Effects of Lime Application and Understory Removal on Soil Microbial Communities in Subtropical Eucalyptus L’Hér. Plantations


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experiment Design
2.3. Soil Sampling and Analysis
2.4. Litter Decomposition Experiment
2.5. Statistical Analysis
3. Results
3.1. Soil Physico-Chemical Properties
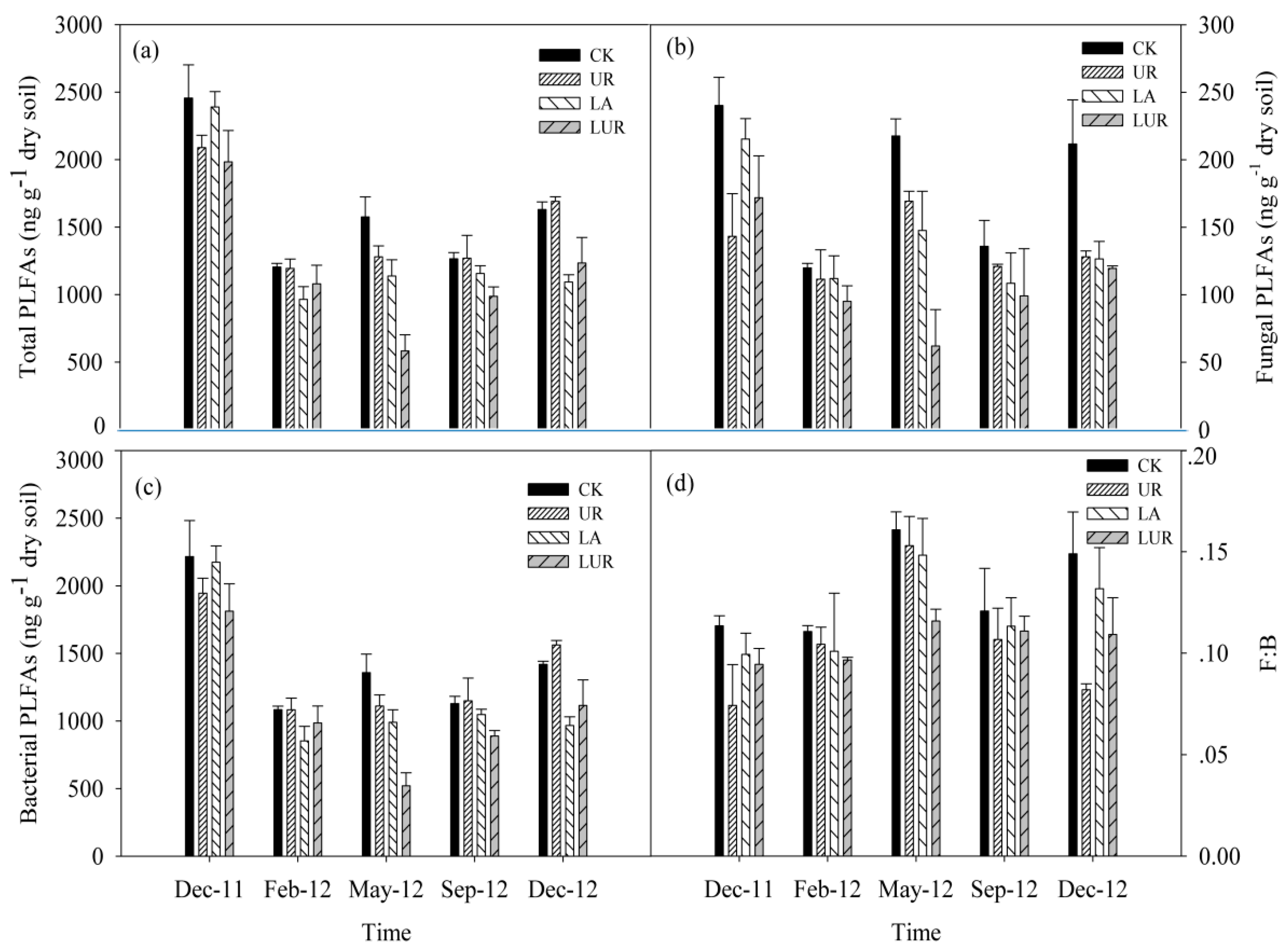
3.2. Soil Microbial Community
3.3. Relationships between Soil Physico-Chemical Properties and Microbial Community
3.4. Litter Decomposition
4. Discussion
4.1. The Impacts of Lime Application on Soil Microbial Community
4.2. The Impacts of Understory Removal on Soil Microbial Community
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wan, S.; Zhang, C.; Chen, Y.; Zhao, J.; Wang, X.; Wu, J.; Zhou, L.; Lin, Y.; Liu, Z.; Fu, S. The understory fern Dicranopteris dichotoma facilitates the overstory Eucalyptus trees in subtropical plantations. Ecosphere 2014, 5, 1–12. [Google Scholar] [CrossRef]
- Leff, J.W.; Jones, S.E.; Prober, S.M.; Barberán, A.; Borer, E.T.; Firn, J.L.; Harpole, W.S.; Hobbie, S.E.; Hofmockel, K.S.; Knops, J.M.H.; et al. Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proc. Natl. Acad. Sci. USA 2015, 112, 10967–10972. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [Green Version]
- Narendrula-Kotha, R.; Nkongolo, K.K. Microbial response to soil liming of damaged ecosystems revealed by pyrosequencing and phospholipid fatty acid analyses. PLoS ONE 2017, 12, e0168497. [Google Scholar] [CrossRef]
- Li, J.; Li, Z.; Wang, F.; Zou, B.; Chen, Y.; Zhao, J.; Mo, Q.; Li, Y.; Li, X.; Xia, H. Effects of nitrogen and phosphorus addition on soil microbial community in a secondary tropical forest of China. Biol. Fertil. Soils 2015, 51, 207–215. [Google Scholar] [CrossRef]
- Zhao, J.; Wan, S.; Fu, S.; Wang, X.; Wang, M.; Liang, C.; Chen, Y.; Zhu, X. Effects of understory removal and nitrogen fertilization on soil microbial communities in Eucalyptus plantations. For. Ecol. Manag. 2013, 310, 80–86. [Google Scholar] [CrossRef]
- Zhao, J.; Zhao, C.; Wan, S.; Wang, X.; Zhou, L.; Fu, S. Soil nematode assemblages in an acid soil as affected by lime application. Nematology 2015, 17, 179. [Google Scholar] [CrossRef]
- Bramble, D.S.E.; Gouveia, G.A.; Ramnarine, R. Organic residues and ammonium effects on CO2 emissions and soil quality indicators in limed acid tropical soils. Soil Syst. 2019, 3, 16. [Google Scholar] [CrossRef]
- Bakina, L.G.; Chugunova, M.V.; Zaitseva, T.B.; Nebol’sina, Z.P. The effect of liming on the complex of soil microorganisms and the humus status of a soddy-podzolic soil in a long-term experiment. Eurasian Soil Sci. 2014, 47, 110–118. [Google Scholar] [CrossRef]
- Pennanen, T.H.; Vanhala, P.; Kiikkila, O.; Neuvonen, S.; Baath, E. Structure of a microbial community in soil after prolonged addition of low levels of simulated acid rain. Appl. Environ. Microb. 1998, 64, 2173–2180. [Google Scholar]
- Pawlett, M.; Hopkins, D.W.; Moffett, B.F.; Harris, J.A. The effect of earthworms and liming on soil microbial communities. Biol. Fertil. Soils 2009, 45, 361–369. [Google Scholar] [CrossRef]
- Bruneau, P.M.C.; Davidson, D.A.; Grieve, I.C.; Young, I.M.; Naoise, N. The effects of soil horizons and faunal excrement on bacterial distribution in an upland grassland soil. Fems Microb. Ecol. 2010, 52, 139–144. [Google Scholar] [CrossRef]
- Kamal, S.; Prasad, R.; Varma, A. Soil microbial diversity in relation to heavy metals. In Soil Heavy Metals; Springer: Berlin, Heidelberg, 2010; volume 19, pp. 31–63. [Google Scholar]
- Wang, Y.; Shi, J.; Wang, H.; Lin, Q.; Chen, X.; Chen, Y. The influence of soil heavy metals pollution on soil microbial biomass, enzyme activity, and community composition near a copper smelter. Ecotoxicol. Environ. Saf. 2007, 67, 75–81. [Google Scholar] [CrossRef]
- Haynes, R.J.; Naidu, R. Influence of lime, fertilizer and manure applications on soil organic matter content and soil physical conditions: A review. Nutr. Cycl. Agroecosyst. 1998, 51, 123–137. [Google Scholar] [CrossRef]
- Shi, L.; Guo, Z.; Liang, F.; Xiao, X.; Peng, C.; Zeng, P.; Feng, W.; Ran, H. Effect of liming with various water regimes on both immobilization of Cadmium and improvement of bacterial communities in Contaminated Paddy: A field experiment. Int. J. Environ. Res. Public Health 2019, 16, 498. [Google Scholar] [CrossRef]
- Wan, S.; Zhang, C.; Chen, Y.; Zhao, J.; Zhu, X.; Wu, J.; Zhou, L.; Lin, Y.; Liu, Z.; Fu, S. Interactive effects of understory removal and fertilization on soil respiration in subtropical Eucalyptus plantations. J. Plant Ecol. 2015, 3, 284–290. [Google Scholar] [CrossRef]
- Wang, F.; Chen, F.; Wang, G.G.; Mao, R.; Fang, X.; Wang, H.; Bu, W. Effects of experimental nitrogen addition on nutrients and nonstructural carbohydrates of dominant understory plants in a Chinese fir plantation. Forests 2019, 10, 155. [Google Scholar] [CrossRef]
- Wu, J.; Liu, Z.; Chen, D.; Huang, G.; Zhou, L.; Fu, S. Understory plants can make substantial contributions to soil respiration: Evidence from two subtropical plantations. Soil Biol. Biochem. 2011, 43, 2355–2357. [Google Scholar] [CrossRef]
- Giuggiola, A.; Zweifel, R.; Feichtinger, L.M.; Vollenweider, P.; Bugmann, H.; Haeni, M.; Rigling, A. Competition for water in a xeric forest ecosystem—Effects of understory removal on soil micro-climate, growth and physiology of dominant Scots pine trees. For. Ecol. Manag. 2018, 409, 241–249. [Google Scholar] [CrossRef]
- Nilsson, M.-C.; Wardle, D.A. Understory vegetation as a forest ecosystem driver: Evidence from the northern Swedish boreal forest. Front. Ecol. Environ. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Wang, J.; Hai, R.; Long, Y.; Duan, W. Establishment and early growth of introduced indigenous tree species in typical plantations and shrubland in South China. For. Ecol. Manag. 2009, 7, 1293–1300. [Google Scholar] [CrossRef]
- Li, Y.L.; Wang, S.L.; Yan, S.K. Short-term effects of understory vegetation removal on nutrient cycling in litter layer of Chinese fir plantation. Chin. J. Appl. Ecol. 2011, 22, 2560. [Google Scholar]
- Wu, J.; Liu, Z.; Wang, X.; Sun, Y.; Zhou, L.; Lin, Y.; Fu, S. Effects of understory removal and tree girdling on soil microbial community composition and litter decomposition in two Eucalyptus plantations in South China. Funct. Ecol. 2011, 25, 921–931. [Google Scholar] [CrossRef] [Green Version]
- Urcelay, C.; Díaz, S.; Gurvich, D.E.; Chapin, F.S., III; Cuevas, E.; Domínguez, L.S. Mycorrhizal community resilience in response to experimental plant functional type removals in a woody ecosystem. J. Ecol. 2010, 97, 1291–1301. [Google Scholar] [CrossRef]
- FAO. World Referernce Base for Soil Resources 2006; World Soil Resources Report 103; FAO: Rome, Italy, 2006. [Google Scholar]
- Bossio, D.; Scow, K. Impacts of carbon and flooding on soil microbial communities: Phospholipid fatty acid profiles and substrate utilization patterns. Microb. Ecol. 1998, 35, 265–278. [Google Scholar] [CrossRef]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Use and misuse of PLFA measurements in soils. Soil Biol. Biochem. 2011, 43, 1621–1625. [Google Scholar] [CrossRef]
- Frostegård, Å.; Bååth, E.; Tunlio, A. Shifts in the structure of soil microbial communities in limed forests as revealed by phospholipid fatty acid analysis. Soil Biol. Biochem. 1993, 25, 723–730. [Google Scholar] [CrossRef]
- Lin, Y.; Ye, G.; Liu, D.; Ledgard, S.; Luo, J.; Fan, J.; Yuan, J.; Chen, Z.; Ding, W. Long-term application of lime or pig manure rather than plant residues suppressed diazotroph abundance and diversity and altered community structure in an acidic Ultisol. Soil Biol. Biochem. 2018, 123, 218–228. [Google Scholar] [CrossRef]
- Xue, D.; Huang, X.; Yao, H.; Huang, C. Effect of lime application on microbial community in acidic tea orchard soils in comparison with those in wasteland and forest soils. J. Environ. Sci. 2010, 22, 1253–1260. [Google Scholar] [CrossRef]
- Thirukkumaran, C.M.; Parkinson, D. Microbial respiration, biomass, metabolic quotient and litter decomposition in a lodgepole pine forest floor amended with nitrogen and phosphorous fertilizers. Soil Biol. Biochem. 2000, 32, 59–66. [Google Scholar] [CrossRef]
- Cox, F.; Barsoum, N.; Lilleskov, E.A.; Bidartondo, M.I. Nitrogen availability is a primary determinant of conifer mycorrhizas across complex environmental gradients. Ecol. Lett. 2010, 13, 1103–1113. [Google Scholar] [CrossRef]
- Winsome, T.; Silva, L.C.R.; Scow, K.M.; Doane, T.A.; Powers, R.F.; Horwath, W.R. Plant-microbe interactions regulate carbon and nitrogen accumulation in forest soils. For. Ecol. Manag. 2017, 384, 415–423. [Google Scholar] [CrossRef]
- He, T.X.; Li, Y.P.; Zhang, F.Y.; Wang, Q.K. Effects of understory removal on soil respiration and microbial community composition structure in a Chinese fir plantation. Chin. J. Plant Ecol. 2015, 39, 797–806. [Google Scholar]
- Yin, K.; Zhang, L.; Chen, D.; Tian, Y.; Zhang, F.; Wen, M.; Yuan, C. Understory herb layer exerts strong controls on soil microbial communities in subtropical plantations. Sci. Rep. 2016, 6, 27066. [Google Scholar] [CrossRef]
- Zhao, J.; Wan, S.; Li, Z.A.; Shao, Y.; Xu, G.; Liu, Z.; Zhou, L.; Fu, S. Dicranopteris-dominated understory as major driver of intensive forest ecosystem in humid subtropical and tropical region. Soil Biol. Biochem. 2012, 49, 78–87. [Google Scholar] [CrossRef]
- Schuldt, A.; Bruelheide, H.; Buscot, F.; Assmann, T.; Erfmeier, A.; Klein, A.-M.; Ma, K.; Scholten, T.; Staab, M.; Wirth, C.; et al. Belowground top-down and aboveground bottom-up effects structure multitrophic community relationships in a biodiverse forest. Sci. Rep. 2017, 7, 4222. [Google Scholar] [CrossRef] [Green Version]
- Benítez, E.; Paredes, D.; Rodríguez, E.; Aldana, D.; González, M.; Nogales, R.; Campos, M.; Moreno, B. Bottom-up effects on herbivore-induced plant defences: A case study based on compositional patterns of rhizosphere microbial communities. Sci. Rep. 2017, 7, 6251. [Google Scholar] [CrossRef]
- Brown, S.; Lenart, M.; Mo, J.; Kong, G. Structure and Organic Matter Dynamics of a Human-Impacted Pine Forest in a MAB Reserve of Subtropical China. Biotropica 1995, 27, 276–289. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | SOC (g kg−1) | TN (g kg−1) | DOC (μg C g−1) | pH |
|---|---|---|---|---|
| Control | 14.43 (0.66) | 1.86 (0.32) | 288.64 (74.26) | 3.97 (0.04) |
| UR | 15.78 (1.82) | 1.72 (0.53) | 268.11 (88.39) | 4.05 (0.04) |
| LA | 17.06 (1.95) | 1.68 (0.37) | 249.22 (62.16) | 4.14 (0.02) |
| LUR | 15.64 (1.21) | 1.71 (0.47) | 246.54 (46.55) | 4.16 (0.01) |
| ANOVA | ns | ns | ns | ns |
| Factors | Time | LA | UR | LA × UR | |||||
|---|---|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | F | P | ||
| SMC | 15.14 | <0.01 | 20.45 | <0.01 | 9.4 | <0.01 | 2.56 | 0.112 | |
| ST | 12.15 | <0.01 | 3.57 | 0.09 | 36.4 | <0.01 | 1.05 | 0.34 | |
| DOC | 1.45 | 0.2 | 0.95 | 0.33 | 0.18 | 0.06 | 0.23 | 0.63 | |
| NH4+–N | 7.15 | <0.01 | 0.27 | 0.6 | 2.31 | 0.14 | 2.2 | 0.99 | |
| NO3––N | 26.01 | <0.01 | 27.13 | <0.01 | 13.31 | <0.01 | 0.99 | 0.32 | |
| Variable | Time | LA | UR | LA × UR | Time × LA | Time × UR | Time × LA × UR |
|---|---|---|---|---|---|---|---|
| P | P | P | P | P | P | P | |
| Total PLFAs | <0.01 | <0.01 | 0.02 | 0.65 | 0.64 | 0.02 | 0.81 |
| Bacteria PLFAs | <0.01 | <0.01 | 0.14 | 0.79 | 0.24 | 0.07 | 0.94 |
| Fungal PLFAs | <0.01 | <0.01 | <0.01 | 0.46 | 0.06 | 0.08 | 0.21 |
| F:B | <0.01 | 0.76 | 0.02 | 0.74 | 0.65 | 0.26 | 0.38 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, S.; Liu, Z.; Chen, Y.; Zhao, J.; Ying, Q.; Liu, J. Effects of Lime Application and Understory Removal on Soil Microbial Communities in Subtropical Eucalyptus L’Hér. Plantations. Forests 2019, 10, 338. https://doi.org/10.3390/f10040338
Wan S, Liu Z, Chen Y, Zhao J, Ying Q, Liu J. Effects of Lime Application and Understory Removal on Soil Microbial Communities in Subtropical Eucalyptus L’Hér. Plantations. Forests. 2019; 10(4):338. https://doi.org/10.3390/f10040338
Chicago/Turabian StyleWan, Songze, Zhanfeng Liu, Yuanqi Chen, Jie Zhao, Qin Ying, and Juan Liu. 2019. "Effects of Lime Application and Understory Removal on Soil Microbial Communities in Subtropical Eucalyptus L’Hér. Plantations" Forests 10, no. 4: 338. https://doi.org/10.3390/f10040338
APA StyleWan, S., Liu, Z., Chen, Y., Zhao, J., Ying, Q., & Liu, J. (2019). Effects of Lime Application and Understory Removal on Soil Microbial Communities in Subtropical Eucalyptus L’Hér. Plantations. Forests, 10(4), 338. https://doi.org/10.3390/f10040338