Influence of Drought on Foliar Water Uptake Capacity of Temperate Tree Species


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Set-Up
2.2. Measurements
2.3. Data-Processing and Terminology
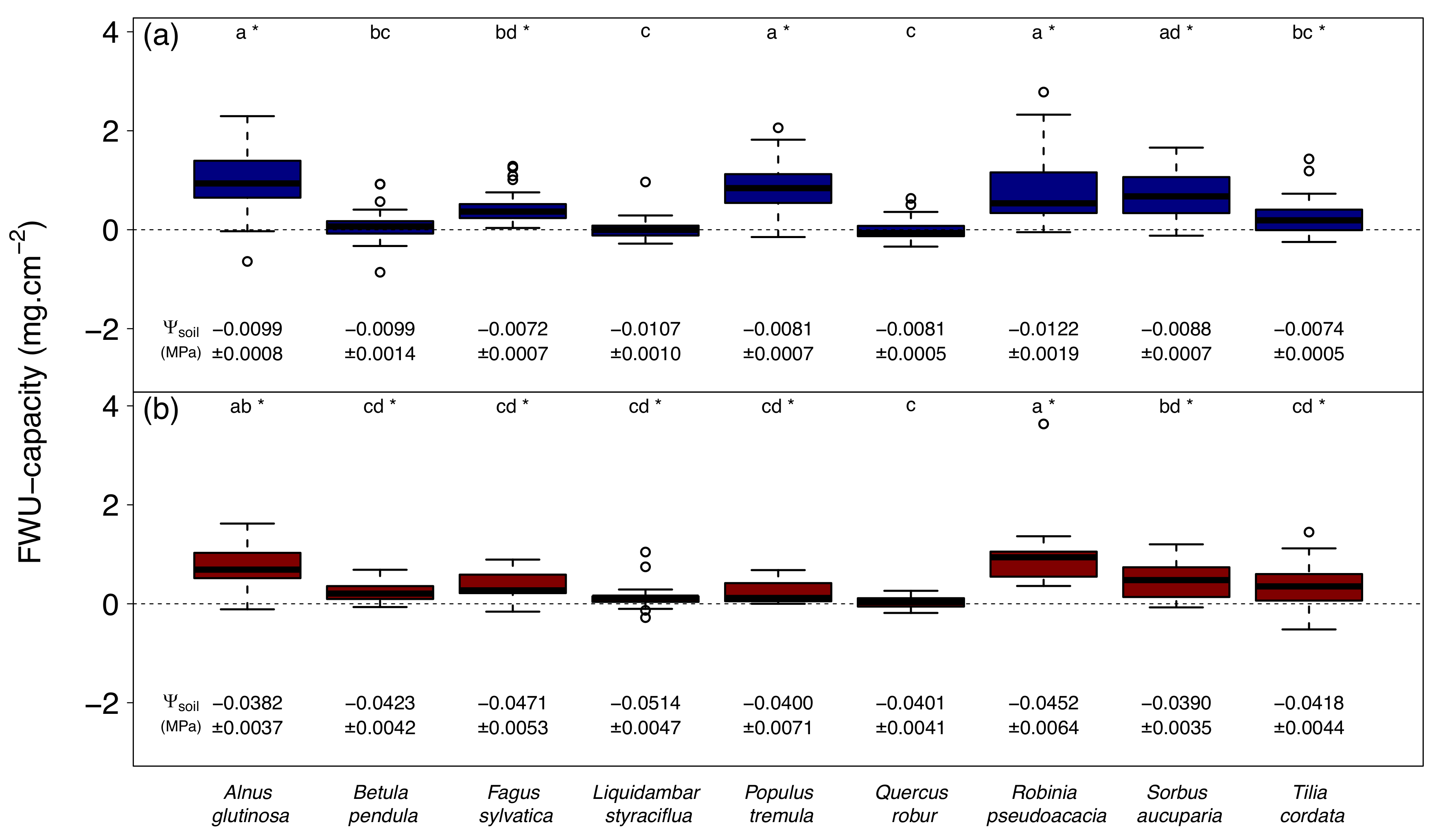
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dixon, H.H.; Joly, J. On the ascent of sap. Philos. Trans. R. Soc. London 1895, 186, 563–576. [Google Scholar]
- Nadezhdina, N.; David, T.S.; David, J.S.; Ferreira, M.I.; Dohnal, M.; Tesař, M.; Gartner, K.; Leitgeb, E.; Nadezhdin, V.; Cermak, J.; et al. Trees never rest: the multiple facets of hydraulic redistribution. Ecohydrology 2010, 3, 431–444. [Google Scholar] [CrossRef]
- Steppe, K.; Sterck, F.; Deslauriers, A. Diel growth dynamics in tree stems: Linking anatomy and ecophysiology. Trends Plant Sci. 2015, 20, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Berry, Z.C.; Emery, N.; Gotsch, S.G.; Goldsmith, G.R. Foliar water uptake: processes, pathways, and integration into plant water budgets. Plant. Cell Environ. 2018, 42, 410–423. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.E.; Goldsmith, G.R. The value of wet leaves. New Phytol. 2018, 219, 1156–1169. [Google Scholar] [CrossRef] [Green Version]
- Vesala, T.; Sevanto, S.; Grönholm, T.; Salmon, Y.; Nikinmaa, E.; Hari, P.; Hölttä, T. Effect of leaf water potential on internal humidity and CO2 dissolution: reverse transpiration and improved water use efficiency under negative pressure. Front. Plant Sci. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, G.R. Changing directions: The atmosphere-plant-soil continuum. New Phytol. 2013, 199, 4–6. [Google Scholar] [CrossRef]
- Breazeale, E.L.; Mcgeorge, W.T. Exudation pressure in roots of tomato plants under humid conditions. Soil 1953, 75, 293–298. [Google Scholar] [CrossRef]
- Stone, E.C. The ecological importance of dew. Q. Rev. Biol. 1963, 38, 328–341. [Google Scholar] [CrossRef]
- Steppe, K.; Vandegehuchte, M.W.; Van de Wal, B.A.E.; Hoste, P.; Guyot, A.; Lovelock, C.E.; Lockington, D.A. Direct uptake of canopy rainwater causes turgor-driven growth spurts in the mangrove Avicennia marina. Tree Physiol. 2018, 38, 979–991. [Google Scholar] [CrossRef]
- Breshears, D.D.; McDowell, N.G.; Goddard, K.L.; Dayem, K.E.; Martens, S.N.; Meyer, C.W.; Brown, K.M. Foliar absorption of intercepted rainfall improves woody plant water status most during drought. Ecology 2008, 89, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, J.; Hunsche, M. “Breath figures” on leaf surfaces—Formation and effects of microscopic leaf wetness. Front. Plant Sci. 2013, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Schreel, J.D.M.; Steppe, K. Foliar water uptake changes the world of tree hydraulics. npj Clim. Atmos. Sci. 2019, 2, 1. [Google Scholar] [CrossRef]
- Churakova Sidorova, O.V.; Saurer, M.; Bryukhanova, M.V.; Siegwolf, R.T.W.; Bigler, C. Site-specific water-use strategies of mountain pine and larch to cope with recent climate change. Tree Physiol. 2016, 36, 942–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonin, K.A.; Santiago, L.S.; Dawson, T.E. Fog interception by Sequoia sempervirens (D. Don) crowns decouples physiology from soil water deficit. Plant Cell Environ. 2009, 32, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Ohrui, T.; Nobira, H.; Sakata, Y.; Taji, T.; Yamamoto, C.; Nishida, K.; Yamakawa, T.; Sasuga, Y.; Yaguchi, Y.; Takenaga, H.; et al. Foliar trichome- and aquaporin-aided water uptake in a drought-resistant epiphyte Tillandsia ionantha Planchon. Planta 2007, 227, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.E.; von Willert, D.J. Leaf epidermal hydathodes and the ecophysiological consequences of foliar water uptake in species of Crassula from the Namib Desert in Southern Africa. Plant Biol. 2000, 2, 229–242. [Google Scholar] [CrossRef]
- Burkhardt, J.; Basi, S.; Pariyar, S.; Hunsche, M. Stomatal penetration by aqueous solutions - an update involving leaf surface particles. New Phytol. 2012, 196, 774–787. [Google Scholar] [CrossRef]
- Ketel, D.H.; Dirkse, W.G.; Ringoet, A. Water uptake from foliar-applied drops and its further distribution in the oat leaf. Acta Bot. Neerl. 1972, 21, 155–166. [Google Scholar] [CrossRef]
- Limm, E.B.; Simonin, K.A.; Bothman, A.G.; Dawson, T.E. Foliar water uptake: A common water acquisition strategy for plants of the redwood forest. Oecologia 2009, 161, 449–459. [Google Scholar] [CrossRef]
- Llamas, F.; Perez-Morales CAcedo, C.; Penas, A. Foliar trichomes of the evergreen and semi- deciduous species of the genus Quercus (Fagaceae) in the Iberian Peninsula. Bot. J. Linn. Soc. 1995, 117, 47–57. [Google Scholar] [CrossRef]
- Benz, B.W.; Martin, C.E. Foliar trichomes, boundary layers, and gas exchange in 12 species of epiphytic Tillandsia (Bromeliaceae). J. Plant Physiol. 2006, 163, 648–656. [Google Scholar] [CrossRef]
- Klockow, P.A.; Vogel, J.G.; Edgar, C.B.; Moore, G.W. Lagged mortality among tree species four years after an exceptional drought in east Texas. Ecosphere 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Hochberg, U.; Rockwell, F.E.; Holbrook, N.M.; Cochard, H. Iso/Anisohydry: a plant - environment interaction rather than a simple hydraulic trait. Trends Plant Sci. 2018, 23, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Abbruzzese, G.; Beritognolo, I.; Muleo, R.; Piazzai, M.; Sabatti, M.; Mugnozza, G.S.; Kuzminsky, E. Leaf morphological plasticity and stomatal conductance in three Populus alba L. genotypes subjected to salt stress. Environ. Exp. Bot. 2009, 66, 381–388. [Google Scholar] [CrossRef]
- Göransson, H.; Bambrick, M.T.; Godbold, D.L. Overyielding of temperate deciduous tree mixtures is maintained under throughfall reduction. Plant Soil 2016, 408, 285–298. [Google Scholar] [Green Version]
- Li, Q.; Wang, N.; Liu, X.; Liu, S.; Wang, H.; Zhang, W.; Wang, R.; Du, N. Growth and physiological responses to successional water deficit and recovery in four warm-temperate woody species. Physiol. Plant. 2019. [Google Scholar] [CrossRef]
- Vogt, U.K. Hydraulic vulnerability, vessel refilling, and seasonal courses of stem water potential of Sorbus aucuparia L. and Sambucus nigra L. J. Exp. Bot. 2001, 52, 1527–1536. [Google Scholar] [CrossRef]
- Stöhr, A.; Lösch, R. Xylem sap flow and drought stress of Fraxinus excelsior saplings. Tree Physiol. 2004, 24, 169–180. [Google Scholar]
- Moser, A.; Rötzer, T.; Pauleit, S.; Pretzsch, H. The urban environment can modify drought stress of small-leaved lime (Tilia cordata Mill.) and black locust (Robinia pseudoacacia L.). Forests 2016, 7. [Google Scholar] [CrossRef]
- Warren, J.M.; Norby, R.J.; Wullschleger, S.D. Elevated CO2 enhances leaf senescence during extreme drought in a temperate forest. Tree Physiol. 2011, 31, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Urli, M.; Lamy, J.-B.; Sin, F.; Burlett, R.; Delzon, S.; Porté, A.J. The high vulnerability of Quercus robur to drought at its southern margin paves the way for Quercus ilex. Plant Ecol. 2014, 216, 177–187. [Google Scholar] [CrossRef]
- Fu, X.; Meinzer, F.C. Metrics and proxies for stringency of regulation of plant water status (iso/anisohydry): A global data set reveals coordination and trade-offs among water transport traits. Tree Physiol. 2018, 39, 122–134. [Google Scholar] [CrossRef]
- Schreel, J.D.M.; Van de Wal, B.A.E.; Hervé-Fernandez, P.; Boeckx, P.; Steppe, K. Hydraulic redistribution of foliar absorbed water causes turgor-driven growth in mangrove seedlings. Plant. Cell Environ. 2019. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, G.R.; Lehmann, M.M.; Cernusak, L.A.; Arend, M.; Siegwolf, R.T.W. Inferring foliar water uptake using stable isotopes of water. Oecologia 2017, 184, 763–766. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| β1 | β0 | R2 | Terminology | |
|---|---|---|---|---|
| Significant (p < 0.05) |  | Significant | ||
| Not significant | Significant (p < 0.05) | | No effect | |
| Not significant | Not significant | ≥0.5 | | Tendency to |
| Not significant | Not significant | <0.5 | | No effect |
| Species. | LWCi | FWUmax | PRI | FWUi | ARI |
|---|---|---|---|---|---|
| Alnus glutinosa | 1.4 ± 0.0 | 1.9 ± 0.0 | 2.7 ± 0.0 | 0.9 ± 0.4 | 1.3 ± 0.6 |
| Betula pendula | 2.1 ± 0.0 | 0.9 ± 0.0 | 1.9 ± 0.0 | x | x |
| Fagus sylvatica | 3.1 ± 0.3 | 1.0 ± 0.0 | 3.3 ± 0.3 | 0.2 ± 0.1 | 0.6 ± 0.4 |
| Liquidambar styraciflua | 1.6 ± 0.0 | 0.3 ± 0.0 | 0.5 ± 0.0 | 0.1 ± 0.0 | 0.2 ± 0.0 |
| Populus tremula | 2.0 ± 0.0 | 1.6 ± 0.0 | 3.2 ± 0.0 | 0.3 ± 0.1 | 0.6 ± 0.2 |
| Quercus robur | 2.2 ± 0.1 | 0.6 ± 0.1 | 1.3 ± 0.3 | x | x |
| Robinia pseudoacacia | 1.7 ± 0.3 | 1.1 ± 0.1 | 1.9 ± 0.5 | 0.5 ± 0.0 | 0.9 ± 0.2 |
| Sorbus aucuparia | 1.7 ± 0.0 | 1.4 ± 0.0 | 2.3 ± 0.0 | x | x |
| Tilia cordata | 2.3 ± 0.1 | 1.4 ± 0.0 | 3.3 ± 0.1 | x | x |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schreel, J.D.M.; von der Crone, J.S.; Kangur, O.; Steppe, K. Influence of Drought on Foliar Water Uptake Capacity of Temperate Tree Species. Forests 2019, 10, 562. https://doi.org/10.3390/f10070562
Schreel JDM, von der Crone JS, Kangur O, Steppe K. Influence of Drought on Foliar Water Uptake Capacity of Temperate Tree Species. Forests. 2019; 10(7):562. https://doi.org/10.3390/f10070562
Chicago/Turabian StyleSchreel, Jeroen D.M., Jonas S. von der Crone, Ott Kangur, and Kathy Steppe. 2019. "Influence of Drought on Foliar Water Uptake Capacity of Temperate Tree Species" Forests 10, no. 7: 562. https://doi.org/10.3390/f10070562
APA StyleSchreel, J. D. M., von der Crone, J. S., Kangur, O., & Steppe, K. (2019). Influence of Drought on Foliar Water Uptake Capacity of Temperate Tree Species. Forests, 10(7), 562. https://doi.org/10.3390/f10070562