Biophysical Gradients and Performance of Whitebark Pine Plantings in the Greater Yellowstone Ecosystem


Abstract
:1. Introduction
2. Materials and Methods
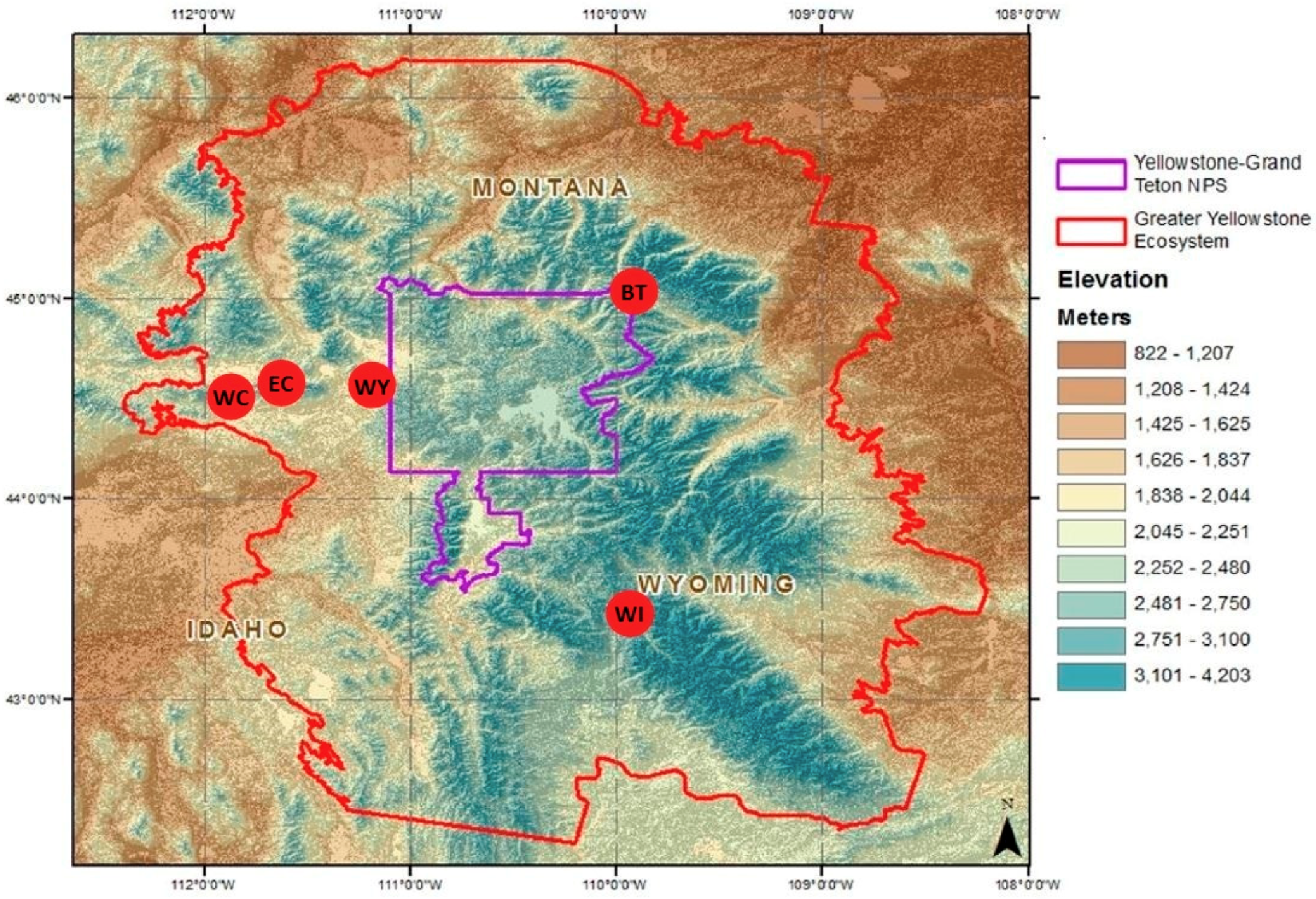
2.1. Study Site
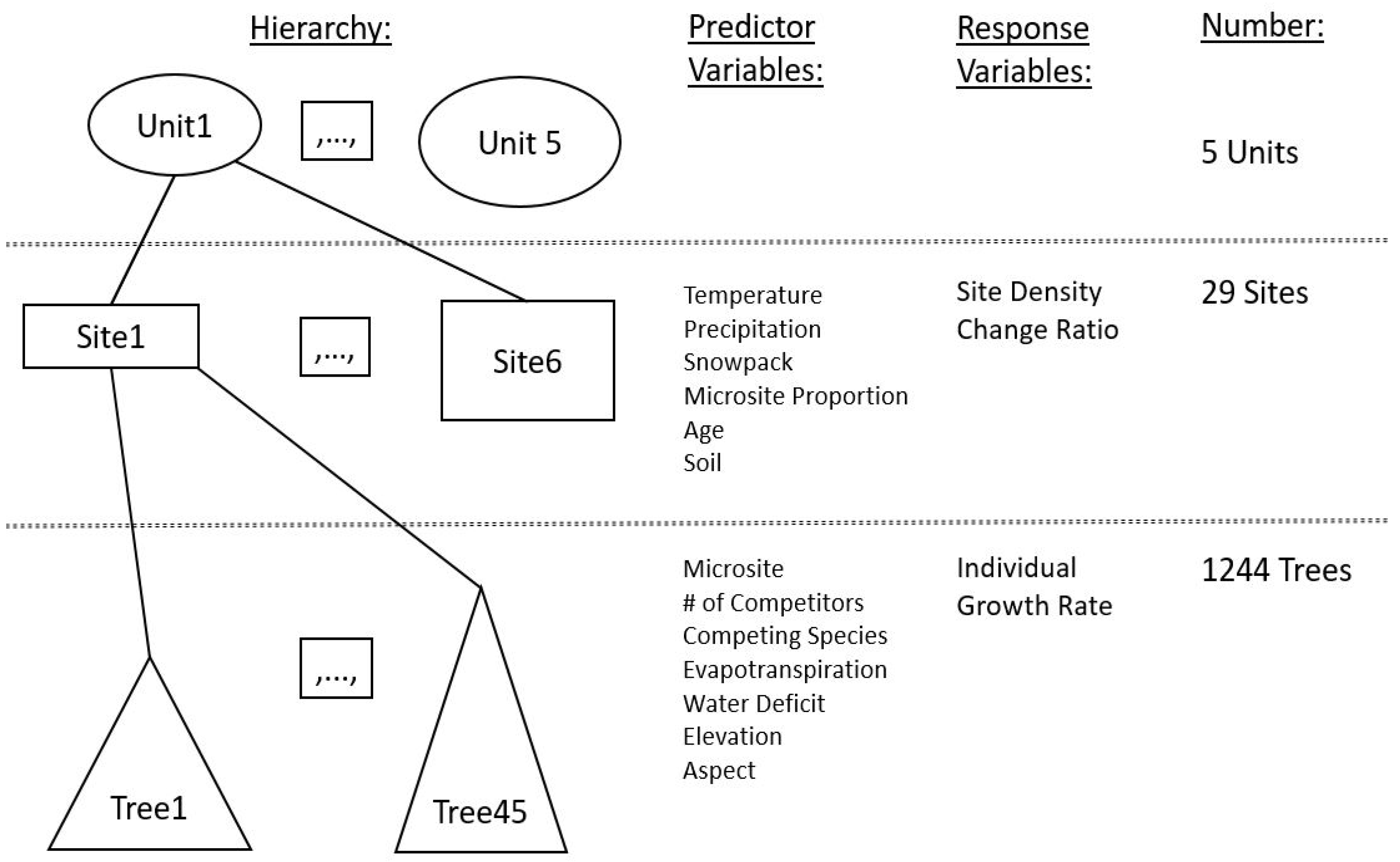
2.2. Sampling Design
2.3. Data
2.3.1. Historical Data
2.3.2. Field Data
2.4. Climate and Water Balance
2.5. Analysis
3. Results
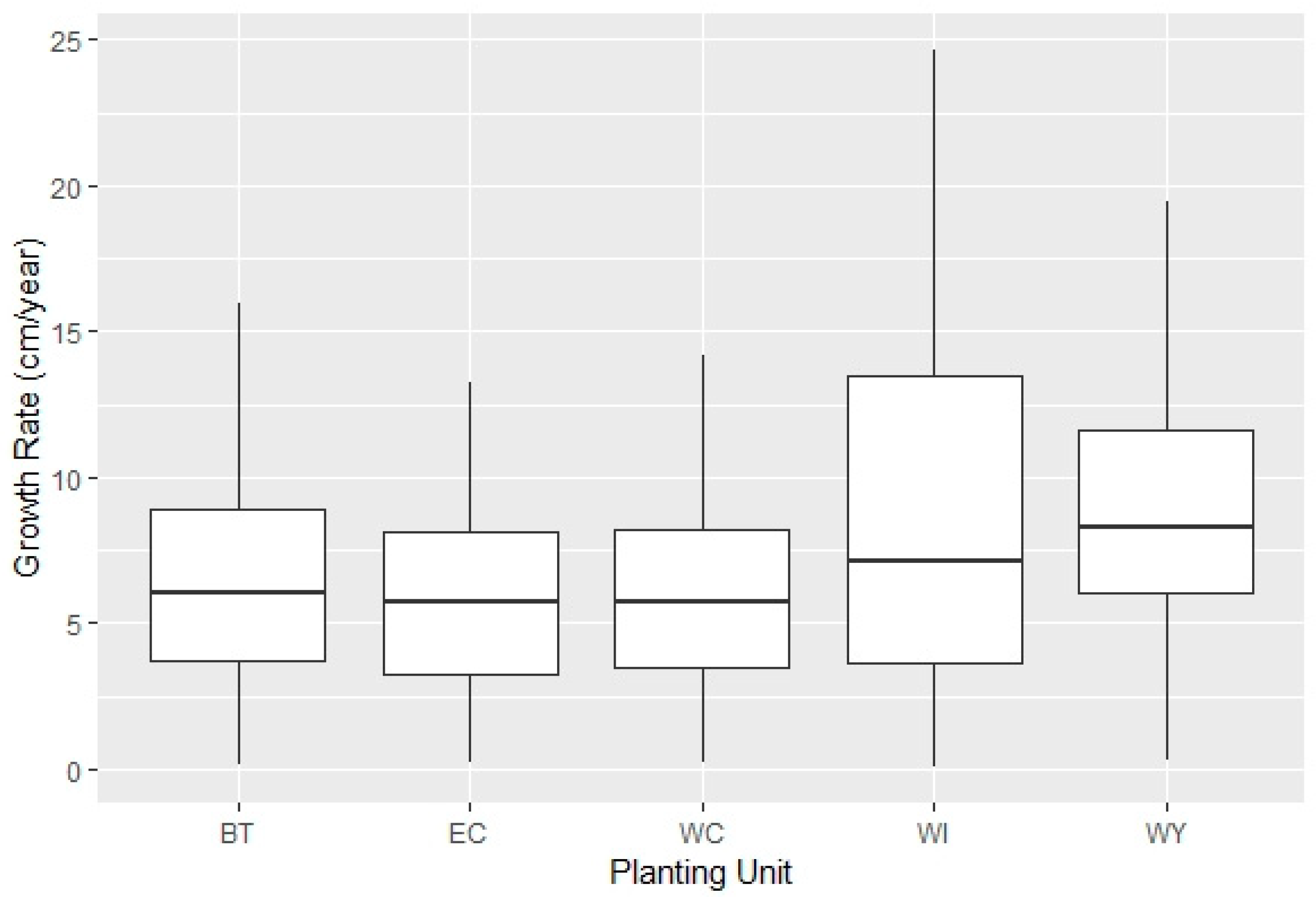
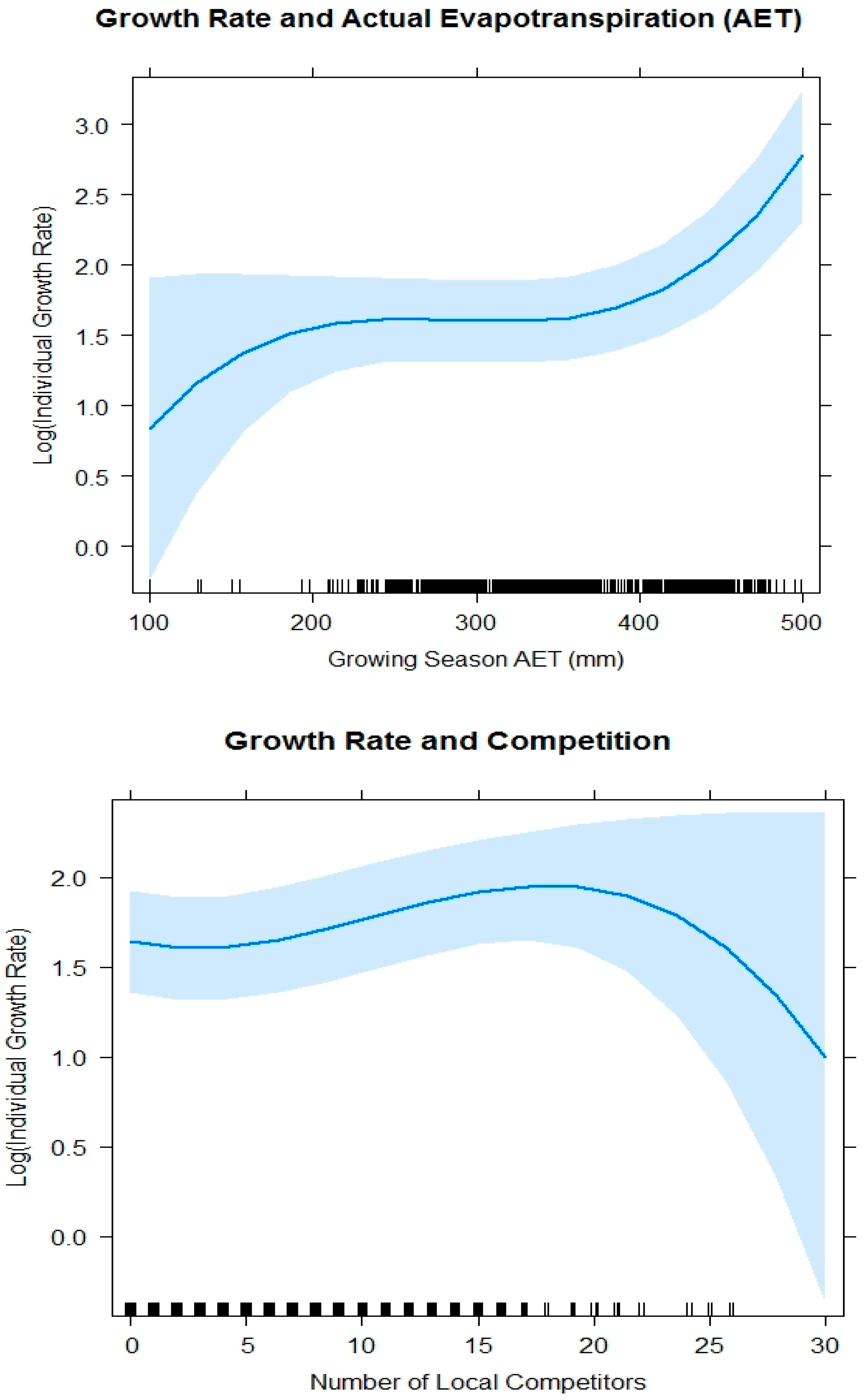
3.1. Individual Growth Rate
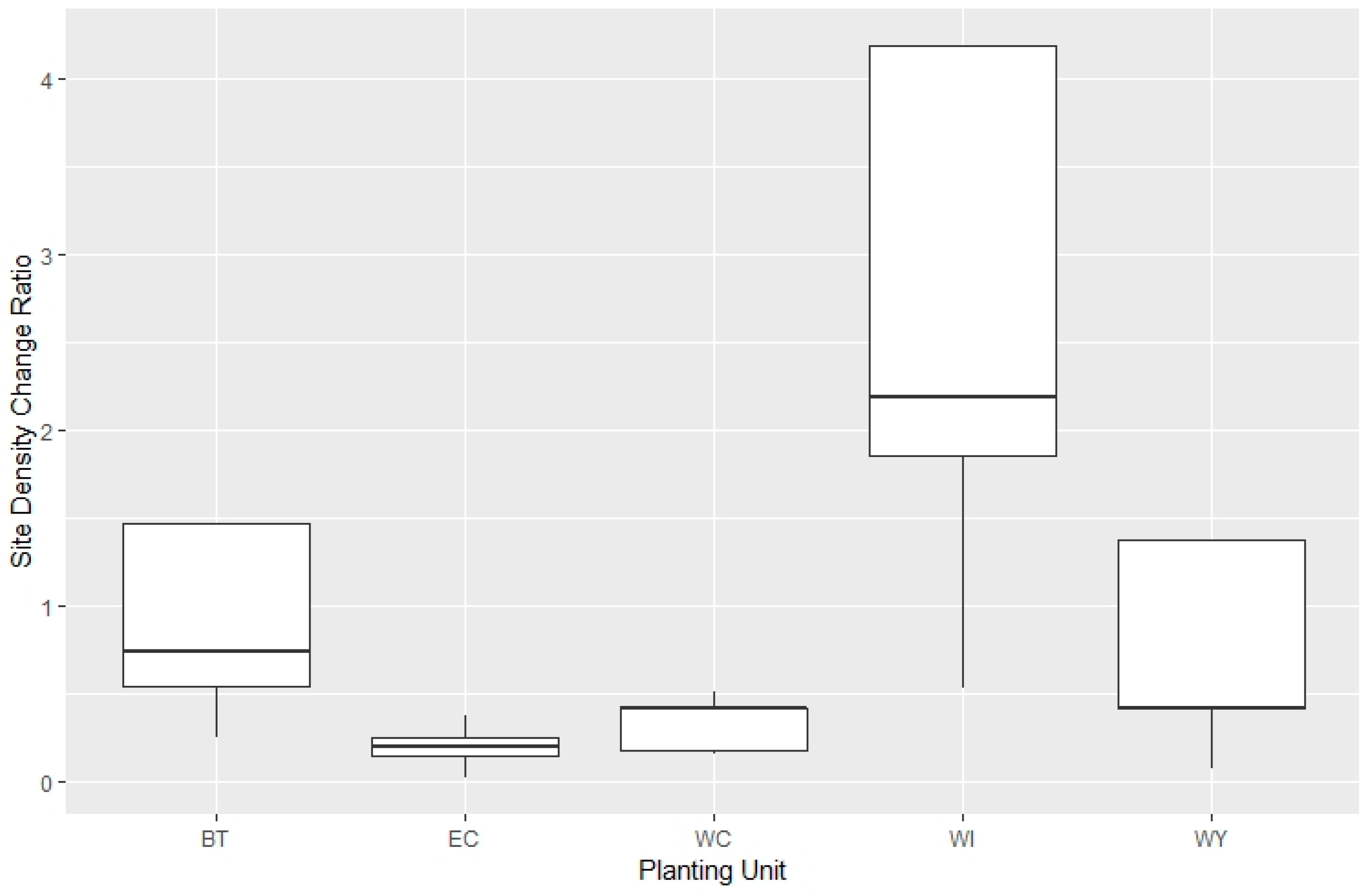
3.2. Site Density Change Ratio
4. Discussion
4.1. Individual Growth Rate Models
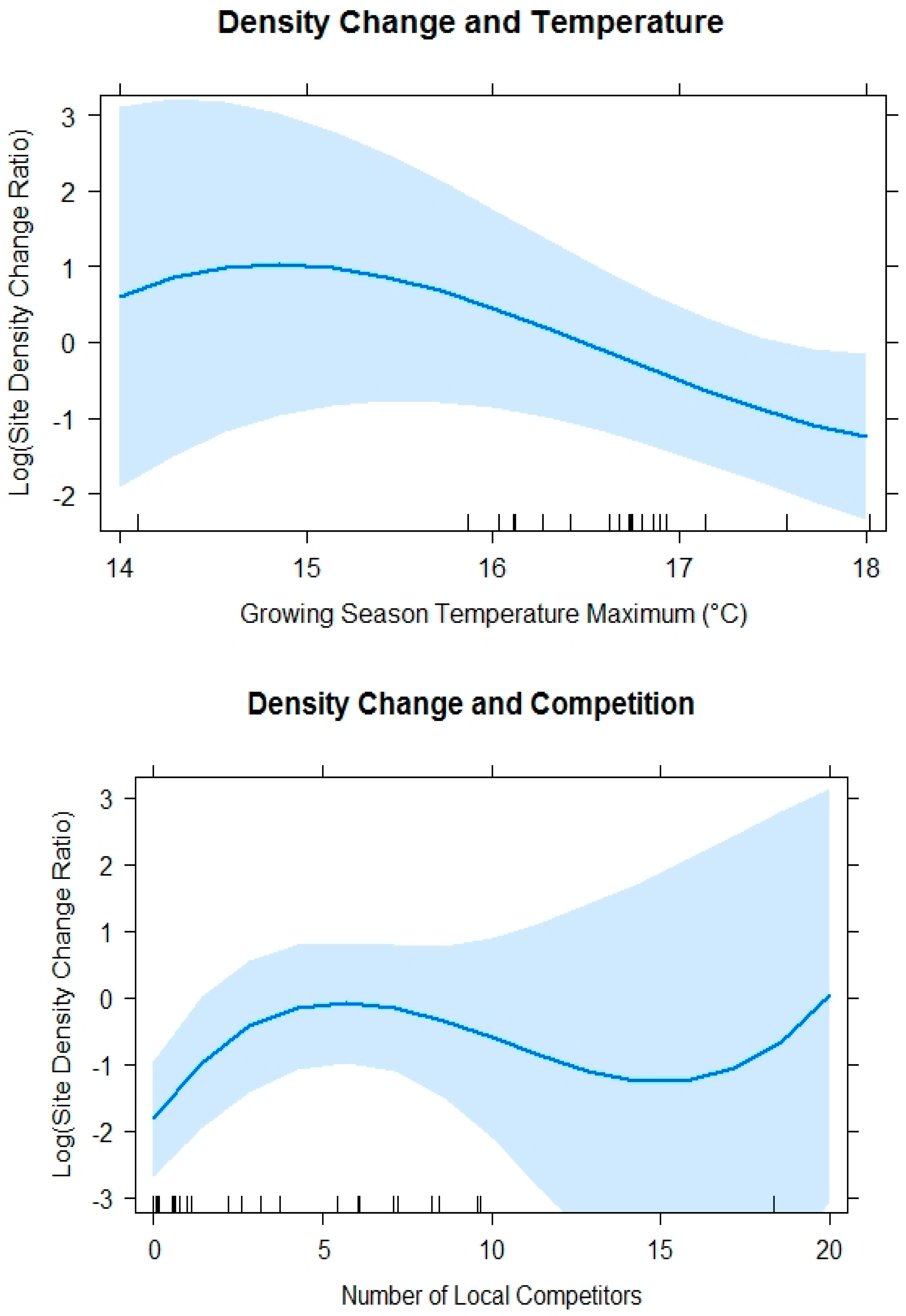
4.2. Site Density Change Ratio Models
4.3. Water Deficit
4.4. Opportunities and Limitations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water (mm) | Soil Type |
|---|---|
| 11.85 | Sand |
| 18.62 | Loamy sand |
| 23.70 | Sandy loam |
| 30.48 | Loam |
| 30.48 | Silt loam |
| 22.01 | Sandy Clay Loam |
| 27.09 | Sandy Clay |
| 27.09 | Clay loam |
| 32.17 | Silty clay loam |
| 40.64 | Silty Clay |
References
- Hansen, A.; Ireland, K.; Legg, K.; Keane, R.; Barge, E.; Jenkins, M.; Pillet, M. Complex Challenges of Maintaining Whitebark Pine in Greater Yellowstone under Climate Change: A Call for Innovative Research, Management, and Policy Approaches. Forests 2016, 7, 54. [Google Scholar] [CrossRef]
- Macfarlane, W.W.; Logan, J.A.; Kern, W.R. An innovative aerial assessment of Greater Yellowstone Ecosystem mountain pine beetle-caused whitebark pine mortality. Ecol. Appl. 2013, 23, 421–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomback, D.; Resler, L.; Keane, R.; Pansing, E.; Andrade, A.; Wagner, A. Community Structure, Biodiversity, and Ecosystem Services in Treeline Whitebark Pine Communities: Potential Impacts from a Non-Native Pathogen. Forests 2016, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Logan, J.A.; Macfarlane, W.W.; Willcox, L. Whitebark pine vulnerability to climate-driven mountain pine beetle disturbance in the Greater Yellowstone Ecosystem. Ecol. Appl. 2010, 20, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Bockino, N.K.; Tinker, D.B. Interactions of White Pine Blister Rust and Mountain Pine Beetle in Whitebark Pine Ecosystems in the Southern Greater Yellowstone Area. Nat. Areas J. 2012, 32, 31–40. [Google Scholar] [CrossRef]
- Jewett, J.T.; Lawrence, R.L.; Marshall, L.A.; Gessler, P.E.; Powell, S.L.; Savage, S.L. Spatiotemporal relationships between climate and whitebark pine mortality in the Greater Yellowstone Ecosystem. For. Sci. 2011, 57, 320–335. [Google Scholar]
- Goeking, S.; Izlar, D. Pinus albicaulis Engelm. (Whitebark Pine) in Mixed-Species Stands throughout Its US Range: Broad-Scale Indicators of Extent and Recent Decline. Forests 2018, 9, 131. [Google Scholar] [CrossRef] [Green Version]
- Costello, C.M.; van Manen, F.T.; Haroldson, M.A.; Ebinger, M.R.; Cain, S.L.; Gunther, K.A.; Bjornlie, D.D. Influence of whitebark pine decline on fall habitat use and movements of grizzly bears in the Greater Yellowstone Ecosystem. Ecol. Evol. 2014, 4, 2004–2018. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service (USFWS). 2011. Available online: http://www.Fws.Gov/mountain-prairie/species/plants/whitebarkpine (accessed on 15 October 2016).
- McCaughey, W.; Scott, G.L.; Izlar, K.L. Whitebark pine planting guidelines. West. J. Appl. For. 2009, 24, 163–166. [Google Scholar] [CrossRef] [Green Version]
- Hansen, A.J.; Phillips, L. Which tree species and biome types are most vulnerable to climate change in the US Northern Rocky Mountains? For. Ecol. Manag. 2015, 338, 68–83. [Google Scholar] [CrossRef]
- Chang, T.; Hansen, A.J.; Piekielek, N. Patterns and Variability of Projected Bioclimatic Habitat for Pinus albicaulis in the Greater Yellowstone Area. PLoS ONE 2014, 9, e111669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkins, D.L.; Swetnam, T.W. A dendroecological assessment of whitebark pine in the Sawtooth–Salmon River region, Idaho. Can. J. For. Res. 1996, 26, 2123–2133. [Google Scholar] [CrossRef] [Green Version]
- Restaino, C.M.; Peterson, D.L.; Littell, J. Increased water deficit decreases Douglas fir growth throughout western US forests. Proc. Natl. Acad. Sci. USA 2016, 113, 9557–9562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, S.T.; Betancourt, J.L.; Booth, R.K.; Gray, S.T. Ecology and the ratchet of events: Climate variability, niche dimensions, and species distributions. Proc. Natl. Acad. Sci. USA 2009, 106, 19685–19692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kueppers, L.M.; Conlisk, E.; Castanha, C.; Moyes, A.B.; Germino, M.J.; de Valpine, P.; Torn, M.S.; Mitton, J.B. Warming and provenance limit tree recruitment across and beyond the elevation range of subalpine forest. Glob. Chang. Biol. 2017, 23, 2383–2395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, C.I.; Westfall, R.D.; Delany, D.L.; Bokach, M.J.; Flint, A.L.; Flint, L.E. Forest mortality in high-elevation whitebark pine (Pinus albicaulis) forests of eastern California, USA; influence of environmental context, bark beetles, climatic water deficit, and warming. Can. J. For. Resh. 2012, 42, 749–765. [Google Scholar] [CrossRef]
- Shanahan, E.; Irvine, K.M.; Thoma, D.; Wilmoth, S.; Ray, A.; Legg, K.; Shovic, H. Whitebark pine mortality related to white pine blister rust, mountain pine beetle outbreak, and water availability. Ecosphere 2016, 7, e01610. [Google Scholar] [CrossRef]
- Stephenson, N. Actual evapotranspiration and deficit: Biologically meaningful correlates of vegetation distribution across spatial scales. J. Biogeogr. 1998, 25, 855–870. [Google Scholar] [CrossRef]
- Buermeyer, K.; Reinhart, D.P.; Legg, K.; Kelly, V. Case study: Whitebark Pine in GYE. In Climate Change in Wildlands: Pioneering Approaches to Science and Management in the Rocky Mountains and Appalachians; Hansen, A.J., Theobald, D.M., Oliff, T., Monihan, W., Eds.; Island Press: Washington, DC, USA, 2016. [Google Scholar]
- Keane, R.E.; Parsons, R.A. Restoring whitebark pine forests of the northern Rocky Mountains, USA. Ecol. Restor. 2010, 28, 56–70. [Google Scholar] [CrossRef]
- Ettinger, A.; HilleRisLambers, J. Competition and facilitation may lead to asymmetric range shift dynamics with climate change. Glob. Chang. Biol. 2017, 23, 3921–3933. [Google Scholar] [CrossRef]
- Loehle, C. Height growth rate tradeoffs determine northern and southern range limits for trees. J. Biogeogr. 1998, 25, 735–742. [Google Scholar] [CrossRef]
- Keane, R.E.; Tomback, D.F.; Aubry, C.A.; Bower, A.D.; Campbell, E.M.; Cripps, C.L.; Jenkins, M.B.; Mahalovich, M.F.; Manning, M.; McKinney, S.T.; et al. A Range-Wide Restoration Strategy for Whitebark Pine (Pinus albicaulis); Gen. Tech. Rep. RMRS-GTR-279; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2012; p. 108.
- Davis, K.; Dobrowski, S.Z.; Holden, Z.A.; Higuera, P.E.; Abatzoglou, J.T. Microclimatic buffering in forests of the future: The role of local water balance. Ecography 2019, 42, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Thornton, P.E.; Thornton, M.M.; Mayer, B.W.; Wei, Y.; Devarakonda, R.; Vose, R.S.; Cook, R.B. Daymet: Daily Surface Weather Data on a 1-km Grid for North America; Version 3; ORNL DAAC: Oak Ridge, TN, USA, 2016.
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration; FAO Irrigation and Drainage Paper N. 56; Food and Agriculture Organization of the United Nations: Rome, Italy, 1998. [Google Scholar]
- Zotarelli, L.; Dukes, M.D.; Romero, C.C.; Migliaccio, K.W. Step by Step Calculation of the Penman-Monteith Evapotranspiration (FAO-56 Method); Document AE459; Florida Cooperative Extension Service, University of Florida: Gainesville, FL, USA, 2013; pp. 1–10. [Google Scholar]
- Lutz, J.A.; van Wagtendonk, J.W.; Franklin, J.F. Climatic water deficit, tree species ranges, and climate change in Yosemite National Park. J. Biogeogr. 2010, 37, 936–950. [Google Scholar] [CrossRef]
- Dilts, T.E.; Weisberg, P.J.; Dencker, C.M.; Chambers, J.C. Functionally relevant climate variables for arid lands: A climatic water deficit approach for modelling desert shrub distributions. J. Biogeogr. 2015, 42, 1986–1997. [Google Scholar] [CrossRef]
- Stephenson, N. Climatic Control of Vegetation Distribution: The Role of the Water Balance. Am. Nat. 1990, 135, 649–670. [Google Scholar] [CrossRef]
- McCune, B.; Keon, D. Equations for potential annual direct incident radiation and heat load. J. Veg. Sci. 2002, 13, 603–606. [Google Scholar] [CrossRef]
- McMaster, G.; Wilhem, W. Growing degree-days: One equation, two interpretations. Agric. For. Meteorol. 1997, 87, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed effects models and extensions in ecology with R. In Statistics for Biology and Health; Springer: New York, NY, USA, 2009; ISBN 978-0-387-87457-9. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Germino, M.J.; Smith, W.K.; Resor, A.C. Conifer seedling distribution and survival in an alpine-treeline ecotone. Plant Ecol. 2002, 162, 157–168. [Google Scholar] [CrossRef]
- Maher, C.T.; Nelson, C.R.; Larson, A.J.; Sala, A. Ecological effects and effectiveness of silvicultural restoration treatments in whitebark pine forests. For. Ecol. Manag. 2018, 429, 534–548. [Google Scholar] [CrossRef]
- Das, A.J.; Stephenson, N.L.; Flint, A.; Das, T.; van Mantgem, P.J. Climatic Correlates of Tree Mortality in Water- and Energy-Limited Forests. PLoS ONE 2013, 8, e69917. [Google Scholar] [CrossRef]
- U.S. Forest Service—Rocky Mountain Research Station. Climate Change Vulnerability and Adaptation in the Intermountain Region [Part 1]; Gen. Tech. Rep. RMRS-GTR-375; Rocky Mountain Research Station: Fort Collins, CO, USA, 2018.
- Stevens-Rumann, C.S.; Kemp, K.B.; Higuera, P.E.; Harvey, B.J.; Rother, M.T.; Donato, D.C.; Morgan, P.; Veblen, T.T. Evidence for declining forest resilience to wildfires under climate change. Ecol. Lett. 2018, 21, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Klutsch, J.G.; Goodrich, B.A.; Jacobi, W.R. Post-fire regeneration dynamics in whitebark pine (Pinus albicaulis) forests in Wind River and Absaroka Mountains, Wyoming, USA. J. For. Res. 2015, 26, 719–733. [Google Scholar] [CrossRef]
- Landenburger, L.; Lawrence, R.; Podruzny, S.; Schwartz, C. Mapping Regional Distribution of a Single Tree Species: Whitebark Pine in the Greater Yellowstone Ecosystem. Sensors 2008, 8, 4983–4994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoylman, Z.H.; Jencso, K.G.; Hu, J.; Holden, Z.A.; Martin, J.T.; Gardner, W.P. The climatic water balance and topography control spatial patterns of atmospheric demand, soil moisture and shallow subsurface flow. Water Resour. Res. 2019, 55, 2370–2389. [Google Scholar] [CrossRef]
- Sepulveda, A.J.; Tercek, M.T.; Al-Chokhachy, R.; Ray, A.M.; Thoma, D.P.; Hossack, B.R.; Pederson, G.T.; Rodman, A.W.; Olliff, T. The shifting climate portfolio of the Greater Yellowstone Area. PLoS ONE 2015, 10, e0145060. [Google Scholar] [CrossRef]
- Jenkins, M.L.; Cripps, C.L.; Gains-Germain, L. Scorched Earth: Suillus colonization of Pinus albicaulis seedlings planted in wildfire-impacted soil affects seedling biomass, foliar nutrient content, and isotope signatures. Plant Soil 2018, 425, 113–131. [Google Scholar] [CrossRef] [Green Version]
- Pansing, E.R.; Tomback, D.F. Survival of whitebark pine seedlings grown from direct seeding: Implications for regeneration and restoration under climate change. Forests 2019, 10, 677. [Google Scholar] [CrossRef] [Green Version]
- Cripps, C.L.; Alger, G.; Sissons, R. Designer niches promote seedling survival in forest restoration: A 7-year study of Whitebark Pine (Pinus albicaulis) seedlings in Waterton Lakes National Park. Forests 2018, 9, 477. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, B.; Jones, B.; Sissons, R.; Cochrane, J.; Park, J.; Smith, C.M.; Stafl, N. Ten years of monitoring illustrates a cascade of effects of white pine blister rust and focuses whitebark pine restoration in the Canadian Rocky and Columbia Mountains. Forests 2018, 9, 138. [Google Scholar] [CrossRef] [Green Version]







| Planting Unit | Number of Planting Sites | Years Planted | Elevation (m) | Latitude (°) | Longitude (°) |
|---|---|---|---|---|---|
| Beartooth | 7 | 1991, 1993 | 2611 | 45.03 | −109.90 |
| East Centennial | 8 | 2010, 2012 | 2352 | 44.53 | −111.61 |
| West Centennial | 6 | 2011 | 2650 | 44.51 | −112.01 |
| West Yellowstone | 4 | 1998–2002 | 2408 | 44.47 | −111.13 |
| Wind River | 4 | 2002, 2013 | 2871 | 43.53 | −109.84 |
| Planting Site | Trees Planted | Hectares Planted | Density (trees/ha) |
|---|---|---|---|
| BT1 | 1700 | 1.59 | 1069 |
| BT2 | 1000 | 0.41 | 2447 |
| BT3 | 980 | 0.88 | 1116 |
| BT4 | 1000 | 1.06 | 943 |
| BT5 | 8335 | 7.86 | 1060 |
| BT6 | 3110 | 2.39 | 1300 |
| BT7 | 1000 | 1.19 | 838 |
| EC1 | 4835 | 11.34 | 426 |
| EC2 | 999 | 2.34 | 426 |
| EC3 | 2896 | 6.79 | 426 |
| EC4 | 3603 | 4.38 | 823 |
| EC5 | 1078 | 1.28 | 840 |
| EC6 | 440 | 0.65 | 680 |
| EC7 | 1839 | 4.31 | 426 |
| EC8 | 3930 | 9.22 | 426 |
| WC1 | 3610 | 7.02 | 514 |
| WC2 | 1390 | 2.70 | 514 |
| WC3 | 2182 | 4.25 | 514 |
| WC4 | 2802 | 5.45 | 514 |
| WC5 | 4833 | 9.40 | 514 |
| WC6 | 6381 | 12.41 | 514 |
| WY1 | 5000 | 4.62 | 1083 |
| WY2 | 1100 | 1.80 | 611 |
| WY3 | 10,000 | 11.18 | 894 |
| WY4 | 2800 | 3.29 | 852 |
| WI1 | 7434 | 12.42 | 598 |
| WI2 | 5101 | 23.18 | 220 |
| WI3 | 3728 | 16.38 | 228 |
| WI4 | 3924 | 16.38 | 2447 |
| Water Balance Variable | Relationship (+/−) with PET |
|---|---|
| Temperature (°C) | + |
| Elevation (m) | − |
| Aspect (°) | +/− (45° is the lowest, 225° is the highest) |
| Shading (%) | − |
| Slope (°) | +/− (minor impact, and dependent upon aspect) |
| Water Balance Variable | Relationship (+/−) with AET |
|---|---|
| Soil Texture (% of sand, silt, clay) | +/− (dependent upon textural triangle) |
| Coarse Fragment (%) | − |
| Temperature (°C) | +/− (dependent upon elevation) |
| Precipitation (mm) | +/− (dependent upon elevation) |
| Predictor Variable | Definition |
|---|---|
| Age | Years since planting |
| Tmean (°C) | Mean annual temperature |
| Tmax (°C) | Maximum monthly temperature |
| PPT (mm) | Mean annual precipitation |
| Snowpack (mm) | Mean spring (March–May) snowpack |
| Rain (mm) | Mean spring (March–May) rain |
| WDannual_mean (mm) | Mean annual water deficit (April–October) |
| WDannual_max (mm) | Maximum annual water deficit (April–October) |
| WDmonth_max (mm) | Maximum monthly water deficit (April–October) |
| PET (mm) | Mean potential evapotranspiration (April–October) |
| AET (mm) | Mean actual evapotranspiration (April–October) |
| GDD | Mean annual growing degree days (April–October) |
| Comp_number | Number of conifers within 3.59 m radius of WBP |
| PIEN | Presence of Pinus engalmanii within 3.59 m radius of WBP |
| ABLA | Presence of Abies lasiocarpa within 3.59 m radius of WBP |
| PICO | Presence of Pinus contorta within 3.59 m radius of WBP |
| Micro | Microsite presence or absence at the individual-level |
| Microprop | Proportion of WBP with a microsite at the site-level |
| Individual Growth Rate Models | AICc | K |
|---|---|---|
| Null Model | ||
| Log(growth_rate) ~ 1 + random (Unit) | 3225.31 | 3 |
| Full Model | ||
| Log(growth_rate) ~ AET + PET + PPT + T + Micro + Comp_number + PICO + PIEN + ABLA + random (Unit) | 3262.92 | 12 |
| Best Model | ||
| Log(growth_rate) ~ AET3 + Comp_number3 + random (Unit) | 3186.57 | 9 |
| Site Density Change Ratio Models | AICc | K |
|---|---|---|
| Null Model | ||
| Log(density_change_ratio) ~ 1 + random (Unit) | 83.72 | 3 |
| Full Model | ||
| Log(density_change_ratio) ~ AET + WDmax_month + Tmax + Microratio + Comp_number + PICO + PIEN + ABLA + random (Unit) | 97.80 | 11 |
| Best Model | ||
| Log(density_change_ratio) ~ Tmax3+ Comp_number3 + random (Unit) | 72.42 | 9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laufenberg, D.; Thoma, D.; Hansen, A.; Hu, J. Biophysical Gradients and Performance of Whitebark Pine Plantings in the Greater Yellowstone Ecosystem. Forests 2020, 11, 119. https://doi.org/10.3390/f11010119
Laufenberg D, Thoma D, Hansen A, Hu J. Biophysical Gradients and Performance of Whitebark Pine Plantings in the Greater Yellowstone Ecosystem. Forests. 2020; 11(1):119. https://doi.org/10.3390/f11010119
Chicago/Turabian StyleLaufenberg, David, David Thoma, Andrew Hansen, and Jia Hu. 2020. "Biophysical Gradients and Performance of Whitebark Pine Plantings in the Greater Yellowstone Ecosystem" Forests 11, no. 1: 119. https://doi.org/10.3390/f11010119
APA StyleLaufenberg, D., Thoma, D., Hansen, A., & Hu, J. (2020). Biophysical Gradients and Performance of Whitebark Pine Plantings in the Greater Yellowstone Ecosystem. Forests, 11(1), 119. https://doi.org/10.3390/f11010119