Chemosensory Characteristics of Two Semanotus bifasciatus Populations



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects for Antennal Transcriptome Sequencing
2.2. RNA-Seq Library Preparation and Sequencing
2.3. De Novo Assembly of the Sequences
2.4. Annotation
2.5. Gene Expression Quantification
2.6. SNP Calling
2.7. Data Analysis
3. Results
3.1. Antennal Transcriptomes Assembly and Annotation
3.2. Chemosensory Gene Identification of S. bifasciatus
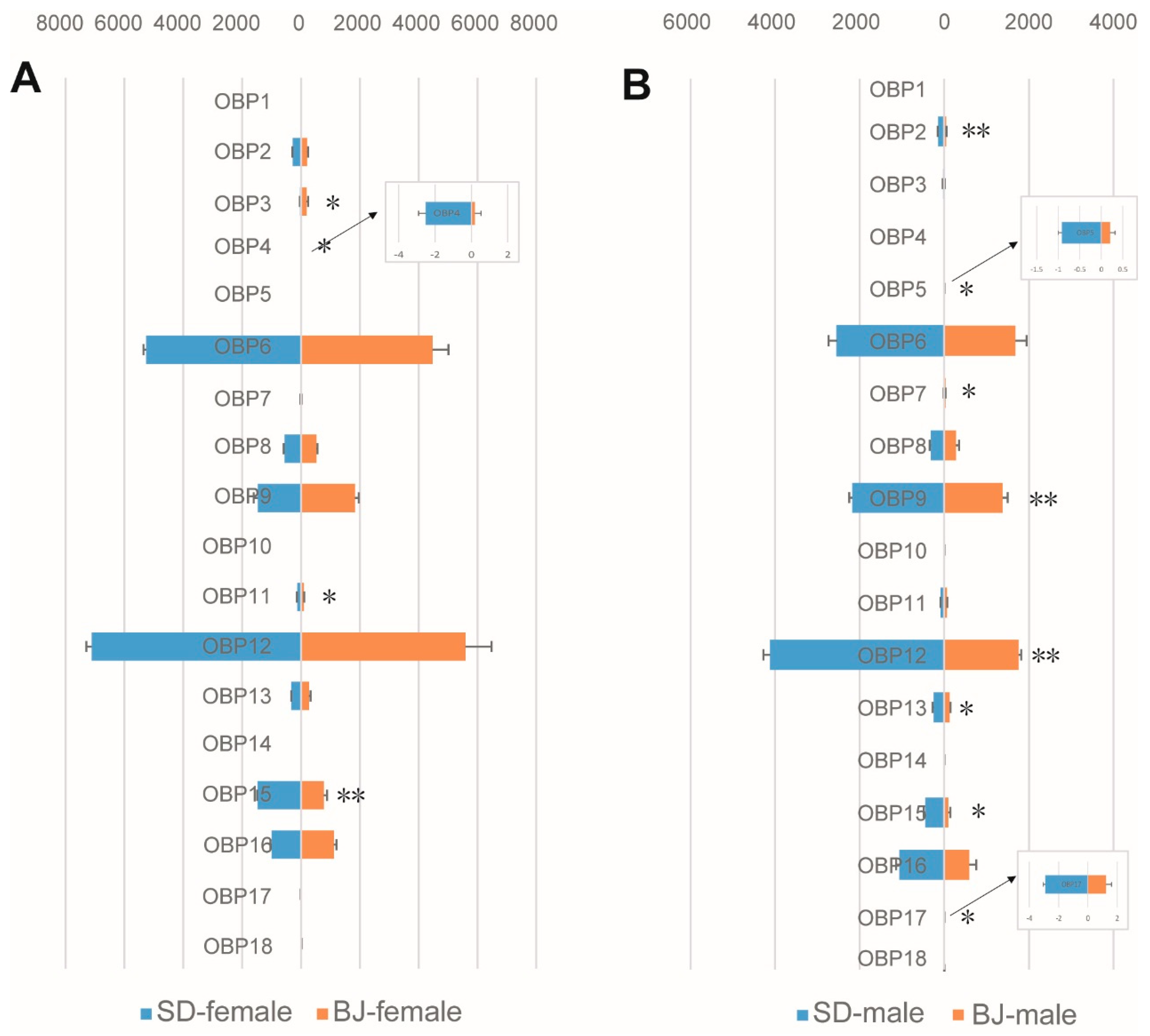
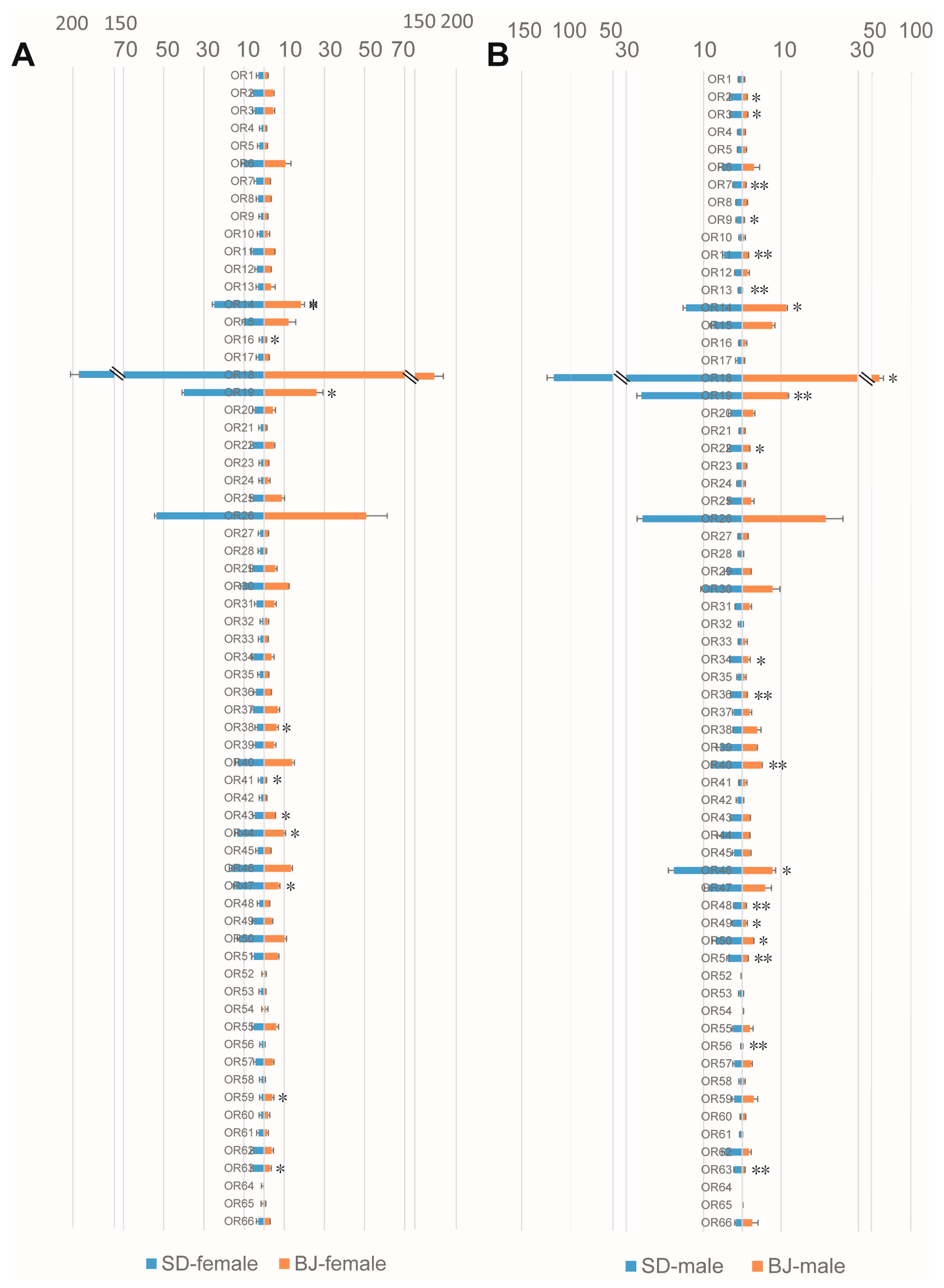
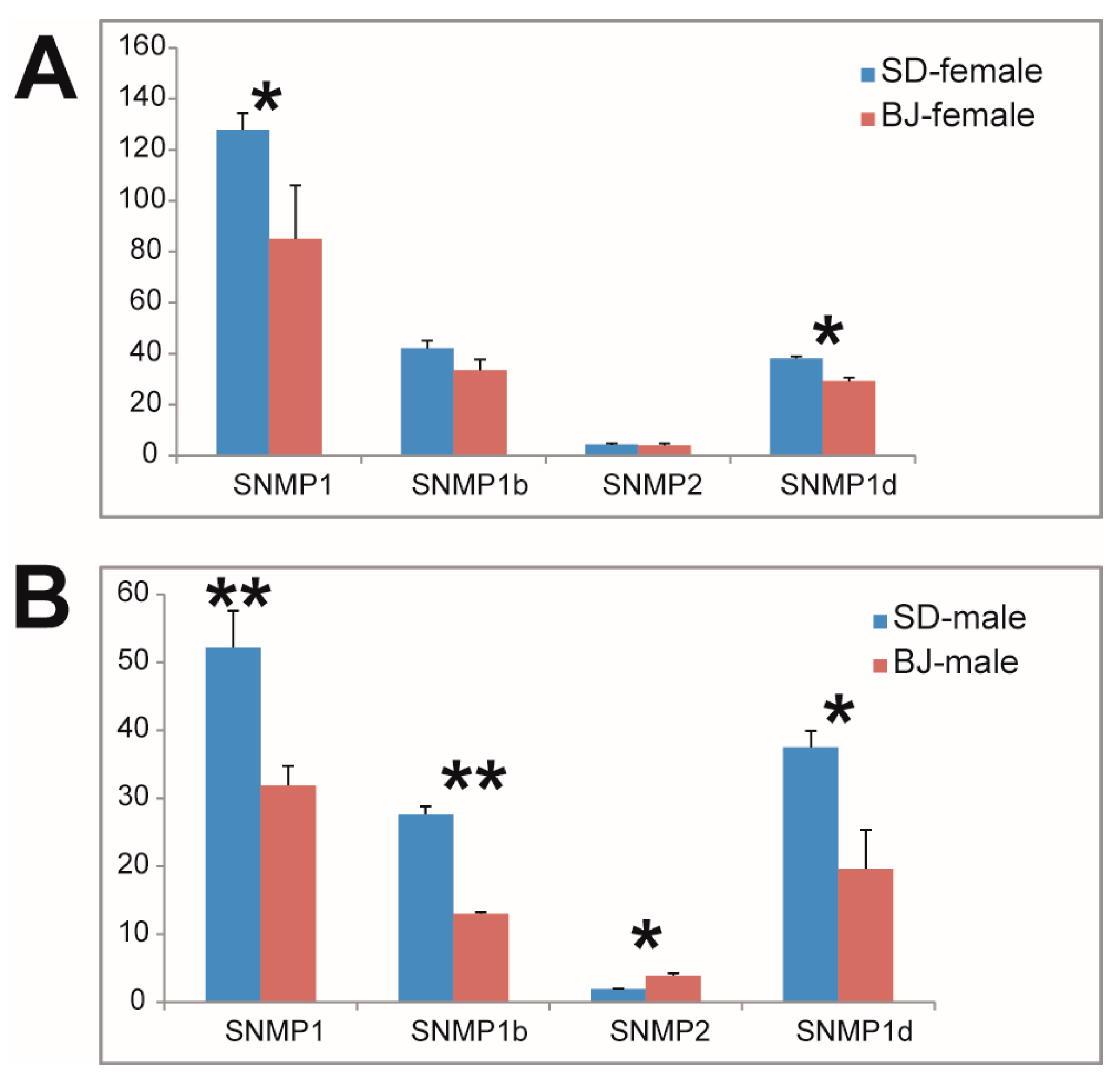
3.3. Chemosensory Gene Expression Characteristics of Two S. bifasciatus Populations
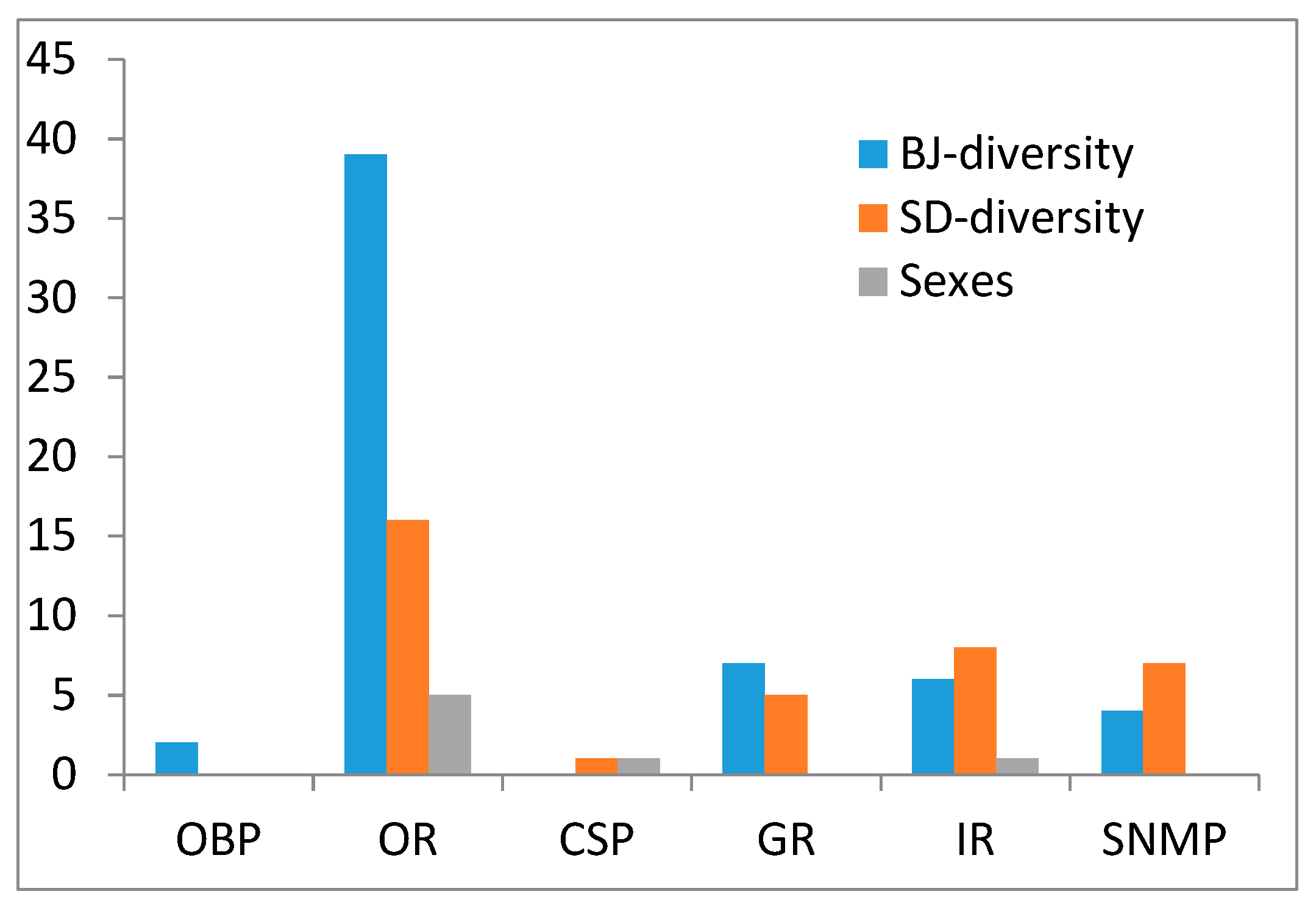
3.4. SNP Analysis of Chemosensory Genes in Two S. bifasciatus Populations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gao, S.; Xu, Z.; Gong, X. Progress in research on semanotus bifasciatus. For. Pest Dis. 2007, 26, 19–22. [Google Scholar]
- Wang, S. Quarantine Objects of Forest Plant in China; China Forestry Publishing House: Beijing, China, 1996. [Google Scholar]
- Wu, X.; Wang, J.; Liu, H.; Dong, l.; Jin, Y. Chemical analysis and electroantennogram responses in Semanotus bifasciatus adults to Platycladus orientalis. Chin. Bull. Entomol. 2007, 44, 671–675. [Google Scholar]
- Kong, X.; Zhang, Z.; Wang, H.; Yang, J.; Hu, Y. Analysis of Platycladus orientalis volatiles and their elecctroantennogram responses with Semanotus bifasciatus. For. Res. 2005, 18, 260–266. [Google Scholar]
- Sun, Y. Sensilla and Behavioral Responses of Adults of Semanotus bifasciatus (Coleopetera: Cerambycidae) to Volatile Compounds of Platycladus orientalis; Beijing Forest Universtiy: Beijing, China, 2008. [Google Scholar]
- Su, C.Y.; Menuz, K.; Carlson, J.R. Olfactory perception: Receptors, cells, and circuits. Cell 2009, 139, 45–59. [Google Scholar] [CrossRef]
- Vogt, R.G.; Miller, N.E.; Litvack, R.; Fandino, R.A.; Sparks, J.; Staples, J.; Friedman, R.; Dickens, J.C. The insect SNMP gene family. Insect Biochem. Mol. Biol. 2009, 39, 448–456. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, Z.; Wang, H.; Kong, X. Antennal transcriptome analysis and comparison of olfactory genes in two sympatric defoliators, Dendrolimus houi and Dendrolimus kikuchii (lepidoptera: Lasiocampidae). Insect Biochem. Mol. Boil. 2014, 52, 69–81. [Google Scholar] [CrossRef]
- Benton, R.; Vannice, K.S.; Gomez-Diaz, C.; Vosshall, L.B. Variant ionotropic glutamate receptors as chemosensory receptors in drosophila. Cell 2009, 136, 149–162. [Google Scholar] [CrossRef]
- Touhara, K.; Vosshall, L.B. Sensing odorants and pheromones with chemosensory receptors. Annu. Rev. Physiol. 2009, 71, 307–332. [Google Scholar] [CrossRef]
- Kaupp, U.B. Olfactory signaling in vertebrates and insects: Differences and commonalities. Nat. Rev. Neurosci. 2010, 11, 188–200. [Google Scholar] [CrossRef]
- Kwon, J.Y.; Dahanukar, A.; Weiss, L.A.; Carlson, J.R. The molecular basis of CO2 reception in drosophila. Proc. Natl. Acad. Sci. USA 2007, 104, 3574–3578. [Google Scholar] [CrossRef]
- Robertson, H.M.; Kent, L.B. Evolution of the gene lineage encoding the carbon dioxide receptor in insects. J. Insect Sci. 2009, 9, 19. [Google Scholar] [CrossRef]
- Clyne, P.J.; Warr, C.G.; Freeman, M.R.; Lessing, D.; Kim, J.; Carlson, J.R. A novel family of divergent seven-transmembrane proteins: Candidate odorant receptors in Drosophila. Neuron 1999, 22, 327–338. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Amrein, H.; Morozov, P.S.; Rzhetsky, A.; Axel, R. A spatial map of olfactory receptor expression in the Drosophila antenna. Cell 1999, 96, 725–736. [Google Scholar] [CrossRef]
- Hallem, E.A.; Carlson, J.R. Coding of odors by a receptor repertoire. Cell 2006, 125, 143–160. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Hansson, B.S. A unified nomenclature system for the insect olfactory coreceptor. Chem. Senses 2011, 36, 497–498. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Stocker, R.F. Molecular architecture of smell and taste in Drosophila. Annu. Rev. Neurosci. 2007, 30, 505–533. [Google Scholar] [CrossRef]
- Sanchez-Gracia, A.; Vieira, F.G.; Rozas, J. Molecular evolution of the major chemosensory gene families in insects. Heredity 2009, 103, 208–216. [Google Scholar] [CrossRef]
- Vogt, R.G. Biochemical diversity of odor detection: OBPs, ODEs and SNMPs. In Insect Pheromone Biochemistry and Molecular Biology; Academic Press: London, UK, 2003; pp. 391–446. [Google Scholar]
- Pelosi, P.; Zhou, J.J.; Ban, L.P.; Calvello, M. Soluble proteins in insect chemical communication. Cell. Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef]
- Engsontia, P.; Sanderson, A.P.; Cobb, M.; Walden, K.K.O.; Robertson, H.M.; Brown, S. The red flour beetle’s large nose: An expanded odorant receptor gene family in Tribolium castaneum. Insect Biochem. Mol. Biol. 2008, 38, 387–397. [Google Scholar] [CrossRef]
- Krieger, J.; Grosse-Wilde, E.; Gohl, T.; Dewer, Y.M.E.; Raming, K.; Breer, H. Genes encoding candidate pheromone receptors in a moth (Heliothis virescens). Proc. Natl. Acad. Sci. USA 2004, 101, 11845–11850. [Google Scholar] [CrossRef]
- Robertson, H.M.; Wanner, K.W. The chemoreceptor superfamily in the honey bee, Apis mellifera: Expansion of the odorant, but not gustatory, receptor family. Genome Res. 2006, 16, 1395–1403. [Google Scholar] [CrossRef]
- Tanaka, K.; Uda, Y.; Ono, Y.; Nakagawa, T.; Suwa, M.; Yamaoka, R.; Touhara, K. Highly selective tuning of a silkworm olfactory receptor to a key mulberry leaf volatile. Curr. Biol. 2009, 19, 881–890. [Google Scholar] [CrossRef]
- Gong, D.-P.; Zhang, H.-J.; Zhao, P.; Xia, Q.-Y.; Xiang, Z.-H. The odorant binding protein gene family from the genome of silkworm, Bombyx mori. BMC Genom. 2009, 10, 332. [Google Scholar] [CrossRef]
- Zhou, J.-J.; Kan, Y.; Antoniw, J.; Pickett, J.A.; Field, L.M. Genome and est analyses and expression of a gene family with putative functions in insect chemoreception. Chem. Senses 2006, 31, 453–465. [Google Scholar] [CrossRef]
- Zhou, J.J.; He, X.L.; Pickett, J.A.; Field, L.M. Identification of odorant-binding proteins of the yellow fever mosquito Aedes aegypti: Genome annotation and comparative analyses. Insect Mol. Biol. 2008, 17, 147–163. [Google Scholar] [CrossRef]
- Bengtsson, J.M.; Trona, F.; Montagné, N.; Anfora, G.; Ignell, R.; Witzgall, P.; Jacquin-Joly, E. Putative chemosensory receptors of the codling moth, Cydia pomonella, identified by antennal transcriptome analysis. PLoS ONE 2012, 7, e31620. [Google Scholar] [CrossRef]
- Grosse-Wilde, E.; Kuebler, L.S.; Bucks, S.; Vogel, H.; Wicher, D.; Hansson, B.S. Antennal transcriptome of Manduca sexta. Proc. Natl. Acad. Sci. USA 2011, 108, 7449–7454. [Google Scholar] [CrossRef]
- Legeai, F.; Malpel, S.; Montagne, N.; Monsempes, C.; Cousserans, F.; Merlin, C.; Francois, M.-C.; Maibeche-Coisne, M.; Gavory, F.; Poulain, J.; et al. An expressed sequence tag collection from the male antennae of the noctuid moth Spodoptera littoralis: A resource for olfactory and pheromone detection research. BMC Genom. 2011, 12, 86. [Google Scholar] [CrossRef]
- Zhang, Y.-N.; Jin, J.-Y.; Jin, R.; Xia, Y.-H.; Zhou, J.-J.; Deng, J.-Y.; Dong, S.-L. Differential expression patterns in chemosensory and non-chemosensory tissues of putative chemosensory genes identified by transcriptome analysis of insect pest the purple stem borer Sesamia inferens (walker). PLoS ONE 2013, 8, e69715. [Google Scholar]
- Mitchell, R.F.; Hughes, D.T.; Luetje, C.W.; Millar, J.G.; Soriano-Agatmn, F.; Hanks, L.M.; Robertson, H.M. Sequencing and characterizing odorant receptors of the cerambycid beetle Megacyllene caryae. Insect Biochem. Mol. Biol. 2012, 42, 499–505. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.; Grosse-Wilde, E.; Keeling, C.; Bengtsson, J.; Yuen, M.; Li, M.; Hillbur, Y.; Bohlmann, J.; Hansson, B.; Schlyter, F. Antennal transcriptome analysis of the chemosensory gene families in the tree killing bark beetles, Ips typographus and Dendroctonus ponderosae (coleoptera: Curculionidae: Scolytinae). BMC Genom. 2013, 14, 1–16. [Google Scholar] [CrossRef]
- Zhang, S.-F.; Liu, H.-H.; Kong, X.-B.; Wang, H.-B.; Liu, F.; Zhang, Z. Identification and expression profiling of chemosensory genes in Dendrolimus punctatus walker. Front. Physiol. 2017, 8, 8. [Google Scholar] [CrossRef]
- Bogdanova, E.A.; Shagin, D.A.; Lukyanov, S.A. Normalization of full-length enriched cDNA. Mol. BioSystems 2008, 4, 205–212. [Google Scholar] [CrossRef]
- Zhulidov, P.A.; Bogdanova, E.A.; Shcheglov, A.S.; Vagner, L.L.; Khaspekov, G.L.; Kozhemyako, V.B.; Matz, M.V.; Meleshkevitch, E.; Moroz, L.L.; Lukyanov, S.A.; et al. Simple cDNA normalization using kamchatka crab duplex-specific nuclease. Nucleic Acids Res. 2004, 32, e37. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-seq data. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B.; et al. Tigr gene indices clustering tools (tgicl): A software system for fast clustering of largest datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2go: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the blast2go suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential Expression of RNA-seq Data at the Gene Level—The Deseq Package; European Molecular Biology Laboratory (EMBL): Heidelberg, Germany, 2012. [Google Scholar]
- Bullard, J.H.; Purdom, E.; Hansen, K.D.; Dudoit, S. Evaluation of statistical methods for normalization and differential expression in mRNA-seq experiments. BMC Bioinform. 2010, 11, 94. [Google Scholar] [CrossRef]
- Noble, W.S. How does multiple testing correction work? Nat. Biotechnol. 2009, 27, 1135–1137. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A mapreduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- He, Y.Q.; Feng, B.; Guo, Q.S.; Du, Y. Age influences the olfactory profiles of the migratory oriental armyworm mythimna separate at the molecular level. BMC Genom. 2017, 18, 32. [Google Scholar] [CrossRef]
- Qiu, C.Z.; Zhou, Q.Z.; Liu, T.-T.; Fang, S.M.; Wang, Y.W.; Fang, X.; Huang, C.L.; Yu, Q.Y.; Chen, C.H.; Zhang, Z. Evidence of peripheral olfactory impairment in the domestic silkworms: Insight from the comparative transcriptome and population genetics. BMC Genom. 2018, 19, 788. [Google Scholar] [CrossRef]
- Gadenne, C.; Barrozo, R.B.; Anton, S. Plasticity in insect olfaction: To smell or not to smell? Annu. Rev. Èntomol. 2016, 61, 317–333. [Google Scholar] [CrossRef]
- Zhan, W.; Zhang, S.; Geng, H.; Wang, Y.; Guo, K.; Chen, J. Research progress on olfactory recognition proteins of longhorn beetle. J. Henan Agric. Sci. 2018, 47, 1–6. [Google Scholar]
- McKenna, D.D.; Scully, E.D.; Pauchet, Y.; Hoover, K.; Kirsch, R.; Geib, S.M.; Mitchell, R.F.; Waterhouse, R.M.; Ahn, S.-J.; Arsala, D.; et al. Genome of the Asian longhorned beetle (Anoplophora glabripennis), a globally significant invasive species, reveals key functional and evolutionary innovations at the beetle–plant interface. Genome Boil. 2016, 17, 227. [Google Scholar] [CrossRef]
- Wang, P.; Lyman, R.F.; Mackay, T.F.C.; Anholt, R.R.H. Natural variation in odorant recognition among odorant-binding proteins in drosophila melanogaster. Genetics 2010, 184, 759–767. [Google Scholar] [CrossRef]
- Wang, P.; Lyman, R.F.; Shabalina, S.A.; Mackay, T.F.C.; Anholt, R.R.H. Association of polymorphisms in odorant-binding protein genes with variation in olfactory response to benzaldehyde in drosophila. Genetics 2007, 177, 1655–1665. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Shen, S.; Zhang, S.; Wang, H.; Kong, X.; Liu, F.; Zhang, Z. Chemosensory Characteristics of Two Semanotus bifasciatus Populations. Forests 2019, 10, 655. https://doi.org/10.3390/f10080655
Zhang S, Shen S, Zhang S, Wang H, Kong X, Liu F, Zhang Z. Chemosensory Characteristics of Two Semanotus bifasciatus Populations. Forests. 2019; 10(8):655. https://doi.org/10.3390/f10080655
Chicago/Turabian StyleZhang, Sufang, Sifan Shen, Shiyu Zhang, Hongbin Wang, Xiangbo Kong, Fu Liu, and Zhen Zhang. 2019. "Chemosensory Characteristics of Two Semanotus bifasciatus Populations" Forests 10, no. 8: 655. https://doi.org/10.3390/f10080655
APA StyleZhang, S., Shen, S., Zhang, S., Wang, H., Kong, X., Liu, F., & Zhang, Z. (2019). Chemosensory Characteristics of Two Semanotus bifasciatus Populations. Forests, 10(8), 655. https://doi.org/10.3390/f10080655