Survival, Height Growth, and Phytoextraction Potential of Hybrid Poplar and Russian Olive (Elaeagnus Angustifolia L.) Established on Soils Varying in Salinity in North Dakota, USA


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site and Soils Description
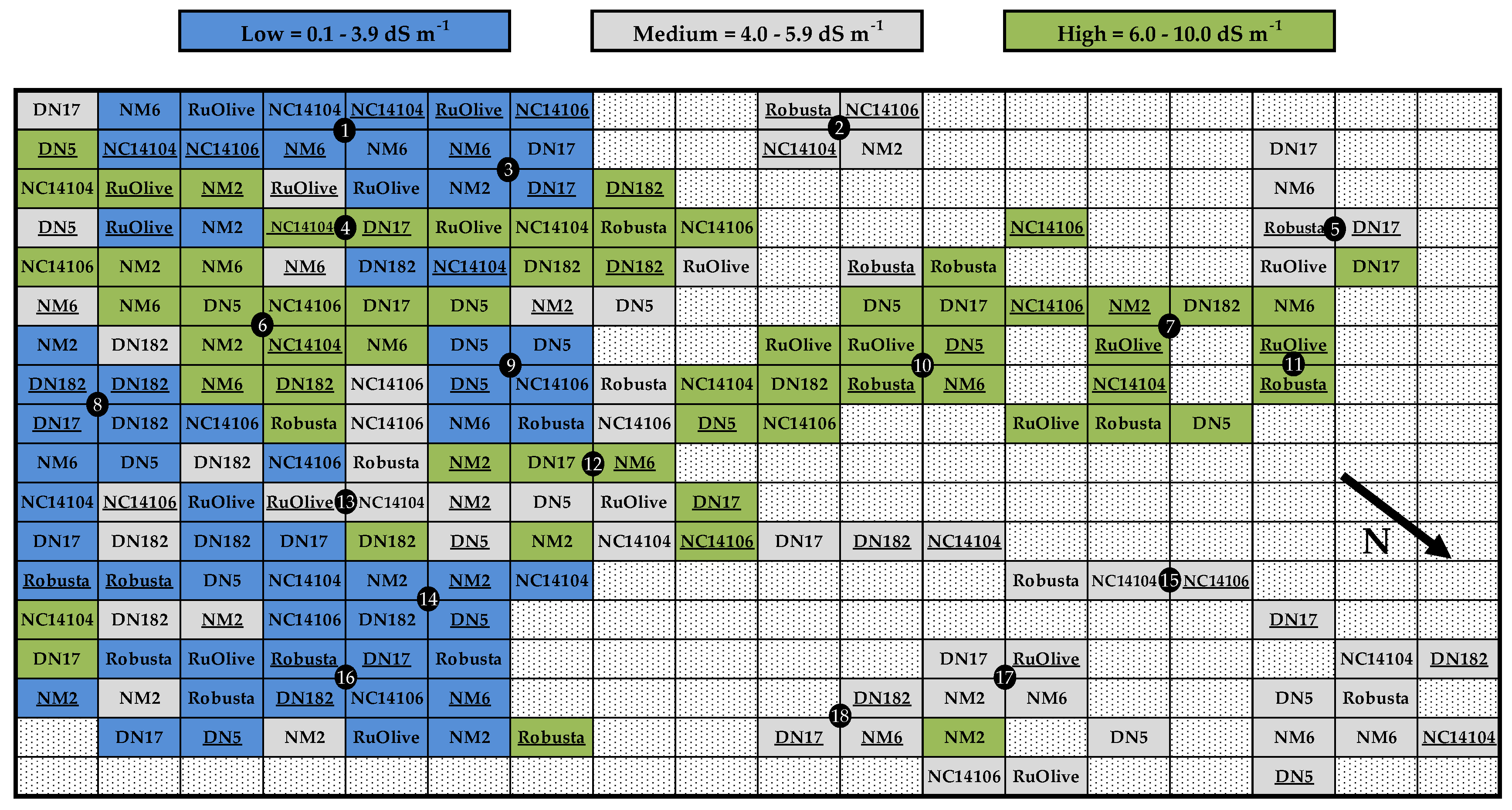
2.2. Plant Material and Experimental Design
2.3. Data Collection and Analysis
3. Results
3.1. Survival
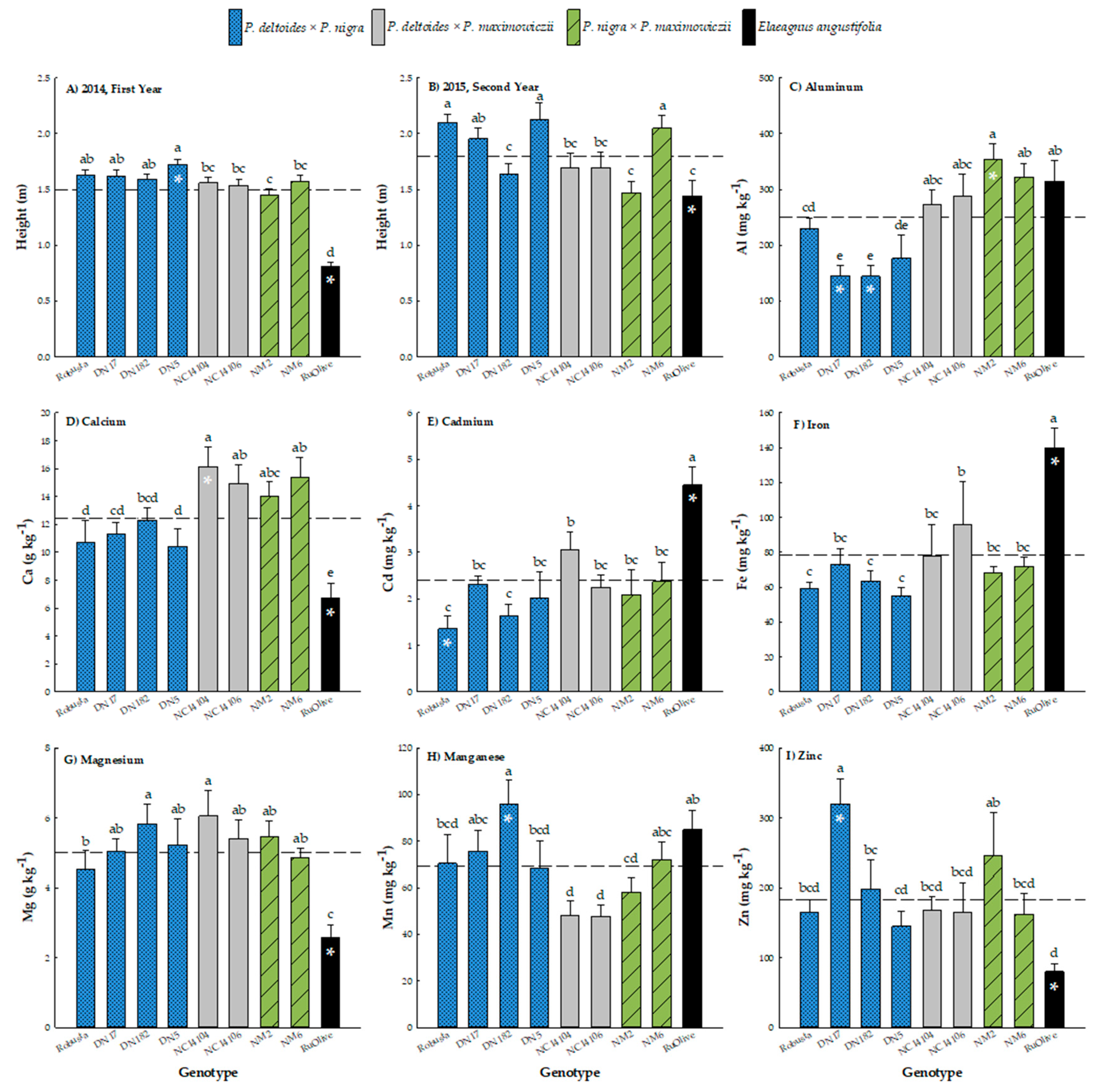
3.2. Height Growth
3.3. Leaf Phytoextraction
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schultz, R.C.; Colletti, J.P.; Faltonson, R.R. Agroforestry opportunities for the United States of America. Agrofor. Syst. 1995, 31, 117–132. [Google Scholar] [CrossRef]
- Franzen, D. Managing saline soils in North Dakota. North Dak. State Univ. Ext. Bull. 2013, SF1087, 11. [Google Scholar]
- Monk, R.W.; Wiebe, H.H. Salt tolerance and protoplasmic salt hardiness of various woody and herbaceous ornamental plants. Plant Physiol. 1961, 36, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Khamzina, A.; Lamers, J.P.; Vlek, P.L. Nitrogen fixation by Elaeagnus angustifolia in the reclamation of degraded croplands of Central Asia. Tree Physiol. 2009, 29, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Enescu, C.M. Russian olive (Elaeagnus angustifolia L.): A multipurpose species with an important role in land reclamation. Curr. Trends Nat. Sci. 2018, 7, 54–60. Available online: http://www.natsci.upit.ro/media/1642/paper-8.pdf (accessed on 5 April 2019).
- Katz, G.L.; Shafroth, P.B. Biology, ecology and management of Elaeagnus angustifolia L. (Russian olive) in western North America. Wetlands 2003, 23, 763–777. [Google Scholar] [CrossRef]
- Riemenschneider, D.E.; Berguson, W.E.; Dickmann, D.I.; Hall, R.B.; Isebrands, J.G.; Mohn, C.A.; Stanosz, G.R.; Tuskan, G.A. Poplar breeding and testing strategies in the north-central U.S.: Demonstration of potential yield and consideration of future research needs. For. Chron. 2001, 77, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Zalesny, R.S., Jr.; Hall, R.B.; Zalesny, J.A.; Berguson, W.E.; McMahon, B.G.; Stanosz, G.R. Biomass and genotype × environment interactions of Populus energy crops in the Midwestern United States. BioEnergy Res. 2009, 2, 106–122. [Google Scholar] [CrossRef]
- Nelson, N.D.; Berguson, W.E.; McMahon, B.G.; Cai, M.; Buchman, D.J. Growth performance and stability of hybrid poplar clones in simultaneous tests on six sites. Biomass Bioenergy 2018, 118, 115–125. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Stanturf, J.A.; Gardiner, E.S.; Perdue, J.H.; Young, T.M.; Coyle, D.R.; Headlee, W.L.; Bañuelos, G.S.; Hass, A. Ecosystem services of woody crop production systems. BioEnergy Res. 2016, 9, 465–491. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Stanturf, J.A.; Gardiner, E.S.; Bañuelos, G.S.; Hallett, R.A.; Hass, A.; Stange, C.M.; Perdue, J.H.; Young, T.M.; Coyle, D.R.; et al. Environmental technologies of woody crop production systems. BioEnergy Res. 2016, 9, 492–506. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Bauer, E.O. Selecting and utilizing Populus and Salix for landfill covers: Implications for leachate irrigation. Int. J. Phytoremed. 2007, 9, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, J.A.; Zalesny, R.S., Jr.; Wiese, A.H.; Hall, R.B. Choosing tree genotypes for Phytoremedion of landfill leachate using phyto-recurrent selection. Int. J. Phytoremed. 2007, 9, 513–530. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, J.A.; Zalesny, R.S., Jr.; Coyle, D.R.; Hall, R.B. Growth and biomass of Populus irrigated with landfill leachate. For. Ecol. Manag. 2007, 248, 143–152. [Google Scholar] [CrossRef]
- Zalesny, J.A.; Zalesny, R.S., Jr.; Wiese, A.H.; Sexton, B.; Hall, R.B. Sodium and chloride accumulation in leaf, woody, and root tissue of Populus after irrigation with landfill leachate. Environ. Pollut. 2008, 155, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, J.A.; Zalesny, R.S., Jr. Chloride and sodium uptake potential over an entire rotation of Populus irrigated with landfill leachate. Int. J. Phytoremed. 2009, 11, 496–508. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos, G.S.; Dhillon, K.S. Developing a sustainable phytomanagement strategy for excessive selenium in western United States and India. Int. J. Phytoremed. 2011, 13, 208–228. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos, G.S.; Shannon, M.C.; Ajwa, H.; Draper, J.H.; Jordahl, J.; Licht, L. Phytoextraction and accumulation of boron and selenium by poplar (Populus) hybrid clones. Int. J. Phytoremed. 1999, 1, 81–96. [Google Scholar] [CrossRef]
- Shannon, M.C.; Banuelos, G.S.; Draper, J.H.; Ajwa, H.; Jordahl, J.; Licht, L. Tolerance of hybrid poplar (Populus) trees irrigated with varied levels of salt, selenium and boron. Int. J. Phytoremed. 1999, 1, 273–288. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; LeDuc, D.; Johnson, J. Evaluating the tolerance of young hybrid poplar trees to recycled waters high in salinity and boron. Int. J. Phytoremed. 2010, 12, 419–439. [Google Scholar] [CrossRef]
- Mirck, J.; Zalesny, R.S., Jr. Mini-review of knowledge gaps in salt tolerance of plants applied to willows and poplars. Int. J. Phytoremed. 2015, 17, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Fung, L.E.; Wang, S.S.; Altman, A.; Hütterman, A. Effect of NaCl on growth, photosynthesis, ion and water relations of four poplar genotypes. For. Ecol. Manag. 1998, 107, 135–146. [Google Scholar] [CrossRef]
- Chen, S.; Li, J.; Fritz, E.; Wang, S.; Hüttermann, A. Sodium and chloride distribution in roots and transport in three poplar genotypes under increasing NaCl stress. For. Ecol. Manag. 2002, 168, 217–230. [Google Scholar] [CrossRef]
- Chen, S.; Li, J.; Wang, S.; Fritz, E.; Hüttermann, A.; Altman, A. Effects of NaCl on shoot growth, transpiration, ion compartmentation, and transport in regenerated plants of Populus euphratica and Populus tomentosa. Can. J. For. Res. 2003, 33, 967–975. [Google Scholar] [CrossRef]
- Steppuhn, H.; Kort, J.; Wall, K.G. First year growth response of selected hybrid poplar cuttings to root zone salinity. Can. J. Plant Sci. 2008, 88, 473–483. [Google Scholar] [CrossRef]
- Steppuhn, H.; Genuchten, M.T.; Grieve, C.M. Root zone salinity. Crop Sci. 2005, 45, 221–232. [Google Scholar] [CrossRef]
- Smesrud, J.K.; Duvendack, G.D.; Obereiner, J.M.; Jordahl, J.L.; Madison, M.F. Practical salinity management for leachate irrigation to poplar trees. Int. J. Phytoremed. 2012, 14, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Rockwood, D.L.; Naidu, C.V.; Carter, D.R.; Rahmani, M.; Spriggs, T.A.; Lin, C.; Alker, G.R.; Isebrands, J.G.; Segrest, S.A. Short-rotation woody crops and Phytoremedion: Opportunities for agroforestry? Agrofor. Syst. 2004, 61, 51–63. [Google Scholar] [CrossRef]
- Limmer, M.A.; Wilson, J.; Westenberg, D.; Lee, A.; Siegman, M.; Burken, J.G. Phytoremedion removal rates of benzene, toluene, and chlorobenzene. Int. J. Phytoremed. 2018, 20, 666–674. [Google Scholar] [CrossRef]
- Larson, P.R.; Isebrands, J.G. The plastochron index as applied to developmental studies of cottonwood. Can. J. For. Res. 1971, 1, 1–11. [Google Scholar] [CrossRef]
- Headlee, W.L.; Zalesny, R.S., Jr.; Donner, D.M.; Hall, R.B. Using a process-based model (3-PG) to predict and map hybrid poplar biomass productivity in Minnesota and Wisconsin, USA. BioEnergy Res. 2013, 6, 196–210. [Google Scholar] [CrossRef]
- Lazarus, W.; Headlee, W.L.; Zalesny, R.S., Jr. Impacts of supplyshed-level differences in productivity and land costs on the economics of hybrid poplar production in Minnesota, USA. BioEnergy Res. 2015, 8, 231–248. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Bauer, E.O.; Hall, R.B.; Zalesny, J.A.; Kunzman, J.; Rog, C.J.; Riemenschneider, D.E. Clonal variation in survival and growth of hybrid poplar and willow in an in situ trial on soils heavily contaminated with petroleum hydrocarbons. Int. J. Phytoremed. 2005, 7, 177–197. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, R.S., Jr.; Bauer, E.O. Genotypic variability and stability of poplars and willows grown on nitrate-contaminated soils. Int. J. Phytoremed. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, R.S., Jr.; Headlee, W.L.; Gopalakrishnan, G.; Bauer, E.O.; Hall, R.B.; Hazel, D.W.; Isebrands, J.G.; Licht, L.A.; Negri, M.C.; Guthrie-Nichols, E.; et al. Ecosystem services of poplar at long-term Phytoremedion sites in the Midwest and Southeast, United States. WIREs Energy Environ. 2019, in press. [Google Scholar] [CrossRef]
- Minogue, P.J.; Miwa, M.; Rockwood, D.L.; Mackowiak, C.L. Removal of nitrogen and phosphorus by Eucalyptus and Populus at a tertiary treated municipal wastewater sprayfield. Int. J. Phytoremed. 2012, 14, 1010–1023. [Google Scholar] [CrossRef] [PubMed]
- Neuman, D.S.; Wagner, M.; Braatne, J.H.; Howe, J. Part II: Stress physiology—Abiotic. In Biology of Populus and its Implications for Management and Conservation; Stettler, R.F., Bradshaw, H.D., Jr., Heilman, P.E., Hinckley, T.M., Eds.; NRC Research Press, National Research Council of Canada: Ottawa, ON, Canada, 1996; Chapter 17; pp. 423–458. Available online: www.nrcresearchpress.com/doi/book/10.1139/9780660165066#.XKUA8JAUmRs (accessed on 5 April 2019).
- Dickmann, D.I.; Liu, Z.; Nguyen, P.V.; Pregitzer, K.S. Photosynthesis, water relations, and growth of two hybrid Populus genotypes during a severe drought. Can. J. For. Res. 1992, 22, 1094–1106. [Google Scholar] [CrossRef]
- Liu, Z.; Dickmann, D.I. Responses of two hybrid Populus clones to flooding, drought, and nitrogen availability. I. Morphology and growth. Can. J. Bot. 1992, 70, 2265–2270. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Donner, D.M.; Coyle, D.R.; Headlee, W.L. An approach for siting poplar energy production systems to increase productivity and associated ecosystem services. For. Ecol. Manag. 2012, 284, 45–58. [Google Scholar] [CrossRef]
- Zalesny, J.A.; Zalesny, R.S., Jr.; Coyle, D.R.; Hall, R.B.; Bauer, E.O. Clonal variation in morphology of Populus root systems following irrigation with landfill leachate or water during 2 years of establishment. BioEnergy Res. 2009, 2, 134–143. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Bauer, E.O. Evaluation of Populus and Salix continuously irrigated with landfill leachate II. Soils and early tree development. Int. J. Phytoremed. 2007, 9, 307–323. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, R.S., Jr.; Wiese, A.H.; Bauer, E.O.; Riemenschneider, D.E. Ex situ growth and biomass of Populus bioenergy crops irrigated and fertilized with landfill leachate. Biomass Bioenergy 2009, 33, 62–69. [Google Scholar] [CrossRef]
- Ayers, R.S.; Westcot, D.W. Water Quality for Agriculture; FAO Irrigation and Drainage Paper 29 (Revision 1); Food and Agriculture Organization of the United Nations: Rome, Italy, 1985; p. 174. Available online: http://www.fao.org/3/t0234e/t0234e00.htm (accessed on 5 April 2019).
- Arık, F.; Yaldız, T. Heavy metal determination and pollution of the soil and plants of southeast Tavşanlı (Kütahya, Turkey). Clean Soil Air Water 2010, 38, 1017–1030. [Google Scholar] [CrossRef]
- Aksoy, A.; Şahin, U. Elaeagnus angustifolia L. as a biomonitor of heavy metal pollution. Turk. J. Bot. 1999, 23, 83–87. [Google Scholar]
- Burges, A.; Alkorta, I.; Epelde, L.; Garbisu, C. From Phytoremedion of soil contaminants to phytomanagement of ecosystem services in metal contaminated sites. Int. J. Phytoremed. 2018, 20, 384–397. [Google Scholar] [CrossRef]
- Pilipović, A.; Zalesny, R.S., Jr.; Rončević, S.; Nikolić, N.; Orlović, S.; Beljin, J.; Katanić, M. Growth, physiology, and phytoextraction potential of poplar and willow established in soils amended with heavy-metal contaminated, dredged river sediments. J. Environ. Manag. 2019, 239, 352–365. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, R.S., Jr.; Bauer, E.O. Evaluation of Populus and Salix continuously irrigated with landfill leachate I. Genotype-specific elemental Phytoremedion. Int. J. Phytoremed. 2007, 9, 281–306. [Google Scholar] [CrossRef]
- Zalesny, J.A.; Zalesny, R.S., Jr.; Wiese, A.H.; Sexton, B.T.; Hall, R.B. Uptake of macro- and micro-nutrients into leaf, woody, and root tissue of Populus after irrigation with landfill leachate. J. Sustain. For. 2008, 27, 303–327. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salinity Treatment | ||||
|---|---|---|---|---|
| Date | Low | Medium | High | Overall |
| 20 May 2014 | 1.89 ± 0.12 | 4.87 ± 0.07 | 7.73 ± 0.15 | 4.83 ± 0.19 |
| 3 July 2014 | 1.79 ± 0.13 | 4.26 ± 0.12 | 6.31 ± 0.19 | 4.12 ± 0.16 |
| 15 July 2014 | 1.67 ± 0.12 | 3.61 ± 0.13 | 5.87 ± 0.16 | 3.71 ± 0.15 |
| 27 August 2014 | 1.35 ± 0.10 | 3.08 ± 0.11 | 4.72 ± 0.14 | 3.05 ± 0.12 |
| 22 October 2014 | 1.19 ± 0.09 | 2.77 ± 0.09 | 3.89 ± 0.17 | 2.62 ± 0.11 |
| 8 July 2015 | 1.22 ± 0.09 | 2.74 ± 0.11 | 4.02 ± 0.17 | 2.66 ± 0.11 |
| 21 June 2016 | 1.21 ± 0.09 | 2.14 ± 0.11 | 3.46 ± 0.16 | 2.27 ± 0.10 |
| 12 October 2016 | 1.11 ± 0.09 | 1.43 ± 0.11 | 2.53 ± 0.12 | 1.47 ± 0.07 |
| Salinity Treatment | ||||
|---|---|---|---|---|
| Parameter | Low | Medium | High | Overall |
| pH | 7.01 ± 0.23 | 7.68 ± 0.17 | 8.02 ± 0.15 | 7.57 ± 0.14 |
| EC | 0.96 ± 0.28 | 2.98 ± 0.54 | 3.56 ± 0.97 | 2.50 ± 0.45 |
| g kg−1 | ||||
| Calcium (Ca) | 3.56 ± 0.18 | 4.09 ± 0.70 | 5.30 ± 0.53 | 4.32 ± 0.33 |
| Chloride (Cl−) | 0.27 ± 0.04 | 0.76 ± 0.18 | 1.33 ± 0.28 | 0.79 ± 0.15 |
| Magnesium (Mg) | 1.20 ± 0.08 | 1.99 ± 0.18 | 1.89 ± 0.17 | 1.69 ± 0.12 |
| Sodium (Na) | 1.28 ± 0.40 | 3.91 ± 0.39 | 5.86 ± 0.72 | 3.68 ± 0.54 |
| mg kg−1 | ||||
| Aluminum (Al) | 533.07 ± 44.86 | 407.26 ± 70.97 | 256.94 ± 58.99 | 399.09 ± 42.21 |
| Cadmium (Cd) | 0.08 ± 0.03 | 0.06 ± 0.03 | 0.10 ± 0.02 | 0.08 ± 0.01 |
| Cobalt (Co) | 1.69 ± 0.11 | 2.09 ± 0.23 | 1.75 ± 0.20 | 1.84 ± 0.11 |
| Chromium (Cr) | 0.54 ± 0.12 | 0.50 ± 0.08 | 0.52 ± 0.10 | 0.52 ± 0.06 |
| Copper (Cu) | 2.48 ± 0.79 | 2.49 ± 0.58 | 2.77 ± 0.56 | 2.58 ± 0.36 |
| Iron (Fe) | 374.21 ± 19.44 | 368.29 ± 8.83 | 344.41 ± 23.33 | 362.30 ± 10.38 |
| Potassium (K) | 711.86 ± 74.87 | 572.25 ± 22.67 | 474.99 ± 24.69 | 586.37 ± 34.87 |
| Manganese (Mn) | 187.01 ± 21.23 | 227.24 ± 24.82 | 181.12 ± 20.03 | 198.46 ± 12.99 |
| Nickel (Ni) | 22.62 ± 2.08 | 28.06 ± 2.76 | 24.79 ± 2.97 | 25.16 ± 1.53 |
| Lead (Pb) | 2.01 ± 0.60 | 2.33 ± 0.70 | 3.12 ± 0.67 | 2.49 ± 0.37 |
| Zinc (Zn) | 4.83 ± 0.58 | 4.16 ± 0.30 | 3.64 ± 0.23 | 4.21 ± 0.25 |
| Accession a | Clone | Genomic Group b |
|---|---|---|
| 9094432 | Robusta | ‘DN’ P. deltoides × P. nigra |
| 9094423 | DN17 | “ |
| 9094422 | DN182 | “ |
| 9094421 | DN5 | |
| 9094424 | NC14104 | ‘DM’ P. deltoides × P. maximowiczii |
| 9094425 | NC14106 | “ |
| 9094426 | NM2 | ‘NM’ P. nigra × P. maximowiczii |
| 9094427 | NM6 | “ |
| Source of Variation | |||
|---|---|---|---|
| Parameter | T | G | T × G |
| Height | |||
| (2014) First Year | <0.0001 | <0.0001 | 0.2405 |
| (2015) Second Year | <0.0001 | <0.0001 | 0.5738 |
| (2016) Third Year | <0.0001 | <0.0001 | 0.0486 |
| (2017) Fourth Year | <0.0001 | 0.0002 | <0.0001 |
| Leaf Phytoextraction | |||
| Aluminum (Al) | 0.0011 | <0.0001 | 0.6113 |
| Calcium (Ca) | 0.0004 | <0.0001 | 0.3609 |
| Cadmium (Cd) | 0.0827 | <0.0001 | 0.8508 |
| Iron (Fe) | 0.0239 | <0.0001 | 0.2088 |
| Potassium (K) | 0.0247 | 0.8734 | 0.1237 |
| Magnesium (Mg) | <0.0001 | <0.0001 | 0.3731 |
| Manganese (Mn) | 0.0010 | 0.0019 | 0.7696 |
| Sodium (Na) | <0.0001 | <0.0001 | 0.0044 |
| Zinc (Zn) | 0.4731 | 0.0008 | 0.2629 |
| Salinity Treatment | |||||||
|---|---|---|---|---|---|---|---|
| Parameter | Low | Medium | High | Overall | |||
| Height (m) | |||||||
| (2014) First Year | 1.63 ± 0.04 | a * | 1.48 ± 0.04 | b | 1.37 ± 0.04 | c * | 1.50 ± 0.02 |
| (2015) Second Year | 2.03 ± 0.07 | z * | 1.84 ± 0.06 | y | 1.31 ± 0.05 | x * | 1.79 ± 0.04 |
| Leaf phytoextraction (g kg−1) | |||||||
| Calcium (Ca) | 10.41 ± 0.65 | b * | 12.77 ± 0.87 | a | 14.16 ± 0.92 | a * | 12.44 ± 0.50 |
| Magnesium (Mg) | 3.80 ± 0.20 | y * | 5.56 ± 0.38 | z | 5.63 ± 0.31 | z | 5.00 ± 0.20 |
| Leaf phytoextraction (mg kg−1) | |||||||
| Aluminum (Al) | 198.94 ± 20.11 | b * | 278.87 ± 21.09 | a | 271.37 ± 21.13 | a | 249.72 ± 12.51 |
| Iron (Fe) | 67.50 ± 5.30 | yx | 92.36 ± 11.22 | z | 75.21 ± 5.73 | zy | 78.35 ± 4.65 |
| Potassium (K) | 28.12 ± 1.01 | b | 28.53 ± 1.20 | b | 32.79 ± 1.70 | a * | 29.81 ± 0.80 |
| Manganese (Mn) | 53.38 ± 5.13 | y * | 76.55 ± 4.69 | z | 77.25 ± 6.06 | z | 69.06 ± 3.28 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zalesny, R.S., Jr.; Stange, C.M.; Birr, B.A. Survival, Height Growth, and Phytoextraction Potential of Hybrid Poplar and Russian Olive (Elaeagnus Angustifolia L.) Established on Soils Varying in Salinity in North Dakota, USA. Forests 2019, 10, 672. https://doi.org/10.3390/f10080672
Zalesny RS Jr., Stange CM, Birr BA. Survival, Height Growth, and Phytoextraction Potential of Hybrid Poplar and Russian Olive (Elaeagnus Angustifolia L.) Established on Soils Varying in Salinity in North Dakota, USA. Forests. 2019; 10(8):672. https://doi.org/10.3390/f10080672
Chicago/Turabian StyleZalesny, Ronald S., Jr., Craig M. Stange, and Bruce A. Birr. 2019. "Survival, Height Growth, and Phytoextraction Potential of Hybrid Poplar and Russian Olive (Elaeagnus Angustifolia L.) Established on Soils Varying in Salinity in North Dakota, USA" Forests 10, no. 8: 672. https://doi.org/10.3390/f10080672
APA StyleZalesny, R. S., Jr., Stange, C. M., & Birr, B. A. (2019). Survival, Height Growth, and Phytoextraction Potential of Hybrid Poplar and Russian Olive (Elaeagnus Angustifolia L.) Established on Soils Varying in Salinity in North Dakota, USA. Forests, 10(8), 672. https://doi.org/10.3390/f10080672