A Useful Technical Application of the Identification of Nucleotide Sequence Polymorphisms and Gene Resources for Cinnamomum osmophloeum Kaneh. (Lauraceae)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Samples and Their Treatment
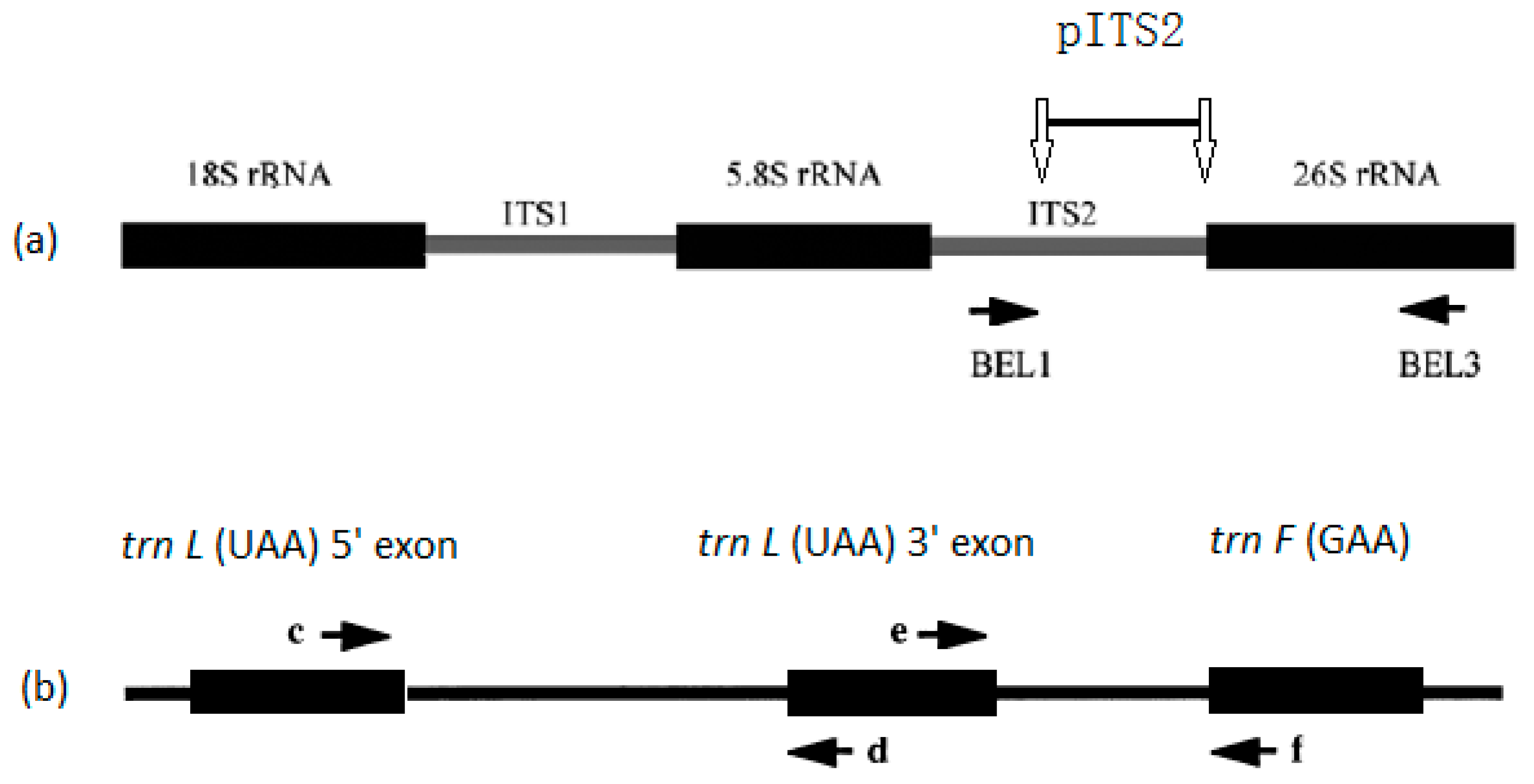
2.2. DNA Extraction, Polymerase Chain Reaction (PCR), and DNA Sequencing
2.3. Local DNA Database Establishment and Sequence Analysis
3. Results
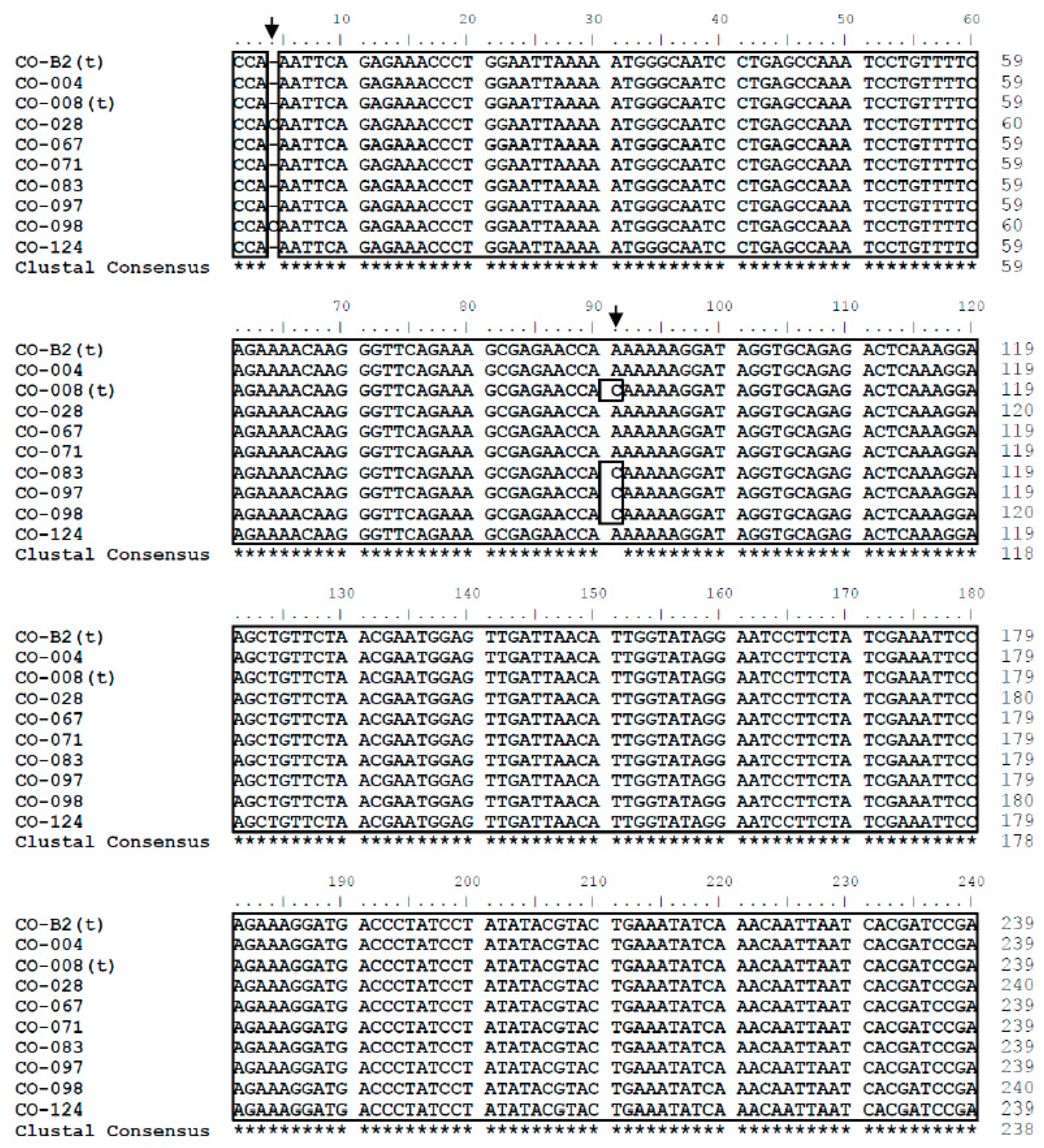
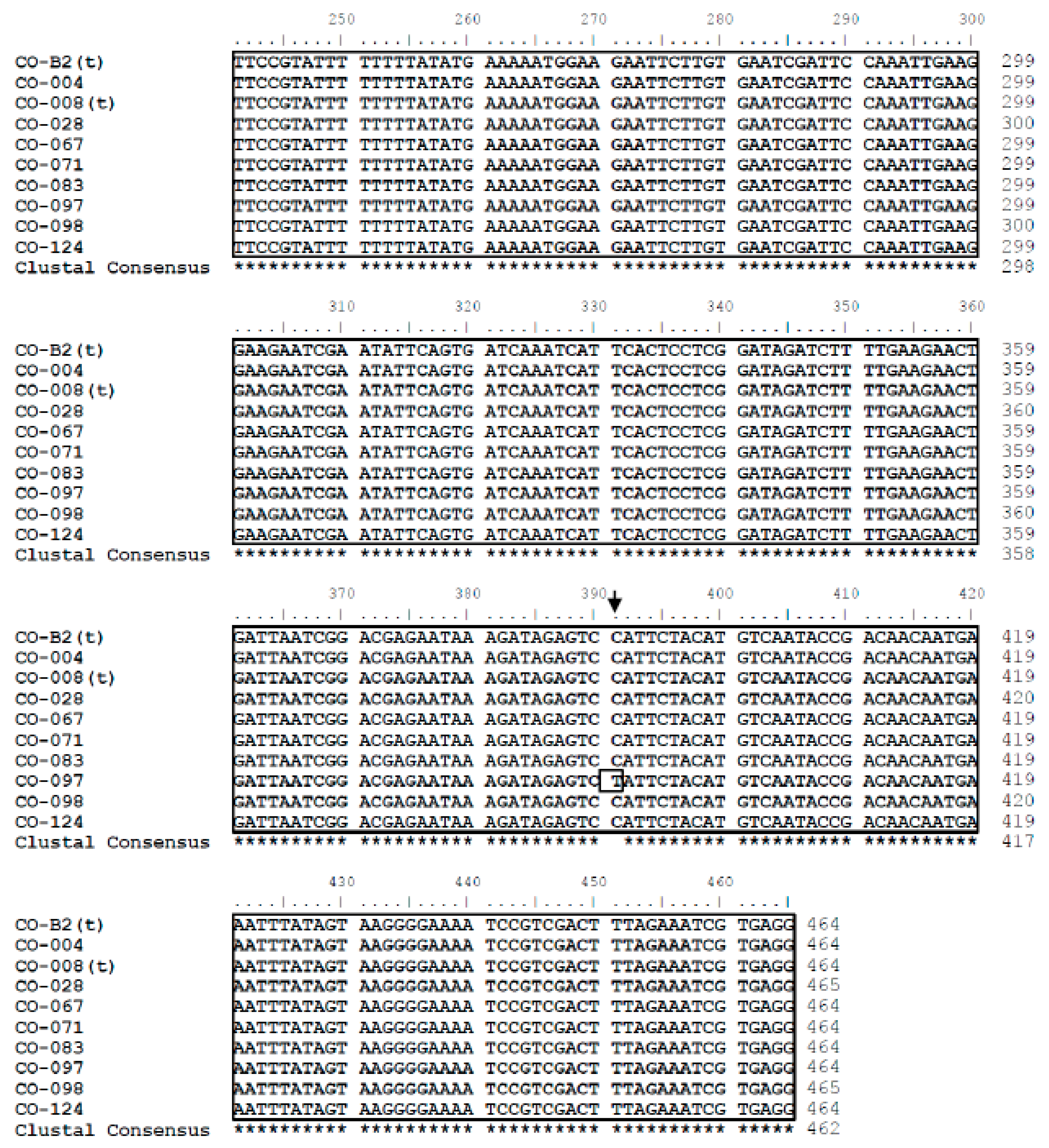
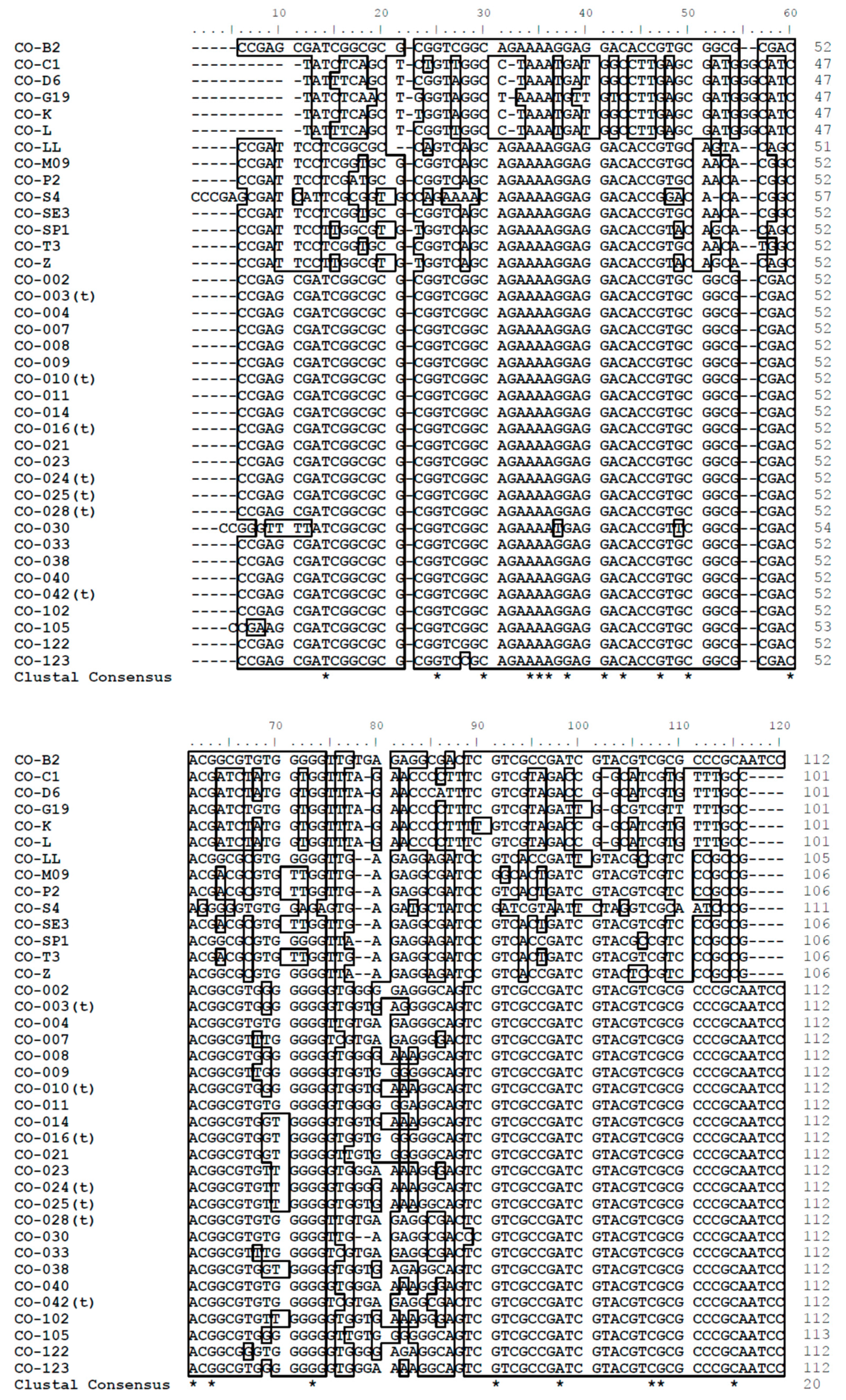
3.1. Sequence Analysis
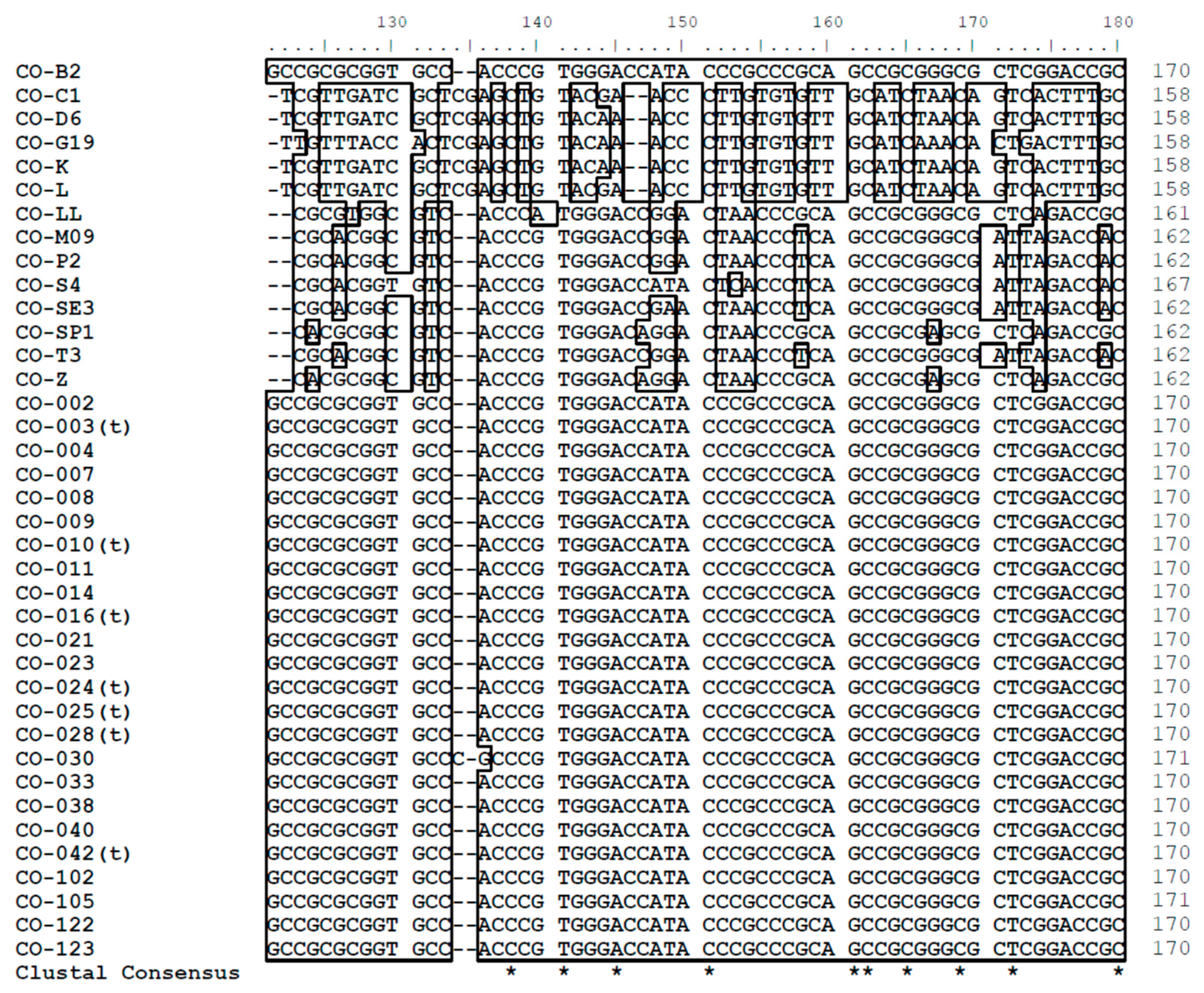
3.2. Nucleotide Sequence Polymorphism of C. osmophloem Kaneh.
3.3. Sequence Analysis of the DNA Database in This Study
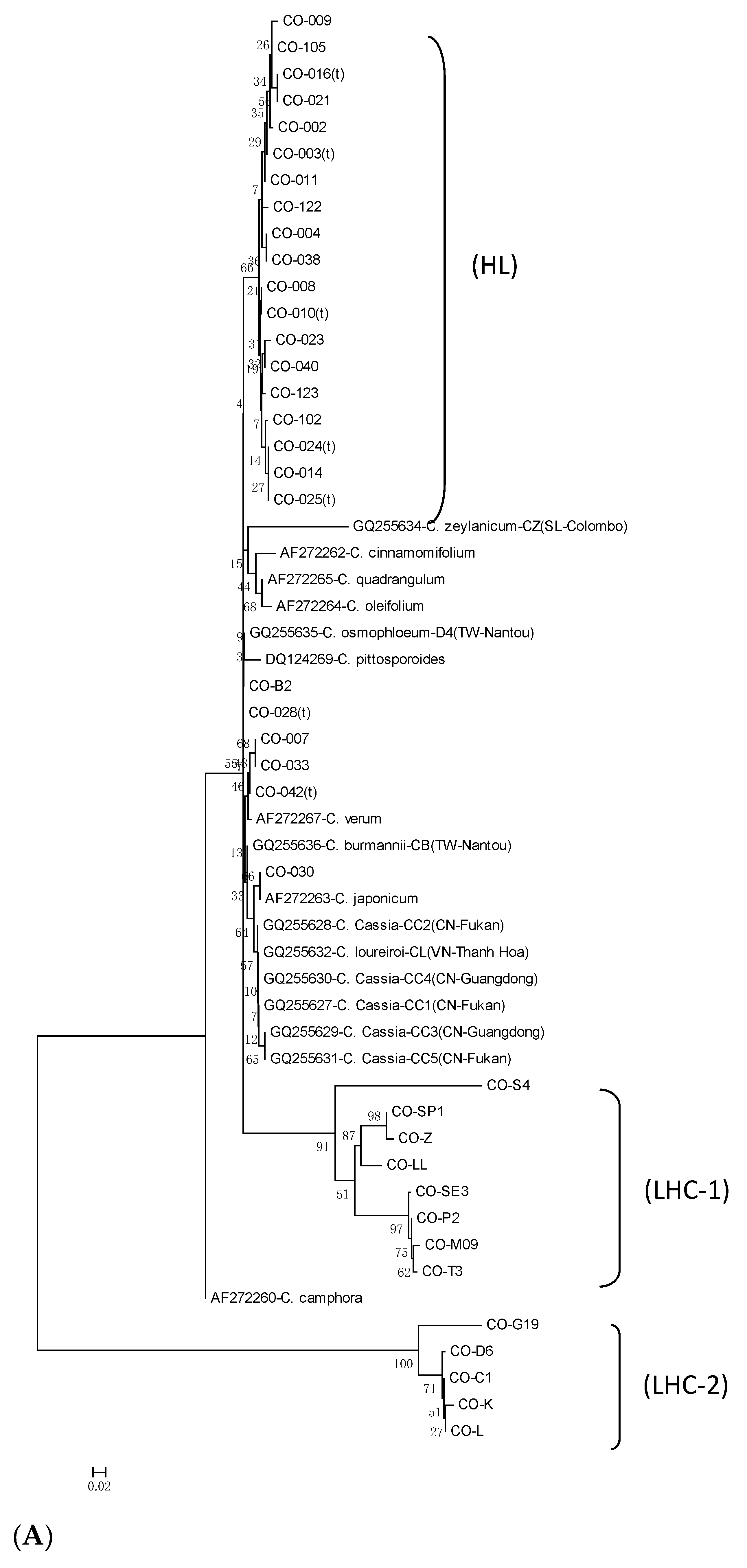
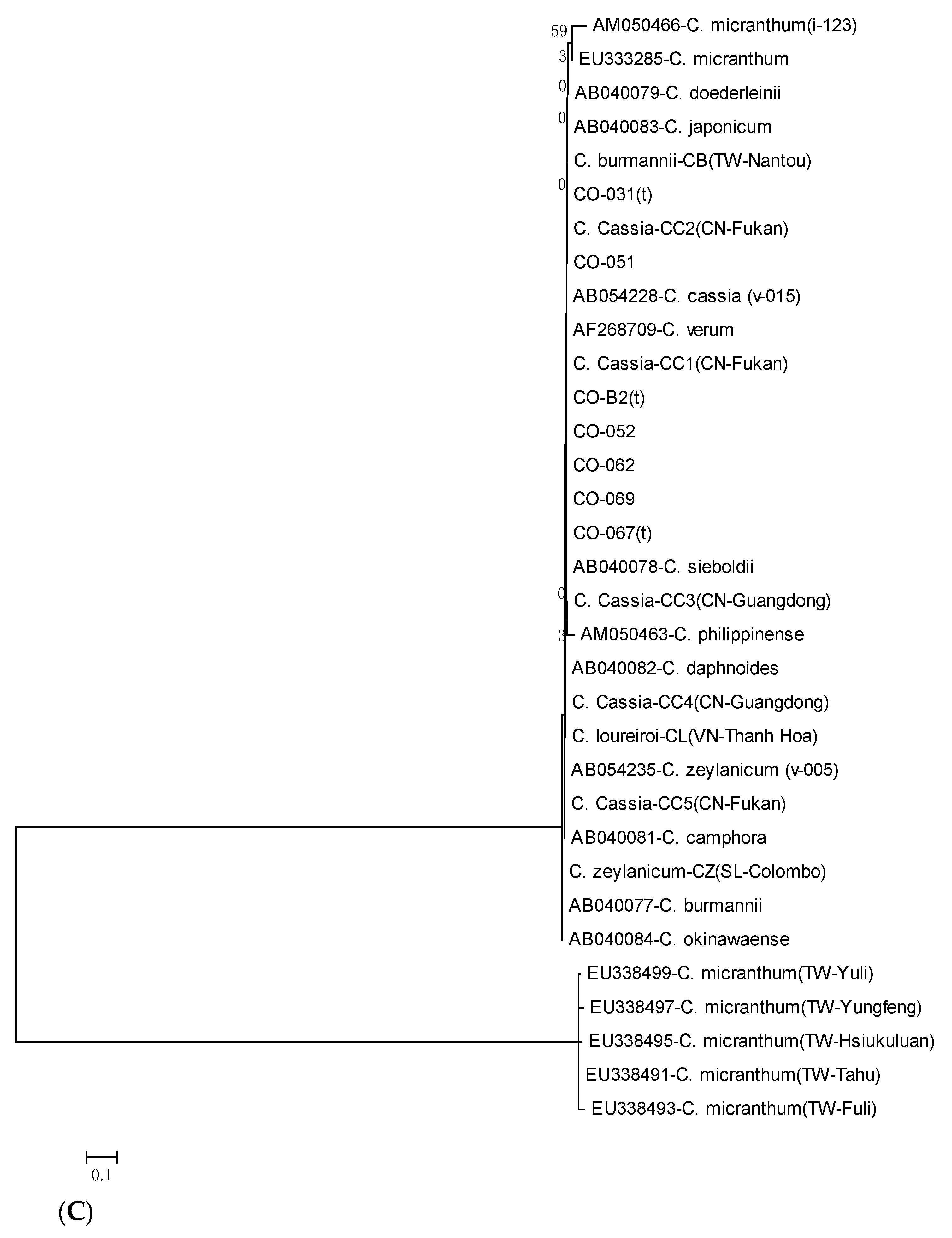
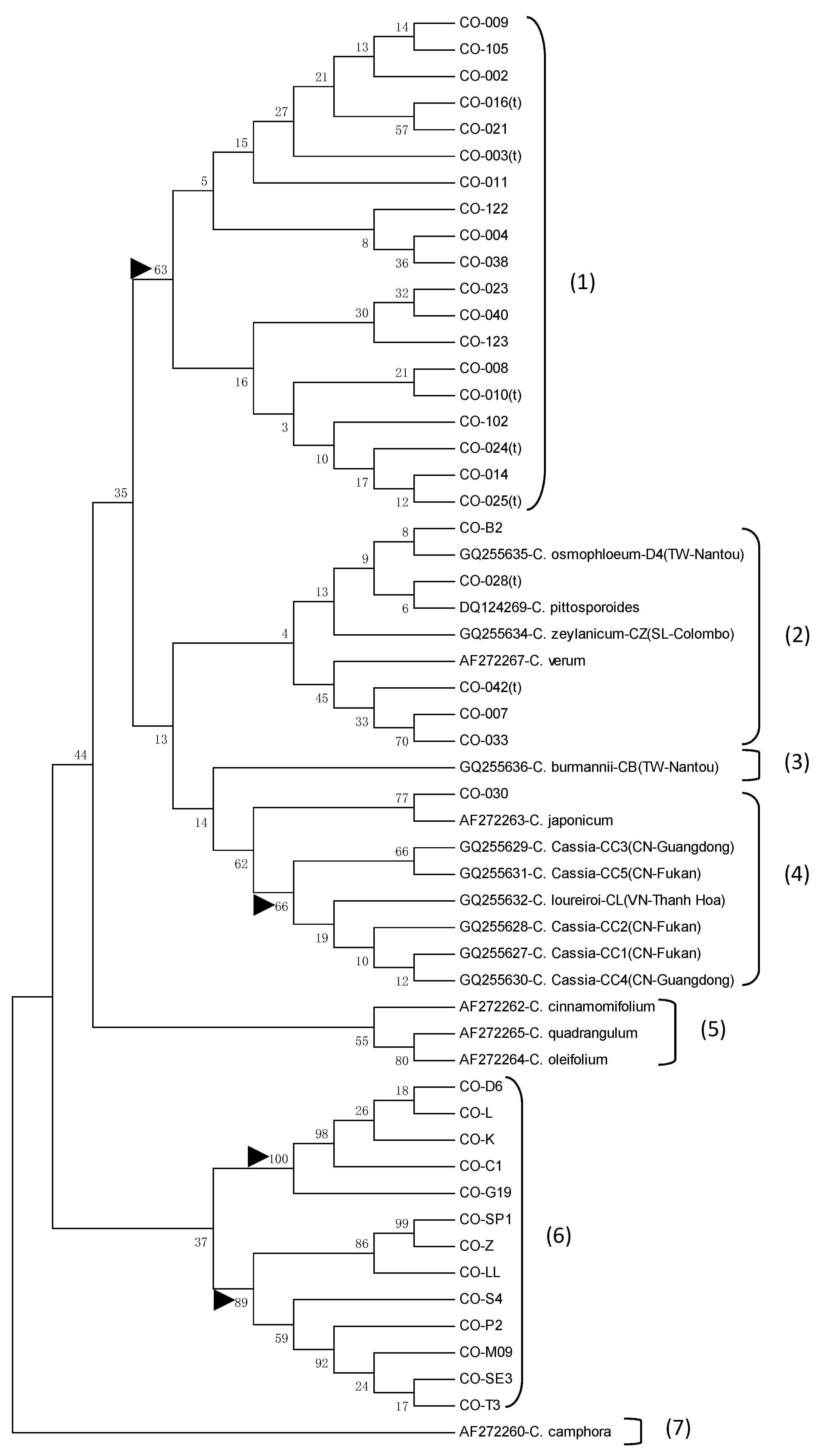
3.4. Phylogenetic Analysis and Gene Resource Identification
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Purseglove, J.W. Lauraceae. In Tropical Crops: Dicotyledons 2, 2nd ed.; Purseglove, J.W., Ed.; Longmans Green and Co. ltd.: London, UK, 1969; pp. 187–192. [Google Scholar]
- Liu, Y.J.; Lu, F.Y.; Ou, C.H. Trees of Taiwan; Monographic Publication No. 7; College of Agriculture, National Chung-Hshing University: Taichung, Taiwan, 1988. (In Chinese) [Google Scholar]
- Boufford, D.E.; Hsieh, C.F.; Huang, T.C.; Kuoh, C.S.; Ohashi, H.; Peng, C.I.; Tsai, J.L.; Yang, K.C. Flora of Taiwan; National Science Council: Taipei, Taiwan, 2003; Volumn 2, pp. 445–446, (Article in Chinese). [Google Scholar]
- Cheng, S.S.; Liu, J.Y.; Tsai, K.H.; Chen, W.J.; Chang, S.T. Chemical composition and mosquito larvicidal activity of essential oils from leaves of different Cinnamomum osmophloeum clones. J. Agric. Food Chem. 2004, 52, 4395–4400. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.S.; Liu, J.Y.; Lin, C.Y.; Hsui, Y.R.; Lu, M.C.; Wu, W.J.; Chang, S.T. Terminating red imported fire ants using Cinnamomum osmophloeum leaf essential oil. Bioresour. Technol. 2008, 99, 889–893. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.S.; Liu, J.Y.; Huang, C.G.; Hsui, Y.R.; Chen, W.J.; Chang, S.T. Insecticidal activities of leaf essential oils from Cinnamomum osmophloeum against three mosquito species. Bioresour. Technol. 2009, 100, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.T.; Chen, P.F.; Chang, S.C. Antibacterial activity of leaf essential oils and their constituents from Cinnamomum osmophloeum. J. Ethnopharmacol. 2001, 77, 123–127. [Google Scholar] [CrossRef]
- Chang, C.W.; Chang, W.L.; Chang, S.T.; Cheng, S.S. Antibacterial activities of plant essential oils against Legionella pneumophila. Water Res. 2008, 42, 278–286. [Google Scholar] [CrossRef]
- Wang, S.Y.; Chen, P.F.; Chang, S.T. Antifungal activities of essential oils and their constituents from indigenous cinnamon (Cinnamomum osmophloeum) leaves against wood decay fungi. Bioresour. Technol. 2005, 96, 813–818. [Google Scholar] [CrossRef]
- Cheng, S.S.; Liu, J.Y.; Hsui, Y.R.; Chang, S.T. Chemical polymorphism and antifungal activity of essential oils from leaves of different clones of indigenous cinnamon (Cinnamomum osmophloeum). Bioresour. Technol. 2006, 97, 306–312. [Google Scholar] [CrossRef]
- Shen, Y.C.; Chou, C.J.; Wang, Y.H.; Chen, C.F.; Chou, Y.C.; Lu, M.K. Anti-inflammatory activity of the extracts from mycelia of Antrodia camphorata cultured with water-soluble fractions from five different Cinnamomum species. FEMS Microbiol. Lett. 2004, 231, 137–143. [Google Scholar] [CrossRef]
- Chao, L.K.; Hua, K.F.; Hsu, H.Y.; Cheng, S.S.; Liu, J.Y.; Chang, S.T. Study on the anti-inflammatory activity of essential oil from leaves of Cinnamomum osmophloeum. J. Agric. Food Chem. 2005, 53, 7274–7278. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.H.; Rao, Y.K.; Tzeng, Y.M. Inhibitory effects of flavonol glycosides from Cinnamomum osmophloeum on inflammatory mediators in LPS/IFN-gamma-activated murine macrophages. Bioorg. Med. Chem. 2005, 13, 2381–2388. [Google Scholar] [CrossRef]
- Rao, Y.K.; Fang, S.H.; Tzeng, Y.M. Evaluation of the anti-inflammatory and anti-proliferation tumoral cells activities of Antrodia camphorata, Cordyceps sinensis, and Cinnamomum osmophloeum bark extracts. J. Ethnopharmacol. 2007, 114, 78–85. [Google Scholar] [CrossRef]
- Chao, L.K.; Hua, K.F.; Hsu, H.Y.; Cheng, S.S.; Lin, I.F.; Chen, C.J.; Chen, S.T.; Chang, S.T. Cinnamaldehyde inhibits pro-inf lammatory cytokines secretion from monocytes/macrophages through suppression of intracellular signaling. Food Chem. Toxicol. 2008, 46, 220–231. [Google Scholar] [CrossRef]
- Tung, Y.T.; Yen, P.L.; Lin, C.Y.; Chang, S.T. Anti-inflammatory activities of essential oils and their constituents from different provenances of indigenous cinnamon (Cinnamomum osmophloeum) leaves. Pharm. Biol. 2010, 48, 1130–1136. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Yang, C.W.; Liao, J.W.; Zhen, W.W.; Chu, F.H.; Chang, S.T. Essential oil from leaves of Cinnamomum osmophloeum acts as a xanthine oxidase inhibitor and reduces the serum uric acid levels in oxonate-induced mice. Phytomedicine 2008, 15, 940–945. [Google Scholar] [CrossRef]
- Chao, L.K.; Chang, W.T.; Shih, Y.W.; Huang, J.S. Cinnamaldehyde impairs high glucose-induced hypertrophy in renal interstitial fibroblasts. Toxicol. Appl. Pharmacol. 2010, 244, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.W. Distribution, Collection and the Germplasm Preservation of Cinnamomum osmophloeum Kaneh. In Monograph of Cinnamomum osmophloeum Kaneh.; Lin, T.P., Ed.; Taiwan Forestry Research Institute: Taipei, Taiwan, 1992; pp. 1–6. (In Chinese) [Google Scholar]
- Lee, H.C.; Chen, S.S.; Liu, J.Y.; Chang, S.T. Chemical polymorphism of leaf essential oils from different geographical clones of indigenous cinnamon (Cinnamomum osmophloeum). Q. J. Chin. Fore. 2003, 36, 411–422. [Google Scholar]
- Cheng, S.S.; Lin, C.Y.; Wang, Y.N.; Hsui, Y.R.; Chang, S.T. Set-up of the Source Bank of Cinnamomum osmophloeum—Chemical Polymorphism and Composition of Leaf Essential Oils. J. Exp. For. Nat. Taiwan Univ. 2008, 22, 21–34. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Alvarez, I.; Wendel, J.F. Ribosomal ITS sequences and plant phylogenetic inference. Mol. Phylogenet. Evol. 2003, 29, 417–434. [Google Scholar] [CrossRef]
- Lau, D.T.; Shaw, P.C.; Wang, J.; But, P.P. Authentication of medicinal Dendrobium species by the internal transcribed spacer of ribosomal DNA. Planta Med. 2001, 67, 456–460. [Google Scholar]
- Chiou, S.J.; Yen, J.H.; Fang, C.L.; Chen, H.L.; Lin, T.Y. Authentication of medicinal herbs using PCR-amplified ITS2 with specific primers. Planta Med. 2007, 73, 1421–1426. [Google Scholar] [CrossRef]
- Saini, A.; Reddy, S.K.; Jawali, N. Intra-individual and intra-species heterogeneity in nuclear rDNA ITS region of Vigna species from subgenus Ceratotropis. Genet. Res. (Camb) 2008, 90, 299–316. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three noncoding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Y.; Andolfatto, P. Methods to detect selection on noncoding DNA. Methods Mol. Biol. 2012, 856, 141–159. [Google Scholar]
- Fukuda, T.; Yokoyama, J.; Ohashi, H. Phylogeny and biogeography of the genus Lycium (Solanaceae): Inferences from chloroplast DNA sequences. Mol. Phylogenet. Evol. 2001, 19, 246–258. [Google Scholar] [CrossRef]
- Yi, T.; Miller, A.J.; Wen, J. Phylogenetic and biogeographic diversification of Rhus (Anacardiaceae) in the Northern Hemisphere. Mol. Phylogenet. Evol. 2004, 33, 861–879. [Google Scholar] [CrossRef]
- Kojoma, M.; Kurihara, K.; Yamada, K.; Sekita, S.; Satake, M.; Iida, O. Genetic identification of cinnamon (Cinnamomum spp.) based on the trnLtrnF chloroplast DNA. Planta Med. 2002, 68, 94–96. [Google Scholar] [CrossRef]
- Tsai, L.C.; Yu, Y.C.; Hsieh, H.M.; Wang, J.C.; Linacre, A.; Lee, J.C. Species identification using sequences of the trnL intron and the trnL-trnF IGS of chloroplast genome among popular plants in Taiwan. Forensic Sci. Int. 2006, 164, 193–200. [Google Scholar] [CrossRef]
- Lee, S.C.; Chiou, S.J.; Yen, J.H.; Lin, T.Y.; Hsieh, K.T.; Yang, J.C. DNA barcoding Cinnamomum osmophloeum Kaneh. based on the partial noncoding ITS2 region of ribosomal genes. J. Food Drug Anal. 2010, 18, 128–135. [Google Scholar]
- Lee, S.C.; Lee, C.H.; Lin, M.Y.; Ho, K.Y. Genetic identification of Cinnamomum species based on partial internal transcribed spacer 2 of ribosomal DNA. J. Food Drug Anal. 2010, 18, 225–231. [Google Scholar]
- Cubero, O.F.; Crespo, A.; Fatehi, J.; Bridge, P.D. DNA extraction and PCR amplification method suitable for fresh, herbarium-stored, lichenized, and other fungi. Plant Syst. Evol. 1999, 216, 243–249. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Yip, P.Y.; Chau, C.F.; Mak, C.Y.; Kwan, H.S. DNA methods for identification of Chinese medicinal materials. Chin. Med. 2007, 2, 9. [Google Scholar] [CrossRef]
- Yao, H.; Song, J.; Liu, C.; Luo, K.; Han, J.; Li, Y.; Pang, X.; Xu, H.; Zhu, Y.; Xiao, P.; et al. Use of ITS2 region as the universal DNA barcode for plants and animals. PLoS ONE 2010, 5, e13102. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yao, H.; Han, J.; Liu, C.; Song, J.; Shi, L.; Zhu, Y.; Ma, X.; Gao, T.; Pang, X.; et al. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS ONE 2010, 5, e8613. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (Code and Location) | GenBank Accession Number (Length: bp) | ||
|---|---|---|---|
| pITS2 | trnL Intron | trnL-trnF IGS | |
| (A) Present Study | |||
| Cinnamomum osmophloeum Kaneh. | |||
| 1. CO-B2 1 | A (170) | A 10 (464) | A 12 (356) |
| 2. CO-C1 | A (158) | ||
| 3. CO-D6 | A (158) | ||
| 4. CO-G19 | A (158) | ||
| 5. CO-K | A (158) | ||
| 6. CO-L | A (158) | ||
| 7. CO-LL | A (161) | ||
| 8. CO-M09 | A (162) | ||
| 9. CO-P2 | A (162) | ||
| 10. CO-S4 | A (167) | ||
| 11. CO-SE3 | A (162) | ||
| 12. CO-SP1 | A (162) | ||
| 13. CO-T3 | A (162) | ||
| 14. CO-Z | A (162) | ||
| 15. CO-002 2 | A (170) | ||
| 16. CO-003 | A 3 (170) | ||
| 17. CO-004 | A (170) | A (464) | |
| 18. CO-007 | A (170) | ||
| 19. CO-008 | A (170) | A 11 (464) | |
| 20. CO-009 | A (170) | ||
| 21. CO-010 | A 4 (170) | ||
| 22. CO-011 | A (170) | ||
| 23. CO-014 | A (170) | ||
| 24. CO-016 | A 5 (170) | ||
| 25. CO-021 | A (170) | ||
| 26. CO-023 | A (170) | ||
| 27. CO-024 | A 6 (170) | ||
| 28. CO-025 | A 7 (170) | ||
| 29. CO-028 | A 8 (170) | A (465) | |
| 30. CO-030 | A (171) | ||
| 31. CO-031 | A 13 (357) | ||
| 32. CO-033 | A (170) | ||
| 33. CO-038 | A (170) | ||
| 34. CO-040 | A (170) | ||
| 35. CO-042 | A 9 (170) | ||
| 36. CO-051 | A (356) | ||
| 37. CO-052 | A (357) | ||
| 38. CO-062 | A (358) | ||
| 39. CO-067 | A (464) | A 14 (361) | |
| 40. CO-069 | A (361) | ||
| 41. CO-071 | A (464) | ||
| 42. CO-083 | A (464) | ||
| 43. CO-097 | A (464) | ||
| 44. CO-098 | A (465) | ||
| 45. CO-102 | A (170) | ||
| 46. CO-105 | A (171) | ||
| 47. CO-122 | A (170) | ||
| 48. CO-123 | A (170) | ||
| 49. CO-124 | A (464) | ||
| (B) GenBank | |||
| C. burmannii | AB040087 (464) | AB040077 (349) | |
| C. burmannii-CB(TW-Nantou) | GQ255636 (170) | A 15 (464) | A 15 (356) |
| C. burmannii (CN-YZR) | DQ822590 (464) | ||
| C. camphora | AF272260 (169) | AB040091 (464) | AB040081 (349) |
| C. camphora var. glaucescens | AB040090 (464) | ||
| C. Cassia | AB040085 (464) | ||
| C. Cassia-CC1(CN-Fukan) | GQ255627 (169) | A 15 (464) | A 15 (356) |
| C. Cassia-CC2(CN-Fukan) | GQ255628 (169) | A 15 (464) | A 15 (356) |
| C. Cassia-CC3(CN-Guangdong) | GQ255629 (169) | A 15 (465) | A 15 (356) |
| C. Cassia-CC4(CN-Guangdong) | GQ255630 (169) | A 15 (464) | A 15 (356) |
| C. Cassia-CC5(CN-Fukan) | GQ255631 (169) | A 15 (464) | A 15 (356) |
| C. cassia (v-015) | AB054228 (356) | ||
| C. cinnamomifolium | AF272262 (170) | ||
| C. daphnoides | AB040092 (464) | AB040082 (349) | |
| C. doederleinii | AB040089 (464) | AB040079 (349) | |
| C. porrectum (C. inunctum) (MID) | DQ822587 (464) | ||
| C. insulari-montanum (C. japonicum) | AF272263 (168) | AB040093 (464) | AB040083 (349) |
| C. insulari-montanum (CN-YZR) | DQ822589 (464) | ||
| C. loureiroi-CL(VN-Thanh Hoa) | GQ255632 (169) | A 15 (464) | A 15 (356) |
| C. micranthum | EU333285 (356) | ||
| C. micranthum(i-123) | AM050466 (328) | ||
| C. micranthum(TW-Fuli) | EU338493 (356) | ||
| C. micranthum(TW-Hsiukuluan) | EU338495 (356) | ||
| C. micranthum(TW-Tahu) | EU338491 (355) | ||
| C. micranthum(TW-Yuli) | EU338499 (356) | ||
| C. micranthum(TW-Yungfeng) | EU338497 (356) | ||
| C. dubium (C. multiflorum) (MID) | DQ822586 (464) | ||
| C. bejolghota (C. obtusifolium) (MID) | DQ822588 (464) | ||
| C. sieboldii (C. okinawense) | AB040094 (464) | AB040084 (349) | |
| C. oleifolium | AF272264 (170) | ||
| C. osmophloeum-D4(TW-Nantou) | GQ255635(170) | ||
| C. philippinense | AM050463 (328) | ||
| C. pittosporoides | DQ124269 (168) | ||
| C. quadrangulum | AF272265 (146) | ||
| C. tamala (MID) | DQ822585 (464) | ||
| C. sieboldii | AB040088 (464) | AB040078 (349) | |
| C. verum | AF272267 (171) | AF268709 (334) | |
| C. wilsonii(CN-YZR) | DQ822592 (464) | ||
| C. verum (C. zeylanicum) | AB040086 (464) | ||
| C. verum -CZ(SL-Colombo) | GQ255634 (167) | A 15 (464) | A 15 (356) |
| C. verum (v-005) | AB054243 (464) | AB054235 (356) | |
| C. verum (v-006) | AB054242 (464) | ||
| Primers 1 | Sequence (5′–3′) | Design |
|---|---|---|
| BEL-1 | GGDGCGGAKAHTGGCCYCCCGTGC 2 | [25] |
| BEL-3 | GACGCTTCTCCAGACTACAAT | [25] |
| c | CGAAATCGGTAGACGCTACG | [27] |
| d | GGGGATAGAGGGACTTGAAC | [27] |
| e | GGTTCAAGTCCCTCTATCCC | [27] |
| f | ATTTGAACTGGTGACACGAG | [27] |
| pITS2 | trnL Intron | trnL-trnF IGS | |
|---|---|---|---|
| (A) Present study | |||
| C. osmophloeum Kaneh. | |||
| 1 LHC | 3 158–170 | 464 | 356 |
| (4 Ave = 161.4) | (Ave = 464) | (Ave = 356) | |
| (5 ID = 0.310–0.872) | (ID = 1) | (ID = 1) | |
| (6 N = 14) | (N = 1) | (N = 1) | |
| 2 HL | 170–171 | 464–465 | 356–361 |
| (Ave = 170.1) | (Ave = 464.2) | (Ave = 358.3) | |
| (ID = 0.872––0.994) | (ID = 0.995–0.997) | (ID = 0.969–0.997) | |
| (N = 24) | (N = 9) | (N = 6) | |
| LHC + HL | 158–171 | 464–465 | 356–361 |
| (Ave = 168.3) | (Ave = 464.1) | (Ave = 357.8) | |
| (ID = 0.310–0.994) | (ID = 0.995–0.997) | (ID = 0.969–0.997) | |
| (N = 38) | (N = 10) | (N = 7) | |
| (B) Cinnamomum spp. | |||
| (GenBank) | 146–171 | 464–465 | 328–356 |
| (Ave = 167.7) | (Ave = 464.0) | (Ave = 351.1) | |
| (ID = 0.721–1) | (ID = 0.993–1) | (ID = 0.418–1) | |
| (N = 16) | (N = 27) | (N = 26) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, W.-K.; Lee, S.-C.; Lu, P.-L. A Useful Technical Application of the Identification of Nucleotide Sequence Polymorphisms and Gene Resources for Cinnamomum osmophloeum Kaneh. (Lauraceae). Forests 2019, 10, 306. https://doi.org/10.3390/f10040306
Hsu W-K, Lee S-C, Lu P-L. A Useful Technical Application of the Identification of Nucleotide Sequence Polymorphisms and Gene Resources for Cinnamomum osmophloeum Kaneh. (Lauraceae). Forests. 2019; 10(4):306. https://doi.org/10.3390/f10040306
Chicago/Turabian StyleHsu, Wen-Kuang, Shih-Chieh Lee, and Pei-Luen Lu. 2019. "A Useful Technical Application of the Identification of Nucleotide Sequence Polymorphisms and Gene Resources for Cinnamomum osmophloeum Kaneh. (Lauraceae)" Forests 10, no. 4: 306. https://doi.org/10.3390/f10040306
APA StyleHsu, W. -K., Lee, S. -C., & Lu, P. -L. (2019). A Useful Technical Application of the Identification of Nucleotide Sequence Polymorphisms and Gene Resources for Cinnamomum osmophloeum Kaneh. (Lauraceae). Forests, 10(4), 306. https://doi.org/10.3390/f10040306