, A.M.; Zasada, M.; Bronisz, K.; Bronisz, A.; Bijak, S. Biomass conversion and expansion factors for a chronosequence of young naturally regenerated silver birch (Betula pendula Roth) stands growing on post-agricultural sites. For. Ecol. Manag. 2017, 384, 208–220. [Google Scholar] [CrossRef]
- Peichl, M.; Arain, M.A. Allometry and partitioning of above- and belowground tree biomass in an age-sequence of white pine forests. For. Ecol. Manag. 2007, 253, 68–80. [Google Scholar] [CrossRef]
- Cao, J.; Wang, X.; Tian, Y.; Wen, Z.; Zha, T. Pattern of carbon allocation across three different stages of stand development of a Chinese pine (Pinus tabulaeformis) forest. Ecol. Res. 2012, 27, 883–892. [Google Scholar] [CrossRef]
- Justine, M.F.; Yang, W.; Wu, F.; Khan, M.N. Dynamics of biomass and carbon sequestration across a chronosequence of masson pine plantations. J. Geophys. Res. Biogeosci. 2017, 122, 122. [Google Scholar] [CrossRef]
- Cheng, J.; Lee, X.; Theng, B.K.G.; Zhang, L.; Fang, B. Biomass accumulation and carbon sequestration in an age-sequence of Zanthoxylum bungeanum plantations under the Grain for Green Program in karst regions, Guizhou province. Agric. For. Meterol. 2015, 203, 88–95. [Google Scholar] [CrossRef]
- White, J.C.; Wulder, M.A.; Varhola, A.; Vastaranta, M.; Coops, N.C.; Cook, B.D.; Pitt, D.G.; Woods, M. A Best Practices Guide for Generating Forest Inventory Attributes from Airborne Laser Scanning Data Using an Area-Based Approach (Version 2.0); Natural Resources Canada, Canadian Forest Service, Canadian Wood Fibre Centre: Victoria, BC, Canada, 2013.
- Wang, F.; Xu, X.; Zou, B.; Guo, Z.; Li, Z.; Zhu, W. Biomass accumulation and carbon sequestration in four different aged Casuarina equisetifolia coastal shelterbelt plantations in South China. PLoS ONE 2013, 8, e77449. [Google Scholar] [CrossRef] [PubMed]
- Justine, M.F.; Yang, W.; Wu, F.; Tan, B.; Khan, M.N.; Zhao, Y. Biomass stock and carbon sequestration in a chronosequence of Pinus massoniana plantations in the upper reaches of the Yangtze river. Forests 2015, 6, 3665–3682. [Google Scholar] [CrossRef]
- Cheng, X.; Han, H.; Kang, F.; Song, Y.; Liu, K. Variation in biomass and carbon storage by stand age in pine (Pinus tabulaeformis) planted ecosystem in Mt. Taiyue. Shanxi, China. J. Plant Interact. 2014, 9, 521–528. [Google Scholar] [CrossRef]
- Chen, G.S.; Yang, Z.J.; Gao, R.; Xie, J.S.; Guo, J.F.; Huang, Z.Q.; Yang, Y.S. Carbon storage in a chrononsequence of Chinese fir plantations in southern China. For. Ecol. Manag. 2013, 300, 68–76. [Google Scholar] [CrossRef]
- Zhao, J.; Kang, F.; Wang, L.; Yu, X.; Zhao, W.; Song, X.; Zhang, Y.; Chen, F.; Sun, Y.; He, T.; et al. Patterns of biomass and carbon distribution across a chronosequence of Chinese Pine (Pinus tabulaeformis) forests. PLoS ONE 2014, 9, e94966. [Google Scholar] [CrossRef]
- Peichl, M.; Arain, M.A. Above- and belowground ecosystem biomass and carbon pools in an age-sequence of temperate pine plantation forests. Agric. For. Meteorol. 2006, 140, 51–63. [Google Scholar] [CrossRef]
- Paul, K.I.; Khanna, P.K.; Polglase, P.J.; Nyakuengama, J.G. Change in soil carbon following afforestation. For. Ecol. Manag. 2002, 168, 241–257. [Google Scholar] [CrossRef]
- Eggleston, S.; Buedia, L.; Miwa, K.; Ngara, T.; Tanabe, K. IPCC Guidelines for National Greenhouse Gas Inventories, Prepared by the National Greenhouse Gas Inventories Programme; IGES: Hayama, Japan, 2006. [Google Scholar]
- Li, X.; Yi, M.J.; Son, Y.; Park, P.S.; Lee, K.H.; Son, Y.M.; Kim, R.H.; Jeong, M.J. Biomass and carbon storage in an age-sequence of Korea pine (Pinus koraiensis) plantation forests in central Korea. J. Plant Biol. 2011, 54, 33–42. [Google Scholar] [CrossRef]
- Taylor, A.R.; Wang, J.R.; Chen, H.Y.H. Carbon storage in a chronosequence of red spruce (Picea rubens) forests in central Nova Scotia, Canada. Can. J. For. Res. 2007, 37, 2260–2269. [Google Scholar] [CrossRef]
- Yaun, L.; Yan, D.; Wang, Y.; Jiang, P.; Liu, Y.; Dang, X. Carbon storage of Pinus sylvestris var. mongolica Litv. Plantation in sandy land. J. Inner Mongolica For. Sci. Technol. 2011, 37, 9–13. [Google Scholar]
- Zhu, J.J.; Kang, H.Z.; Tan, H.; Xu, M.L. Effects of drought stresses induced by polyethylene glycol on germination of Pinus sylvestris var. mongolica seeds from natural and plantation forests on sandy land. J. For. Res. 2006, 11, 319–328. [Google Scholar] [CrossRef]
- Song, L.; Zhu, J.; Li, M.; Zhang, J.; Zheng, X.; Wang, K. Canopy transpiration of Pinus sylvestris var. mongolica in a sparse wood grassland in the semiarid sandy region of Northeast China. Agric. For. Meteorol. 2018, 250–251, 192–201. [Google Scholar] [CrossRef]
- Zhu, J.J.; Li, F.Q.; Xu, M.L.; Kang, H.Z.; Xu, D.Y. The role of ectomycorrhizal fungi in alleviating pine decline in semiarid sandy soil of northern China: An experimental approach. Ann. For. Sci. 2008, 65, 1–12. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, A.; An, Y.; Lian, P.; Wu, J.; Meinzer, F.C.; Hao, G. Hydraulics play an important role in causing low growth rate and dieback of aging Pinus sylvestris var. mongolica trees in plantations of Northeast China. Plant Cell Environ. 2018, 41, 1500–1511. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.R. Report on the causes of the early decline of Pinus sylvestris var. mongolica shelterbelt and its preventative and control measures in Zhanggutai of Liaoning province. Sci. Silva. Sin. 2001, 37, 131–138. [Google Scholar]
- Song, L.; Zhu, J.; Li, M.; Zhang, J.; Lv, L. Sources of water used by Pinus sylvestris var. mongolica trees based on stable isotope measurements in a semiarid sandy region of Northeast China. Agric. Water Manag. 2016, 164, 281–290. [Google Scholar] [CrossRef]
- Zheng, X.; Zhu, J.J.; Yan, Q.L.; Song, L.N. Effects of land use changes on the groundwater table and the decline of Pinus sylvestris var. mongolica plantations in southern Horqin Sandy Land, Northeast China. Agric. Water Manag. 2012, 109, 94–106. [Google Scholar] [CrossRef]
- Zhu, J.; Zeng, D.; Kang, H.; Wu, X.; Fan, Z. Decline of Pinus sylvestris var. mongolica Plantations on Sandy Land; Chinese Forestry Publisher: Beijing, China, 2005; pp. 1–242. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon and Organic Matter. In Methods of Soil Analysis, Part 2; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy, Inc.: Madison, WI, USA, 1982; pp. 539–579. [Google Scholar]
- Jiao, S. A preliminary study of the biomass and nutrient elements distribution in cultivated Mongolian pine forests in the Zhanggutai region, Liaoning province. Acta Phytotax. Geobot. 1985, 9, 257–265. [Google Scholar]
- Liu, H.; Lv, S.; Liu, Q.; Liu, L.; Wang, Y.; Zhang, H. Biomass and carbon storage of the Pinus sylvestris var. mongolica Litv. plantation in the Duolun country. J. Inner Mongolia Agric. Univ. 2013, 34, 49–53. [Google Scholar]
- Xing, J.; Zheng, C.; Feng, C.; Zeng, F. Change of growth characters and carbon stocks in plantations of Pinus sylvestris var. mongolica in Saihanba, Hebei, China. Chin. J. Plan. Ecol. 2017, 41, 840–849. [Google Scholar]
- Samuelson, L.J.; Stokes, T.A.; Butnor, J.R.; Johnsen, K.H.; Gonzalea-Benecke, C.A.; Anderson, P.; Jackson, J.; Ferrari, L.; Martin, T.A.; Cropper, W.P., Jr. Ecosystem carbon stocks in Pinus palustris forests. Can. J. For. Res. 2014, 44, 476–486. [Google Scholar] [CrossRef]
- Lee, J.; Tolunay, D.; Makineci, E.; Cömez, A.; Son, Y.M.; Kim, R.; Son, Y. Estimating the age-dependent changes in carbon stocks of Scots pine (Pinus sylvestris L.) stands in Turkey. Ann. For. Sci. 2016, 73, 523–531. [Google Scholar] [CrossRef]
- Ryan, M.G.; Binkley, D.; Fownes, J.H. Age-related decline in forest productivity: Pattern and process. Adv. Ecol. Res. 1997, 27, 213–262. [Google Scholar]
- Fang, J.; Chen, A.; Peng, C.; Zhao, S.; Ci, L. Changes in forest biomass carbon storage in China between 1949 and 1998. Science 2001, 292, 2320–2322. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Wei, Y.; Qin, S.; Deng, L.; Zhou, Y. Carbon storage dynamics and its distribution pattern in Pinus sylvestris var. mongolica plantation in sandy land. Chin. J. Ecol. 2015, 34, 1798–1803. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Age (Years) | Location | Altitude (m) | Number of Trees | Mean Height (m) | Range (m) | Mean D (cm) | Range (cm) | Soil Bulk Density (g cm−3) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0–10 cm | 10–20 cm | 20–40 cm | 40–60 cm | 60–80 cm | 80–100 cm | ||||||||
| 12 | 42.7215° N, 122.4946° E | 231.4 | 324 | 2.22 | 0.70–3.50 | 5.9 | 4.33–7.09 | 1.56 ± 0.02 | 1.60 ± 0.04 | 1.59 ± 0.03 | 1.60 ± 0.02 | 1.62 ± 0.02 | 1.61 ± 0.02 |
| 19 | 42.7143° N, 122.4925° E | 230.4 | 168 | 4.15 | 1.50–7.30 | 9.95 | 4.30–14.20 | 1.61 ± 0.02 | 1.64 ± 0.04 | 1.63 ± 0.03 | 1.61 ± 0.01 | 1.58 ± 0.02 | 1.56 ± 0.04 |
| 34 | 42.7112° N, 122.4905° E | 235.4 | 91 | 8.06 | 6.00–10.50 | 17.3 | 10.30–26.00 | 1.58 ± 0.12 | 1.59 ± 0.02 | 1.56 ± 0.02 | 1.56 ± 0.01 | 1.56 ± 0.01 | 1.59 ± 0.02 |
| 48 | 42.7127° N, 122.4816° E | 247.5 | 61 | 11.08 | 8.00–12.70 | 23.68 | 13.00–31.50 | 1.53 ± 0.04 | 1.59 ± 0.01 | 1.55 ± 0.00 | 1.58 ± 0.02 | 1.61 ± 0.07 | 1.63 ± 0.07 |
| 58 | 42.7136° N, 122.4834° E | 225.5 | 77 | 12.2 | 10.50–15.60 | 23.56 | 15.30–35.30 | 1.62 ± 0.05 | 1.57 ± 0.06 | 1.58 ± 0.01 | 1.57 ± 0.05 | 1.57 ± 0.03 | 1.56 ± 0.02 |
| Biomass of Tree Components (kg) | Parameters in Equation (1) | Parameters in Equation (2) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| a | b | R2 | Significance | a | b | c | R2 | Significance | |
| Total tree | 0.630 | 1.852 | 0.975 | 0.000 ** | 1.515 | 2.644 | −0.902 | 0.987 | 0.000 ** |
| Aboveground | 0.373 | 1.922 | 0.974 | 0.000 ** | 0.954 | 2.769 | −0.965 | 0.986 | 0.000 ** |
| Tree stem | 0.052 | 2.415 | 0.992 | 0.000 ** | 0.065 | 2.608 | −0.220 | 0.993 | 0.000 ** |
| Stem wood | 0.035 | 2.515 | 0.991 | 0.000 ** | 0.042 | 2.683 | −0.192 | 0.991 | 0.000 ** |
| Stem bark | 0.033 | 1.830 | 0.993 | 0.000 ** | 0.035 | 1.892 | −0.071 | 0.993 | 0.000 ** |
| Branch | 0.192 | 1.671 | 0.913 | 0.000 ** | 0.781 | 2.939 | −1.444 | 0.948 | 0.001 ** |
| Foliage | 0.610 | 1.036 | 0.835 | 0.002 ** | 2.265 | 2.220 | −1.348 | 0.907 | 0.003 ** |
| Fruit | 3.616 × 10−7 | 4.187 | 0.595 | 0.045 * | 4.836 × 10−12 | 0.139 | 6.412 | 0.920 | 0.011 * |
| Root | 0.286 | 1.676 | 0.977 | 0.000 ** | 0.585 | 2.323 | −0.737 | 0.987 | 0.000 ** |
| Ecosystem Components | Stand Age (Years) | ||||
|---|---|---|---|---|---|
| 12 | 19 | 34 | 48 | 58 | |
| Total Tree | 23.10 ± 2.83 B | 29.32 ± 5.95 AB | 41.37 ± 5.28 A | 50.88 ± 8.56 A | 61.49 ± 20.66 AB |
| Tree aboveground | 15.77 ± 1.99 C | 20.39 ± 4.06 BC | 29.51 ± 3.18 AB | 37.35 ± 5.78 A | 44.44 ± 14.60 ABC |
| Tree stem | 5.12 ± 0.64 B | 9.13 ± 1.83 B | 18.18 ± 2.10 A | 26.32 ± 4.19 A | 33.35 ± 11.00 AB |
| Tree stem wood | 4.01 ± 0.51 B | 7.66 ± 1.53 B | 16.05 ± 1.75 A | 23.77 ± 3.69 A | 30.14 ± 9.88 AB |
| Tree stem bark | 1.11 ± 0.13 A | 1.47 ± 0.31 A | 2.12 ± 0.36 A | 2.54 ± 0.50 A | 3.21 ± 1.12 A |
| Tree branch | 5.28 ± 0.70 B | 6.54 ± 1.27 AB | 7.48 ± 0.61 A | 7.66 ± 1.02 A | 7.43 ± 2.34 A |
| Tree foliage | 5.37 ± 0.66 A | 4.71 ± 0.96 AB | 3.84 ± 0.51 BC | 3.26 ± 0.56 C | 3.19 ± 1.08 C |
| Tree fruit | 0.00 ± 0.00 B | 0.00 ± 0.00 A | 0.02 ± 0.01 AB | 0.11 ± 0.04 AB | 0.47 ± 0.19 AB |
| Tree root | 7.32 ± 0.84 A | 8.93 ± 1.90 A | 11.86 ± 2.17 A | 13.53 ± 2.82 A | 17.05 ± 6.07 A |
| Total Understory | 1.87 ± 0.74 A | 1.47 ± 0.32 A | 2.02 ± 0.62 A | 2.45 ± 1.83 A | 1.99 ± 0.50 A |
| Understory aboveground | 1.32 ± 0.53 A | 0.90 ± 0.20 A | 1.24 ± 0.38 A | 1.50 ± 1.12 A | 1.22 ± 0.30 A |
| Understory belowground | 0.54 ± 0.22 A | 0.57 ± 0.12 A | 0.78 ± 0.24 A | 0.95 ± 0.71 A | 0.77 ± 0.19 A |
| Forest Floor | 0.00 ± 0.00 B | 1.12 ± 0.97 AB | 2.04 ± 0.64 AB | 6.69 ± 4.49 AB | 3.65 ± 0.18 A |
| Total Ecosystem | 24.97 | 31.91 | 45.43 | 60.02 | 67.13 |
| Ecosystem Components | Stand Age (Years) | Average | ||||
|---|---|---|---|---|---|---|
| 12 | 19 | 34 | 48 | 58 | ||
| Tree stem wood | 55.96 | 56.19 | 55.42 | 57.07 | 56.49 | 56.23 ± 0.62 A |
| Tree stem bark | 52.31 | 53.04 | 49.05 | 53.28 | 49.96 | 51.53 ± 1.91 B |
| Tree branch | 52.55 | 52.96 | 56.84 | 56.01 | 54.13 | 54.50 ± 1.88 A |
| Tree foliage | 53.20 | 51.44 | 53.74 | 55.45 | 56.05 | 53.98 ± 1.84 AB |
| Tree fruit | 47.81 | 47.21 | 49.22 | 48.08 ± 1.03 C | ||
| Tree root | 52.65 | 54.77 | 57.64 | 56.04 | 57.27 | 55.68 ± 2.03 A |
| Understory aboveground | 44.22 | 46.37 | 44.02 | 44.87 ± 1.30 C | ||
| Understory belowground | 36.42 | 38.31 | 41.57 | 38.77 ± 2.61 D | ||
| Forest floor | 41.31 | 45.79 | 48.15 | 45.08 ± 3.48 C | ||
| Ecosystem Components | Stand Age (years) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 12 | 19 | 34 | 48 | 58 | ||||||
| Carbon Density | Percentage | Carbon Density | Percentage | Carbon Density | Percentage | Carbon Density | Percentage | Carbon Density | Percentage | |
| Total Tree | 12.31 | 30.63 | 15.87 | 21.61 | 23.10 | 24.85 | 28.66 | 31.88 | 34.43 | 39.47 |
| Tree aboveground | 8.46 | 21.04 | 10.98 | 14.95 | 16.26 | 17.50 | 21.07 | 23.44 | 24.67 | 28.27 |
| Tree stem | 2.82 | 7.03 | 5.08 | 6.92 | 9.94 | 10.69 | 14.92 | 16.60 | 18.63 | 21.35 |
| Tree stem wood | 2.24 | 5.58 | 4.30 | 5.86 | 8.90 | 9.57 | 13.57 | 15.09 | 17.02 | 19.51 |
| Tree stem bark | 0.58 | 1.45 | 0.78 | 1.06 | 1.04 | 1.12 | 1.36 | 1.51 | 1.60 | 1.84 |
| Tree branch | 2.77 | 6.90 | 3.47 | 4.72 | 4.25 | 4.58 | 4.29 | 4.77 | 4.02 | 4.61 |
| Tree foliage | 2.86 | 7.11 | 2.42 | 3.30 | 2.06 | 2.22 | 1.81 | 2.01 | 1.79 | 2.05 |
| Tree fruit | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 0.01 | 0.05 | 0.06 | 0.23 | 0.26 |
| Tree root | 3.86 | 9.59 | 4.89 | 6.66 | 6.83 | 7.35 | 7.58 | 8.44 | 9.77 | 11.19 |
| Total Understory | 0.78 | 1.95 | 0.64 | 0.87 | 0.86 | 0.92 | 1.05 | 1.17 | 0.85 | 0.97 |
| Understory aboveground | 0.59 | 1.46 | 0.42 | 0.57 | 0.56 | 0.60 | 0.66 | 0.73 | 0.55 | 0.63 |
| Understory belowground | 0.20 | 0.49 | 0.22 | 0.30 | 0.30 | 0.33 | 0.39 | 0.44 | 0.30 | 0.34 |
| Forest Floor | 0.00 | 0.00 | 0.51 | 0.70 | 0.92 | 0.99 | 3.22 | 3.58 | 1.65 | 1.89 |
| Soil Carbon Stock | 27.10 | 67.42 | 56.41 | 76.82 | 68.06 | 73.23 | 56.97 | 63.37 | 50.32 | 57.68 |
| Ecosystem Carbon Stock | 40.20 | 100.00 | 73.42 | 100.00 | 92.93 | 100.00 | 89.90 | 100.00 | 87.25 | 100.00 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhang, X.; Han, H.; Shi, Z.; Yang, X. Biomass Accumulation and Carbon Sequestration in an Age-Sequence of Mongolian Pine Plantations in Horqin Sandy Land, China. Forests 2019, 10, 197. https://doi.org/10.3390/f10020197
Zhang X, Zhang X, Han H, Shi Z, Yang X. Biomass Accumulation and Carbon Sequestration in an Age-Sequence of Mongolian Pine Plantations in Horqin Sandy Land, China. Forests. 2019; 10(2):197. https://doi.org/10.3390/f10020197
Chicago/Turabian StyleZhang, Xiao, Xueli Zhang, Hui Han, Zhongjie Shi, and Xiaohui Yang. 2019. "Biomass Accumulation and Carbon Sequestration in an Age-Sequence of Mongolian Pine Plantations in Horqin Sandy Land, China" Forests 10, no. 2: 197. https://doi.org/10.3390/f10020197
APA StyleZhang, X., Zhang, X., Han, H., Shi, Z., & Yang, X. (2019). Biomass Accumulation and Carbon Sequestration in an Age-Sequence of Mongolian Pine Plantations in Horqin Sandy Land, China. Forests, 10(2), 197. https://doi.org/10.3390/f10020197