Monitoring of a Seedling Planting Restoration in a Permanent Preservation Area of the Southeast Atlantic Forest Biome, Brazil


Abstract
:1. Introduction
2. Materials and Methods
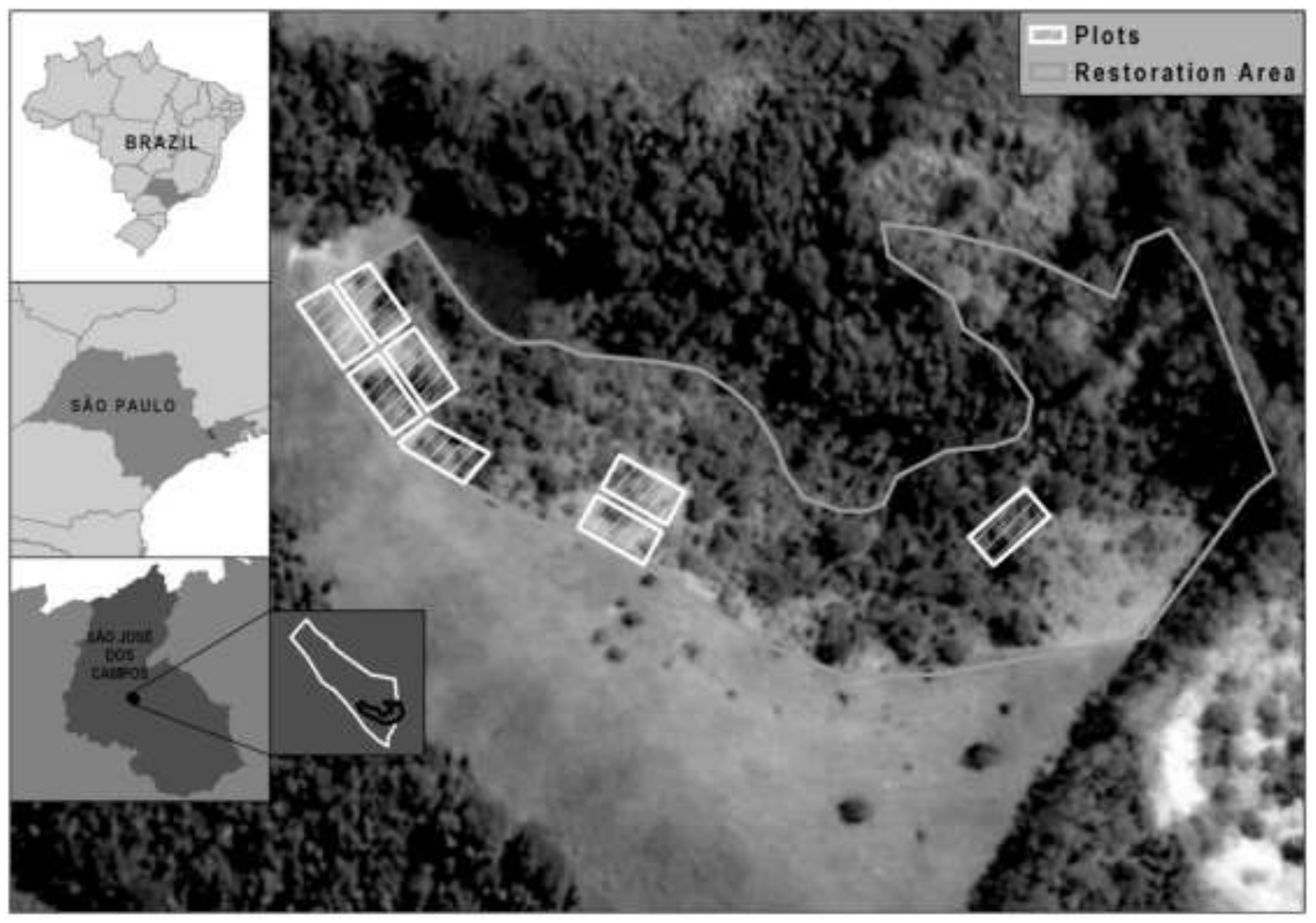
2.1. Study Area
2.2. Plant Measurements
2.3. Statistical Analysis
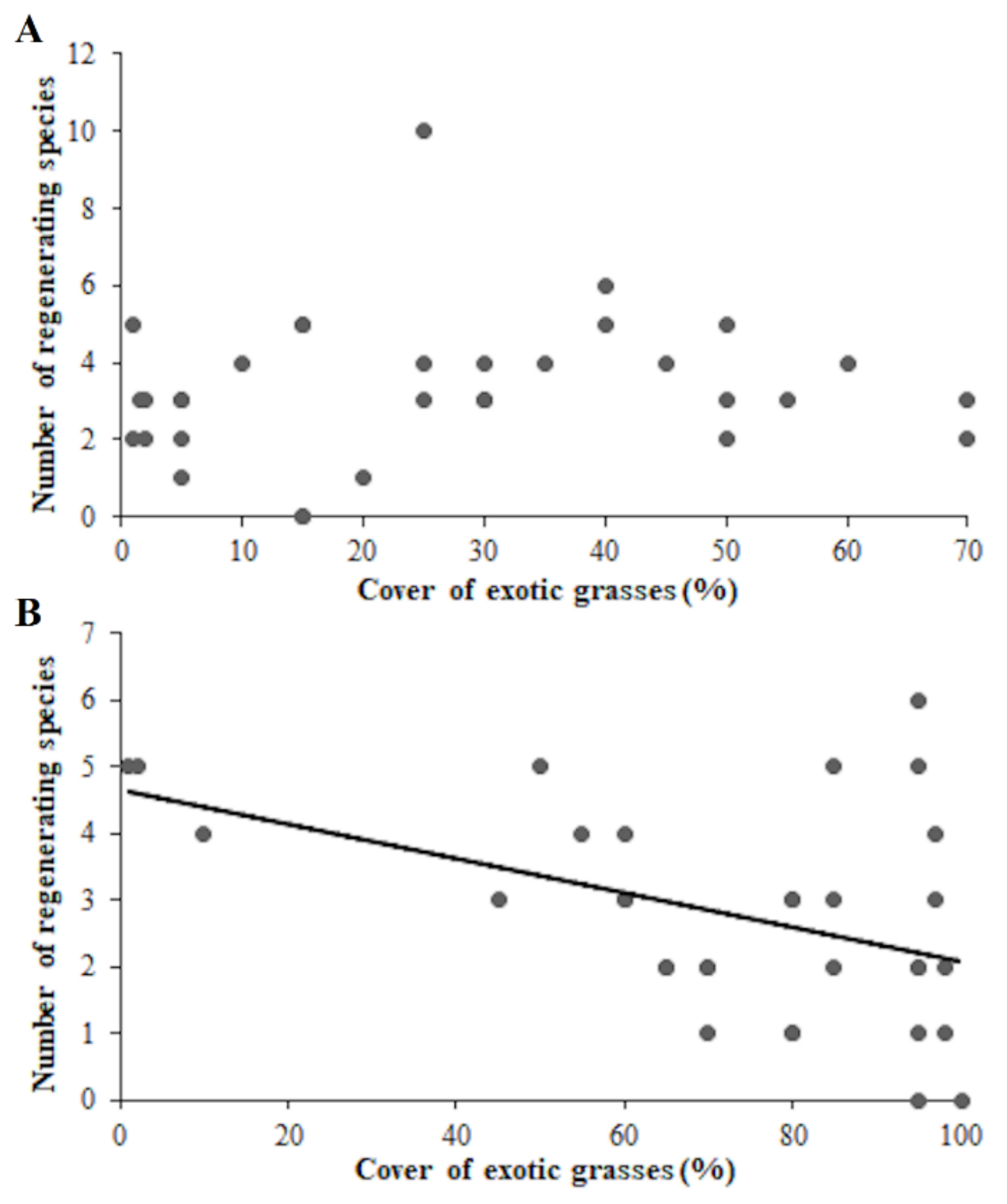
3. Results
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Fundação SOS Mata Atlântica; INPE. Atlas Dos Remanescentes Florestais da Mata Atlântica, Período 2015–2016; Fundação SOS Mata Atlântica e Inpe: São Paulo, Brazil, 2017. [Google Scholar]
- Dean, W. A Ferro e Fogo: A História e a Devastação da Mata Atlântica Brasileira; Companhia das Letras: São Paulo, Brazil, 1996. [Google Scholar]
- Ribeiro, M.C.; Metzger, J.P.; Martensen, A.C.; Ponzoni, F.J.; Hirota, M.M. The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biol. Conserv. 2009, 142, 1141–1153. [Google Scholar] [CrossRef]
- Silva, R.F.B.D.; Batistella, M.; Moran, E.F.; Lu, D. Land changes fostering Atlantic Forest transition in Brazil: Evidences from the Paraíba Valley. Prof. Geographer. 2017, 69, 80–93. [Google Scholar] [CrossRef]
- Brancalion, P.H.S.; Niamir, A.; Broadbent, E.; Crouzeilles, R.; Barros, F.S.M.; Zambrano, A.M.A.; Baccini, A.; Aronson, J.; Goetz, S.; Reid, J.L.; et al. Global restoration opportunities in tropical rainforest landscapes. Sci. Adv. 2019, 5, eaav3223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WRI (World Resources Institute). Restoration Commitments. Initiative 20 × 20. 2018. Available online: https://www.wri.org/our-work/project/initiative-20x20/ restoration-commitments (accessed on 11 July 2019).
- Brasil. Lei no 12.651 of 2012b. Available online: http://www.planalto.gov.br/ccivil_03/_ato2011-2014/2012/lei/L12651compilado.htm (accessed on 10 March 2019).
- Brasil. Decreto no 7.830 of 2012a. Available online: http://www.planalto.gov.br/ccivil_03/_ato2011-2014/2012/decreto/d7830.htm (accessed on 10 March 2019).
- Brasil. Decreto no 8.972 of 2017. Available online: http://www.planalto.gov.br/ccivil_03/_ato2015-2018/2017/decreto/D8972.htm (accessed on 10 March 2019).
- Soares-Filho, B.; Rajão, R.; Macedo, M.; Carneiro, A.; Costa, W.; Coe, M.; Rodrigues, H.; Alencar, A. Cracking Brazil’s Forest Code. Science 2014, 344, 363–364. [Google Scholar] [CrossRef]
- Pinto, S.; Melo, F.; Tabarelli, M.; Padovesi, A.; Mesquita, A.; Scaramuzza, C.A.; Castro, P.; Carrascosa, H.; Calmon, M.; Rodrigues, R.R.; et al. Governing and delivering a biome-wide restoration initiative: The case of Atlantic Forest Restoration Pact in Brazil. Forests 2014, 5, 2212–2229. [Google Scholar] [CrossRef]
- Rodrigues, R.R.; Leitão-Filho, H.F. Matas Ciliares: Conservação e Recuperação; Edusp: São Paulo, Brazil, 2004. [Google Scholar]
- Morrison, E.B.; Lindell, C.A. Active or Passive Forest Restoration? Assessing Restoration Alternatives with Avian Foraging Behavior. Restor. Ecol. 2010, 19, 170–177. [Google Scholar] [CrossRef]
- Norden, N.; Chazdon, R.L.; Chao, A.; Jiang, Y.J.; Vilchez-Alvarado, B. Resilience of tropical rain forests: Tree community reassem- bly in secondary forests. Ecol. Lett. 2009, 12, 385–394. [Google Scholar] [CrossRef]
- Gómez-Pompa, A.; Vazquez-Yanes, C.; Guevara, S. The Tropical Rain Forest: A Nonrenewable Resource. Science 1972, 177, 762–765. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Jaen, M.C.; Aide, T.M. Restoration success: How is it being measured? Restor. Ecol. 2005, 13, 569–577. [Google Scholar] [CrossRef]
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of Degraded Tropical Forest Landscapes. Science 2005, 310, 1628–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brancalion, P.H.S.; Schweizer, D.; Gaudare, U.; Mangueira, J.R.; Lamonato, F.; Farah, F.T.; Nave, A.G.; Rodrigues, R.R. Balancing economic costs and ecological outcomes of passive and active restoration in agricultural landscapes: The case of Brazil. Biotropica 2016, 48, 856–867. [Google Scholar] [CrossRef]
- Rodrigues, R.R.; Brancalion, P.H.S.; Isernhagen, I. Pacto Para a Restauração Ecológica da Mata Atlântica: Referencial dos Conceitos e Ações de Restauração Florestal; Instituto BioAtlântica: São Paulo, Brazil, 2009. [Google Scholar]
- Souza, F.M.; Batista, J.L.F. Restoration of seasonal semideciduous forests in Brazil: Influence of age and restoration design on forest structure. For. Ecol. Manag. 2004, 191, 185–200. [Google Scholar] [CrossRef]
- Rodrigues, R.R.; Lima, R.A.; Gandolfi, S.; Nave, A.G. On the restoration of high diversity forests: 30 years of experience in the Brazilian Atlantic Forest. Biol. Conserv. 2009, 142, 1242–1251. [Google Scholar] [CrossRef]
- Rodrigues, R.R.; Gandolfi, S.; Nave, A.G.; Aronson, J.; Barreto, T.E.; Vidal, C.Y.; Brancalion, P.H. Large-scale ecological restoration of high-diversity tropical forests in SE Brazil. For. Ecol. Manag. 2011, 261, 1605–1613. [Google Scholar] [CrossRef]
- Garcia, L.C.; Hobbs, R.J.; Ribeiro, D.B.; Tamashiro, J.Y.; Santos, F.A.M.; Rodriues, R.R. Restoration over time: Is it possible to restore trees and non-trees in high-diversity for- ests? Appl. Veg. Sci. 2016, 19, 655–666. [Google Scholar] [CrossRef]
- Chaves, R.B.; Durigan, G.; Brancalion, P.H.S.; Aronson, J. On the need of legal frameworks for assessing restoration projects success: New perspectives from So Paulo state (Brazil). Restor. Ecol. 2015, 23, 754–759. [Google Scholar] [CrossRef]
- Pontes, D.M.F.; Engel, V.L.; Parrotta, J.A. Forest Structure, Wood Standing Stock, and Tree Biomass in Diferent Restoration Systems in the Brazilian Atlantic Forest. Forests 2019, 10, 588. [Google Scholar] [CrossRef]
- Rodrigues, R.R.; Lima, R.A.F.; Gandolfi, S.; Brancalion, P.H.S. Restauração Florestal; Oficina de Textos: São Paulo, Brazil, 2015. [Google Scholar]
- Melo, F.P.; Tabarelli, M.; Rodrigues, R.R.; Brancalion, P.H.S. Biodiversity Persistence in Highly Human-Modified Tropical Landscapes Depends on Ecological Restoration. Trop. Conserv. Sci. 2013, 6, 705–710. [Google Scholar] [Green Version]
- Brancalion, P.H.S.; Viani, R.A.G.; Calmon, M.; Carrascosa, H.; Rodrigues, R.R. How to Organize a Large-Scale Ecological Restoration Program? The Framework Developed by the Atlantic Forest Restoration Pact in Brazil. J. Sustain. For. 2013, 32, 728–744. [Google Scholar] [CrossRef]
- Viani, R.A.G.; Barreto, T.E.; Farah, F.T.; Rodrigues, R.R.; Brancalion, P.H.S. Monitoring Young Tropical Forest Restoration Sites: How Much to Measure? Trop. Conserv. Sci. 2018, 11, 1–9. [Google Scholar] [CrossRef]
- Viani, R.A.G.; Holl, K.D.; Padovezi, A.; Strassburg, B.B.N.; Farah, F.T.; Garcia, L.C.; Chaves, R.B.; Rodrigues, R.R.; Brancalion, P.H.S. Protocol for Monitoring Tropical Forest Restoration: Perspectives from the Atlantic Forest Restoration Pact in Brazil. Trop. Conserv. Sci. 2017, 10, 1–8. [Google Scholar] [CrossRef]
- Suganuma, M.S.; Durigan, G. Indicators of restoration success in riparian tropical forests using multiple reference ecosystems. Restor. Ecol. 2015, 23, 238–251. [Google Scholar] [CrossRef]
- Mantoani, M.C.; Torezan, J.M.D. Regeneration response of Brazilian Atlantic Forest woody species to four years of Megathyrsus maximus removal. For. Ecol. Manag. 2016, 359, 141–146. [Google Scholar] [CrossRef]
- Embrapa (Empresa Brasileira de Pesquisa Agropecuária). Súmula da X Reunião Técnica de Levantamento de Solos; Serviço Nacional de Levantamento e Conservação dos Solos: Rio de Janeiro, Brazil, 1979. [Google Scholar]
- Brasil Ministério da Agricultura. Levantamento de Reconhecimento dos Solos do Estado de São Paulo; Serviço Nacional de Pesquisas Agronomicas: Rio de Janeiro, Brazil, 1960.
- Köppen, W. Climatologia: Con un Estudio de los Climas de la Tierra; Fundo de Cultura Econômica: Mexico City, Mexico, 1948. [Google Scholar]
- Urbanetz, C.; Tamashiro, J.Y.; Kinoshita, L.S. Chave de identificação de espécies lenhosas de um trecho de Floresta Ombrófila Densa Atlântica, no Sudeste do Brasil, baseada em caracteres vegetativos. Biota Neotropica 2010, 10, 349–398. [Google Scholar] [CrossRef]
- Eltink, M.; Ramos, E.; Torres, R.B.; Tamashiro, J.Y.; Galembeck, E.; Kimura, E. Chave de identificação de espécies do estrato arbóreo da Mata Atlântica em Ubatuba (SP), com base em caracteres vegetativos. Biota Neotropica 2011, 11, 393–405. [Google Scholar] [CrossRef]
- APG IV (Angiosperm Phylogeny Group IV). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Flora do Brasil 2020 em construção. Jardim Botânico do Rio de Janeiro. Available online: http://www.floradobrasil.jbrj.gov.br (accessed on 03 March 2019).
- São Paulo (Estado). Listagem das Espécies Arbóreas e Indicação de sua Ocorrência Natural nos Biomas/Ecossistemas e Regiões Ecológicas do Estado de São Paulo, com a Classificação Sucessional e a Categoria de Ameaça de Extinção; Secretaria do Meio Ambiente: São Paulo, Brazil, 2008.
- Barbosa, L.M.; Shirasuna, R.T.; Lima, F.C.; De Ortiz, P.R.T. Lista de Espécies Indicadas Para Restauração Ecológica Para Diversas Regiões do Estado de São Paulo. Instituto de Botânica: São Paulo; Instituto de Botânica: São Paulo, Brazil, 2016. Available online: http://botanica.sp.gov.br/files/2016/01/Lista_de_especies_de_SP_CERAD-IBTSMA_2015.pdf (accessed on 10 March 2019).
- Lorenzi, H. Árvores Brasileiras: Manual de Identificação e Cultivo de Plantas Arbóreas Nativas do Brasil; Instituto Plantarum: Nova Odessa, Brazil, 2008. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 03 March 2019).
- Lima, H.C.; Guedes-Bruni, R.R. Serra de Macaé de Cima: Diversidade florística e conservação em Mata Atlântica; Jardim Botânico do Rio de Janeiro: Rio de Janeiro, Brazil, 1997. [Google Scholar]
- Marques, M.C.M.; Silva, S.M.; Liebsch, D. Coastal plain forests in southern and southeastern Brazil: Ecological drivers, floristic patterns and conservation status. Braz. J. Bot. 2015, 38, 1–18. [Google Scholar] [CrossRef]
- Oliveira-Filho, A.T.; Machado, J.N.M. Composição florística de uma floresta semidecídua montana, na Serra de São José, Tiradentes, MG. Acta Bot. Bras. 1993, 7, 71–88. [Google Scholar] [CrossRef]
- Sartori, R.A.; Carvalho, D.A.; Berg, E.V.D.; Marques, J.J.; Dos Santos, R.M. Variações florísticas e estruturais do componente arbóreo de uma floresta estacional semidecidual montana em Socorro, SP. Rodriguésia 2015, 66, 33–49. [Google Scholar] [CrossRef]
- Stopes, C.; Millington, S.; Woodward, L. Dry matter and nitrogen accumulation by three leguminous green manure species and the yield of a following wheat crop in an organic production system. Agric. Ecosyst. Environ. 1996, 57, 189–196. [Google Scholar] [CrossRef]
- Sultani, M.I.; Gill, M.A.; Anwar, M.M.; Athar, M. Evaluation of soil physical properties as influenced by various green manuring legumes and phosphorus fertilization under rain fed conditions. Int. J. Environ. Sci. Technol. 2007, 4, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Engel, V.L.; Parrotta, J.A.; Lamb, D.; Nardoto, G.B.; Ometto, J.P.H.B.; Martinelli, L.A.; Schmidt, S.; Siddique, I.; Siddique, I. Dominance of legume trees alters nutrient relations in mixed species forest restoration plantings within seven years. Biogeochemistry 2008, 88, 89–101. [Google Scholar]
- Howe, H.F.; Smallwood, J. Ecology of Seed Dispersal. Annu. Rev. Ecol. Evol. S 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Da Silva, F.R.; Montoya, D.; Furtado, R.; Memmott, J.; Pizo, M.A.; Rodrigues, R.R. The restoration of tropical seed dispersal networks. Restor. Ecol. 2015, 23, 852–860. [Google Scholar] [CrossRef]
- Carson, W.P.; Schnitzer, S.A. Tropical Forest Community Ecology; Wiley-Blackwell: Oxford, UK, 2008. [Google Scholar]
- Janzen, D.H. Synchronization of sexual reproduction of trees within the dry season in central America. Evolution 1967, 21, 620–637. [Google Scholar] [CrossRef]
- Murphy, P.G.; Lugo, A.E. Ecology of tropical dry forest. Annu. Rev. Ecol. Evol. S 1986, 17, 67–88. [Google Scholar] [CrossRef]
- Derroire, G.; Tigabu, M.; Odén, P.C.; Healey, J.R. The Effects of Established Trees on Woody Regeneration during Secondary Succession in Tropical Dry Forests. Biotropica 2016, 48, 290–300. [Google Scholar] [CrossRef] [Green Version]
- Vieira, D.L.M.; Scariot, A. Principles of Natural Regeneration of Tropical Dry Forests for Restoration. Restor. Ecol. 2006, 14, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Moraes, V.H.F. Periodicidade de crescimento do tronco em árvores da floresta amazônica. Pesqui. Agropecu. Bras. 1970, 5, 315–320. [Google Scholar]
- Melotto, A.; Nicodemo, M.L.; Laura, V.A.; Schleder, D.D.; Pott, A.; Da Silva, V.P.; Bocchese, R.A.; Neto, M.M.G. Sobrevivência e crescimento inicial em campo de espécies florestais nativas do Brasil Central indicadas para sistemas silvipastoris. Revista Árvore 2009, 33, 425–432. [Google Scholar] [CrossRef]
- Bray, E.A. Encyclopedia of Life Sciences; John Wiley & Sons: New York, NY, USA, 2007; pp. 1–7. [Google Scholar]
- Pretzsch, H. Forest Dynamics, Growth and Yield; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–39. [Google Scholar]
- Shimamoto, C.Y.; Botosso, P.C.; Marques, M.C. How much carbon is sequestered during the restoration of tropical forests? Estimates from tree species in the Brazilian Atlantic forest. For. Ecol. Manag. 2014, 329, 1–9. [Google Scholar] [CrossRef]
- Zahran, H.H. Rhizobium-Legume Symbiosis and Nitrogen Fixation under Severe Conditions and in an Arid Climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [PubMed]
- Campoe, O.C.; Stape, J.L.; Mendes, J.C.T. Can intensive management accelerate the restoration of Brazil’s Atlantic forests? For. Ecol. Manag. 2010, 259, 1808–1814. [Google Scholar] [CrossRef]
- D’Antonio, C.M.; Vitousek, P.M. Biological Invasions by Exotic Grasses, the Grass/Fire Cycle, and Global Change. Annu. Rev. Ecol. Syst. 1992, 23, 63–87. [Google Scholar] [CrossRef]
- Leger, E.A.; Espeland, E.K. Coevolution between native and invasive plant competitors: Implications for invasive species management. Evol. Appl. 2010, 3, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Ferez, A.P.C.; Campoe, O.C.; Mendes, J.C.T.; Stape, J.L. Silvicultural opportunities for increasing carbon stock in restoration of Atlantic forests in Brazil. For. Ecol. Manag. 2015, 350, 40–45. [Google Scholar] [CrossRef]
- Elgar, A.T.; Freebody, K.; Pohlman, C.L.; Shoo, L.P.; Catterall, C.P. Overcoming barriers to seedling regeneration during forest restoration on tropical pasture land and the potential value of woody weeds. Front. Plant Sci. 2014, 5, 1–10. [Google Scholar] [CrossRef]
- Brancalion, P.H.S.; Campoe, O.; Mendes, J.C.T.; Noel, C.; Moreira, G.G.; Van Melis, J.; Stape, J.L.; Guillemot, J. Intensive silviculture enhances biomass accumulation and tree diversity recovery in tropical forest restoration. Ecol. Appl. 2019, 29, e01847. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Family | Species | N | EG | SD | LD |
|---|---|---|---|---|---|
| Anacardiaceae | Schinus molle L. | 1 | P | ZOO | E |
| Schinus terebinthifolia Raddi | 4 | P | ZOO | E | |
| Bignoniaceae | Handroanthus chrysotrichus (Mart. ex D.C.) Mattos | 3 | NP | ANE | D |
| Handroanthus impetiginosus (Mart. ex D.C). Mattos | 2 | NP | ANE | D | |
| Handroanthus sp. | 1 | - | ANE | - | |
| Handroanthus umbellatus (Sond.) Mattos | 1 | NP | ANE | D | |
| Connaraceae | Connarus regnellii G. Schellenb. | 2 | NP | ZOO | E |
| Euphorbiaceae | Croton sp. | 1 | - | AUTO | - |
| Fabaceae | Anadenanthera colubrina (Vell.) Brenan | 8 | NP | AUTO | D |
| Dalbergia nigra (Vell.) Allemão ex Benth. | 2 | P | AUTO | D | |
| Inga laurina (Sw.) Willd. | 1 | NP | ZOO | E | |
| Inga marginata Willd. | 1 | NP | ZOO | S | |
| Inga sp. | 7 | - | ZOO | - | |
| Inga vera Willd. subsp. affinis (DC.) T. D. Penn. | 15 | P | ZOO | S | |
| Piptadenia gonoacantha (Mart.) J.F. Macbr. | 5 | P | AUTO | S | |
| Poincianella pluviosa (DC.) L.P. Queiroz | 1 | NP | AUTO | S | |
| Schizolobium parahyba (Vell.) Blake | 10 | P | AUTO | D | |
| Lauraceae | Ocotea sp. | 1 | - | ZOO | - |
| Malpighiaceae | Byrsonima crassifolia (L.) Kunth | 1 | P | ZOO | S |
| Malvaceae | Ceiba speciosa (A.St.-Hil.) Ravenna | 1 | NP | ANE | D |
| Melastomataceae | Tibouchina granulosa (Desr.) Cogn. | 4 | P | ANE | S |
| Myrtaceae | Campomanesia guazumifolia (Cambess.) O. Berg | 1 | P | ZOO | D |
| Eugenia stipitata Mc Vaugh | 1 | NP | ZOO | E | |
| Plinia edulis (Vell.) Sobral | 4 | NP | ZOO | E | |
| Psidium guajava L. | 6 | NP | ZOO | S | |
| Psidium guineense Sw. | 2 | NP | ZOO | E | |
| Primulaceae | Myrsine sp. | 1 | - | ZOO | - |
| Sapindaceae | Cupania vernalis Cambess. | 1 | NP | ZOO | S |
| Urticaceae | Cecropia hololeuca Miq. | 13 | P | ZOO | E |
| Verbenaceae | Citharexylum myrianthum Cham. | 10 | P | ZOO | D |
| Unknown | Unknown 1 | 1 | - | - | - |
| Unknown 2 | 1 | - | - | - | |
| Unknown 3 | 1 | - | - | - | |
| Unknown 4 | 1 | - | - | - | |
| Unknown 5 | 4 | - | - | - | |
| 119 |
| Interval | Parameters | Legumes | Non-Legumes | p-Value | Test |
|---|---|---|---|---|---|
| S1–S2 | DGL (cm) | 1.6 (0.8; 2.5) | 1.1 (0.5; 1.6) | 0.005 | W = 1780.5 |
| H (m) | 0.6 (0.4; 1.0) | 0.4 (0.2; 0.8) | 0.024 | W = 1695.5 | |
| CD (m) | 0.6 (0.1; 0.9) | 0.2 (0; 0.5) | 0.028 | W = 1685.5 | |
| S2–S3 | DGL (cm) | 0.4 (0; 1.0) | 0.3 (0; 0.6) | 0.421 | W = 1416.0 |
| H (m) | 0.1 (0; 0.4) | 0.1 (0; 0.4) | 0.321 | W = 1147.5 | |
| CD (m) | 0.1 (0; 0.7) | 0 (0; 0.1) | 0.122 | W = 1527.0 | |
| DGL (cm) | 2.3 (1.4; 3.3) | 1.6 (0.6; 2.3) | 0.002 | W = 1972.5 | |
| S1–S3 | H (m) | 0.8 (0.4; 1.1) | 0.6 (0. 3;1) | 0.178 | W = 1697.0 |
| CD (m) | 0.7 (0.4; 1.2) | 0.3 (0; 0.5) | <0.001 | W = 2114.0 |
| Interval | Parameters | Pioneers | Non-Pioneers | p-Value | Test |
|---|---|---|---|---|---|
| S1–S2 | DGL (cm) | 1.4 (0.8; 2.2) | 1.2 (0.6; 1.8) | 0.206 | W = 846.5 |
| H (m) | 0.7 (0.3; 1) | 0.5 (0.2; 0.7) | 0.015 | W = 698.0 | |
| CD (m) | 0.4 (0; 0.7) | 0.2 (0; 0.9) | 0.843 | W = 981.0 | |
| S2–S3 | DGL (cm) | 0.5 (0; 0.9) | 0.3 (0; 0.6) | 0.481 | W = 887.0 |
| H (m) | 0.2 (0; 0.4) | 0.1 (0; 0.4) | 0.295 | W = 1102.5 | |
| CD (m) | 0 (0; 0.5) | 0 (0; 0.2) | 0.435 | W = 877.0 | |
| DGL (cm) | 1.9 (1.3; 2.9) | 1.6 (0.6; 2.7) | 0.107 | W = 871.5 | |
| S1–S3 | H (m) | 0.8 (0.4; 1.2) | 0.6 (0.4; 1) | 0.433 | W = 982.5 |
| CD (m) | 0.5 (0.1; 0.9) | 0.4 (0; 0.6) | 0.238 | W = 929.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiore, N.V.; Ferreira, C.C.; Dzedzej, M.; Massi, K.G. Monitoring of a Seedling Planting Restoration in a Permanent Preservation Area of the Southeast Atlantic Forest Biome, Brazil. Forests 2019, 10, 768. https://doi.org/10.3390/f10090768
Fiore NV, Ferreira CC, Dzedzej M, Massi KG. Monitoring of a Seedling Planting Restoration in a Permanent Preservation Area of the Southeast Atlantic Forest Biome, Brazil. Forests. 2019; 10(9):768. https://doi.org/10.3390/f10090768
Chicago/Turabian StyleFiore, Nathalia V., Carolina C. Ferreira, Maíra Dzedzej, and Klécia G. Massi. 2019. "Monitoring of a Seedling Planting Restoration in a Permanent Preservation Area of the Southeast Atlantic Forest Biome, Brazil" Forests 10, no. 9: 768. https://doi.org/10.3390/f10090768
APA StyleFiore, N. V., Ferreira, C. C., Dzedzej, M., & Massi, K. G. (2019). Monitoring of a Seedling Planting Restoration in a Permanent Preservation Area of the Southeast Atlantic Forest Biome, Brazil. Forests, 10(9), 768. https://doi.org/10.3390/f10090768