Effects of Light Intensity and Girdling Treatments on the Production of Female Cones in Japanese Larch (Larix kaempferi (Lamb.) Carr.): Implications for the Management of Seed Orchards

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Girdling Treatments
2.3. Light Intensity
2.4. Tree size and Reproductive Status
2.5. Data Analysis
3. Results
3.1. Status of the L. kaempferi Orchard
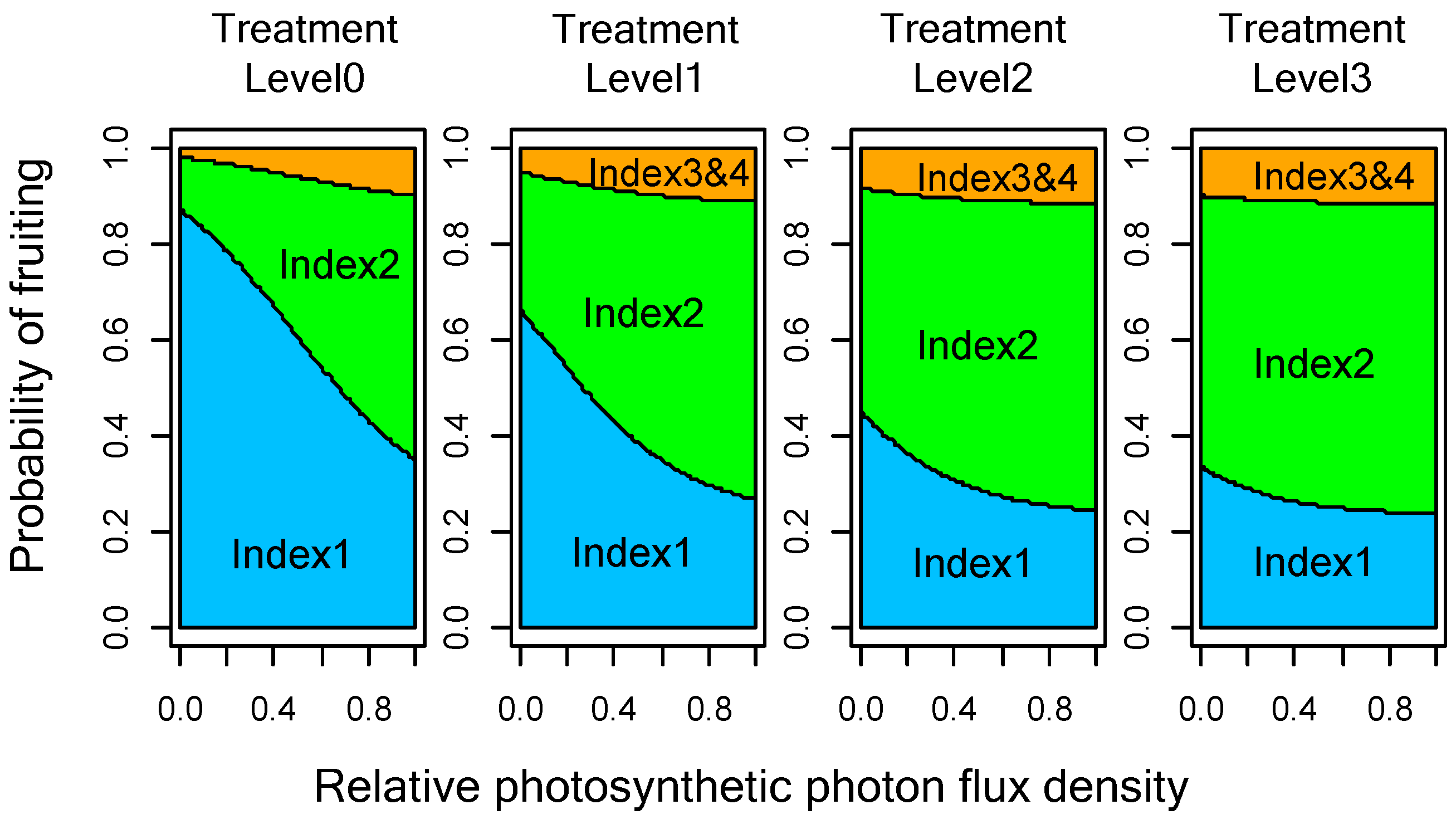
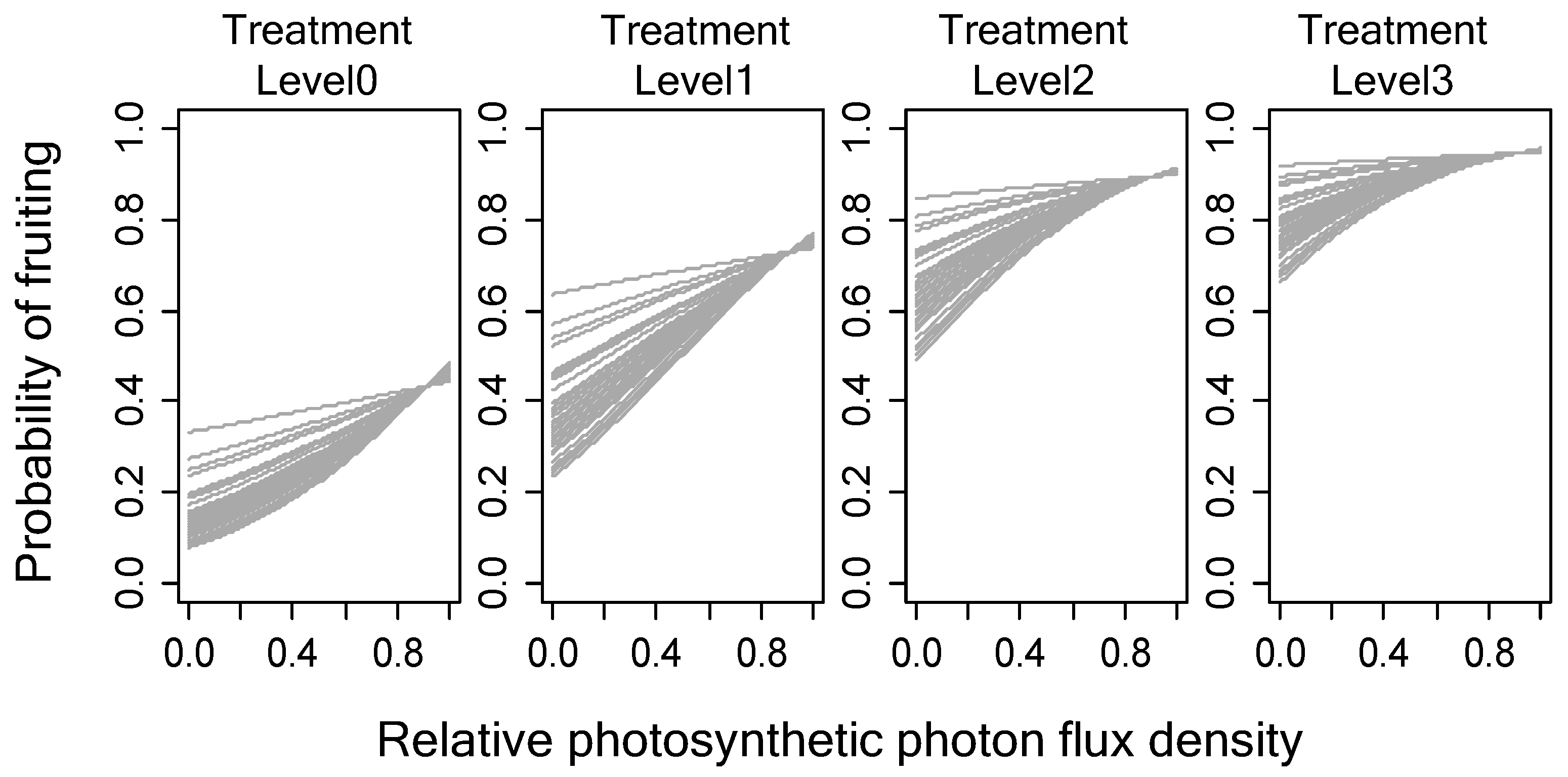
3.2. Relationship between Female Cone Production and Light Intensity or Girdling Treatments
3.3. Relationship between Female Cone Production and Tree Size
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Kurinobu, S. Forest Tree Breeding for Japanese larch. Eurasian J. For. Res. 2005, 8, 127–134. [Google Scholar]
- Park, Y.S.; Fowler, D.P. A provenance test of Japanese larch in eastern Canada, including comparative data on European larch and tamarack. Silvae Genet. 1983, 32, 96–101. [Google Scholar]
- Pâques, L.E. Roles of European and Japanese larch in the genetic control of growth, architecture and wood quality traits in interspecific hybrids (Larix × eurolepis Henry). Ann. For. Sci. 2004, 61, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Baltunis, B.S.; Greenwood, M.S.; Eysteinsson, T. Hybrid vigor in Larix: Growth of intra-and interspecific hybrids of Larix decidua, L. laricina, and L. kaempferi after 5-years. Silvae Genet. 1998, 47, 288–293. [Google Scholar]
- Kita, K.; Fujimoto, T.; Uchiyama, K.; Kuromaru, M.; Akutsu, H. Estimated amount of carbon accumulation of hybrid larch in three 31-year-old progeny test plantations. J. Wood Sci. 2009, 55, 425–434. [Google Scholar] [CrossRef]
- Kita, K.; Sugai, T.; Fujita, S.; Koike, T. Breeding effort on hybrid larch F1 and its responses to environmental stresses. For. Gen. Tree Breed. 2018, 7, 107–114, (In Japanese with English Summary). [Google Scholar]
- Fukatsu, E.; Hiraoka, Y.; Matsunaga, K.; Tsubomura, M.; Nakada, R. Genetic relationship between wood properties and growth traits in Larix kaempferi obtained from a diallel mating test. J. Wood Sci. 2015, 61, 10–18. [Google Scholar] [CrossRef]
- Fukatsu, E.; Tsubomura, M.; Fujisawa, Y.; Nakada, R. Genetic improvement of wood density and radial growth in Larix kaempferi: Results from a diallel mating test. Ann. For. Sci. 2013, 70, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Nagamitsu, T.; Nagasaka, K.; Yoshimaru, H.; Tsumura, Y. Provenance tests for survival and growth of 50-year-old Japanese larch (Larix kaempferi) trees related to climatic conditions in central Japan. Tree Genet. Genomes 2014, 10, 87–99. [Google Scholar] [CrossRef]
- Forestry Agency, Ministry of Agriculture, Forestry and Fisheries of Japan. Annual Report on Forest and Forestry in Japan for FY 2018; Forestry Agency, Ministry of Agriculture, Forestry and Fisheries of Japan: Tokyo, Japan, 2019.
- Forest Tree Breeding Center. The Current States and Statistics in Forest Tree Breeding in Japan; Forest Tree Breeding Center, Forestry and Forestry Product Research Institute: Hitachi, Japan, 2019. [Google Scholar]
- Bonnet-Masimbert, M. Flower induction in conifers: A review of available techniques. For. Ecol. Manag. 1987, 19, 135–146. [Google Scholar] [CrossRef]
- Crain, B.A.; Cregg, B.M. Regulation and management of cone induction in temperate conifers. For. Sci. 2017, 64, 82–101. [Google Scholar] [CrossRef]
- Uchiyama, K.; Kuromaru, M.; Kita, K. Effect of light intensity and girdling on seed production of Larix gmelinii var. japonica clones. Bull. Hokkaido For. Res. Inst. 2007, 44, 119–127, (In Japanese with English Summary). [Google Scholar]
- Asakawa, S.; Fujita, K.; Nagao, A.; Yokoyama, T. The effect of girdling on the coning of larch seed trees as affected by stand density. J. Jpn. For. Soc. 1966, 48, 245–249, (In Japanese with English Summary). [Google Scholar]
- Tamura, A.; Ubukata, M.; Yamada, H.; Fukuda, Y.; Yano, K.; Orita, H. Effect of line thinning on stimulation of flowering in a Japanese larch orchard. Jpn. For. Soc. Cong. 2015, 126, 334. [Google Scholar]
- Shearer, R.C.; Schmidt, W.C. Cone production and stand density in young Larix occidentalis. For. Ecol. Manag. 1987, 19, 219–226. [Google Scholar] [CrossRef]
- Bonnet-Masimbert, M. Effect of growth regulators, girdling, and mulching on flowering of young European and Japanese larches under field conditions. Can. J. For. Res. 1981, 12, 276–279. [Google Scholar] [CrossRef]
- Prill, R. Cone induction on western larch seed trees. B.C. Min. For. Silv. Br. Prog. Rep. 1990, SX87601-10, 29. [Google Scholar]
- Mikami, S.; Asakawa, S.; Iizuka, M.; Yokoyama, T.; Nagao, A.; Takehana, S.; Kaneko, T. Flower induction in Japanese larch, Larix leptolepis Gord. Bull. FFPRI 1979, 307, 9–24, (In Japanese with English summary). [Google Scholar]
- Miller, L.K.; Debell, J. Current seed orchard techniques and innovations. In National Proceedings: Forest and Conservation Nursery Associations 2012; USDA Forest Service: Fort Collins, CO, USA, 2013; pp. 80–86. [Google Scholar]
- Lee, W.Y.; Lee, J.S.; Lee, J.H.; Noh, E.W.; Park, E.-J. Enhanced seed production and metabolic alterations in Larix leptolepis by girdling. For. Ecol. Manag. 2011, 261, 1957–1961. [Google Scholar] [CrossRef]
- Markiewicz, P. Problems with seed production of European larch in seed orchards in Poland. In Proceedings of a Seed Orchard Conference; Lindgren, D., Ed.; Swedish University of Agricultural Sciences: Umeå, Sweden, 2007; pp. 161–164. [Google Scholar]
- Philipson, J.J. Effects of cultural treatments and gibberellin A4/17 on flowering of container-grown European and Japanese larch. Can. J. For. Res. 1995, 25, 184–192. [Google Scholar] [CrossRef]
- Colas, F.; Perron, M.; Tousignant, D.; Parent, C.; Pelletier, M.; Lemay, P. A novel approach for the operational production of hybrid larch seeds under northern climatic conditions. For. Chron. 2008, 84, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Verkaik, I.; Espelta, J.M. Post-fire regeneration thinning, cone production, serotiny and regeneration age in Pinus halepensis. For. Ecol. Manag. 2006, 231, 155–163. [Google Scholar] [CrossRef]
- Peters, G.; Sala, A. Reproductive output of ponderosa pine in response to thinning and prescribed burning in western Montana. Can. J. For. Res. 2008, 38, 844–850. [Google Scholar] [CrossRef]
- Matsushita, M.; Nakagawa, M.; Tomaru, N. Sexual differences in year-to-year flowering trends in the dioecious multi-stemmed shrub Lindera triloba: Effects of light and clonal integration. J. Ecol. 2011, 99, 1520–1530. [Google Scholar] [CrossRef]
- Lindh, B.C. Flowering of understory herbs following thinning in the western Cascades, Oregon. For. Ecol. Manag. 2008, 256, 929–936. [Google Scholar] [CrossRef]
- Matsushita, M.; Setsuko, S.; Tamaki, I.; Nakagawa, M.; Nishimura, N.; Tomaru, N. Thinning operations increase the demographic performance of the rare subtree species Magnolia stellata in a suburban forest landscape. Landsc. Ecol. Eng. 2016, 12, 179–186. [Google Scholar] [CrossRef]
- Setsuko, S.; Tamaki, I.; Ishida, K.; Tomaru, N. Relationships between flowering phenology and female reproductive success in the Japanese tree species Magnolia stellata. Botany 2008, 86, 248–258. [Google Scholar] [CrossRef]
- Levin, A.G.; Lavee, S. The influence of girdling on flower type, number, inflorescence density, fruit set, and yields in three different olive cultivars (Barnea, Picual, and Souri). Austral J. Agric. Res. 2005, 56, 827–831. [Google Scholar] [CrossRef]
- Brar, H.S.; Singh, Z.; Swinny, E.; Cameron, I. Girdling and grapevine leafroll associated viruses affect berry weight, colour development and accumulation of anthocyanins in ‘Crimson Seedless’ grapes during maturation and ripening. Plant Sci. 2008, 175, 885–897. [Google Scholar] [CrossRef]
- Rivas, F.; Gravina, A.; Agustí, M. Girdling effects on fruit set and quantum yield efficiency of PSII in two Citrus cultivars. Tree Physiol. 2007, 27, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Van Kleunen, M.; Stuefer, J.F. Quantifying the effects of reciprocal assimilate and water translocation in a clonal plant by the use of steam girdling. Oikos 1999, 85, 135–145. [Google Scholar] [CrossRef]
- Isogimi, T.; Matsushita, M.; Watanabe, Y.; Nakagawa, M. Sexual differences in physiological integration in the dioecious shrub Lindera triloba: A field experiment using girdling manipulation. Ann. Bot. 2011, 107, 1029–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isogimi, T.; Matsushita, M.; Nakagawa, M. Species-specific sprouting pattern in two dioecious Lindera shrubs: The role of physiological integration. Flora 2014, 209, 718–724. [Google Scholar] [CrossRef]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2016. [Google Scholar]
- Ross, S.D. Promotion of Flowering in Western Larch by Girdling and Gibberellin A4/7, and Recommendations for Selection and Treatment of Seed Trees; BC Ministry of Forests, Research Laboratory: Victoria, BS, Canada, 1991; p. 13.
- Longman, K.A.; Nasr, T.A.; Wareing, P.F. Gravimorphism in trees. 4. The effect of gravity on flowering. Ann. Bot. 1965, 29, 105–111. [Google Scholar] [CrossRef]
- Hashizume, H. Studies on flower bud formation, flower sex differentiation and their control in conifers. Bull. Tottori Univ. For. 1973, 7, 1–139, (In Japanese with English Summary). [Google Scholar]
- Katsuta, M.; Saito, M.; Yamamoto, C.; Kaneko, T.; Itoo, M. Effect of gibberellins on the promotion of strobilus production in Larix leptolepis Gord. and Abies hornolepis Sieb. and Zucc. Bull. FFPRI 1981, 313, 37–45, (In Japanese with English Summary). [Google Scholar]
- Philipson, J.J. Effects of girdling and gibberellin A4/17 on flowering of European and Japanese larch grafts in an outdoor clone bank. Can. J. For. Res. 1996, 26, 355–359. [Google Scholar] [CrossRef]
- Wheeler, N.C.; Cade, S.C.; Masters, C.J.; Ross, S.D.; Keeley, J.W.; Hsin, L.Y. Girdling: A safe, effective and practical treatment for enhancing seed yields in Douglas-fir seed orchards. Can. J. For. Res. 1985, 15, 505–510. [Google Scholar] [CrossRef]
- Graham, R.T. Effect of nitrogen fertilizer and girdling on cone and seed production of western larch. In Proc. Conifer Tree Seed in the Inland Mountain West Symposium General Technical Report; Shearer, R.C., Ed.; USDA Forest Service Intermountain Research Station: Ogden, UT, USA, 1986; pp. 166–170. [Google Scholar]
- Owens, J.N.; Blake, M.D. Forest seed tree production. In Information Report PI-X-53; Petawawa National Forestry Institute, Canada Forest Service: Chalk River, ON, Canada, 1985; p. 161. [Google Scholar]
- Chałupka, W.; Giertych, M.; Kopcewicz, J. Effect of polyethylene covers on the flowering of Norway spruce (Picea abies (L.) Karst.) grafts. Physiol. Plant. 1982, 54, 79–81. [Google Scholar] [CrossRef]
- Matthews, J.D. Factors affecting the production of seed by forest trees. For. Abstr. 1963, 24, 1–13. [Google Scholar]
- Despland, E.; Houle, G. Aspect influences cone abundance within the crown of Pinus banksiana Lamb. trees at the limit of the species distribution in northern Quebec (Canada). Écoscience 1997, 4, 521–525. [Google Scholar] [CrossRef]
- Funda, T.; El-Kassaby, Y.A. Seed orchard genetics. Cab. Rev. 2012, 7, 1–23. [Google Scholar] [CrossRef]
- Chałupka, W. Do we need flower stimulation in seed orchards? In Proceedings of a Seed Orchard Conference; Lindgren, D., Ed.; Swedish University of Agricultural Sciences: Umeå, Sweden, 2007; pp. 37–42. [Google Scholar]
- Funda, T. Population Genetics of Conifer Seed Orchards. Ph.D. Thesis, University of British Columbia, Vancouver, BS, Canada, 2012. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coef. | S.E. | Z-Value | p-Value | |
|---|---|---|---|---|
| Intercept | −1.84 | 0.45 | −4.05 | <0.001 |
| Girdling level 1 | 1.25 | 0.78 | 1.61 | 0.109 |
| Girdling level 2 | 2.39 | 0.71 | 3.39 | <0.001 |
| Girdling level 3 | 3.09 | 1.12 | 2.77 | 0.006 |
| rPPFD | 1.73 | 0.79 | 2.20 | 0.028 |
| rPPFD: 0.2 | rPPFD: 0.5 | rPPFD: 0.8 | |
|---|---|---|---|
| Girdling level 0 | 26.4% | 13.1% | 2.9% |
| Girdling level 1 | 16.8% | 7.4% | 1.4% |
| Girdling level 2 | 8.3% | 3.3% | 0.6% |
| Girdling level 3 | 4.8% | 1.8% | 0.3% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsushita, M.; Nishikawa, H.; Tamura, A.; Takahashi, M. Effects of Light Intensity and Girdling Treatments on the Production of Female Cones in Japanese Larch (Larix kaempferi (Lamb.) Carr.): Implications for the Management of Seed Orchards. Forests 2020, 11, 1110. https://doi.org/10.3390/f11101110
Matsushita M, Nishikawa H, Tamura A, Takahashi M. Effects of Light Intensity and Girdling Treatments on the Production of Female Cones in Japanese Larch (Larix kaempferi (Lamb.) Carr.): Implications for the Management of Seed Orchards. Forests. 2020; 11(10):1110. https://doi.org/10.3390/f11101110
Chicago/Turabian StyleMatsushita, Michinari, Hiroki Nishikawa, Akira Tamura, and Makoto Takahashi. 2020. "Effects of Light Intensity and Girdling Treatments on the Production of Female Cones in Japanese Larch (Larix kaempferi (Lamb.) Carr.): Implications for the Management of Seed Orchards" Forests 11, no. 10: 1110. https://doi.org/10.3390/f11101110
APA StyleMatsushita, M., Nishikawa, H., Tamura, A., & Takahashi, M. (2020). Effects of Light Intensity and Girdling Treatments on the Production of Female Cones in Japanese Larch (Larix kaempferi (Lamb.) Carr.): Implications for the Management of Seed Orchards. Forests, 11(10), 1110. https://doi.org/10.3390/f11101110