Dwarf Mistletoe Infection Interacts with Tree Growth Rate to Produce Opposing Direct and Indirect Effects on Resin Duct Defenses in Lodgepole Pine


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
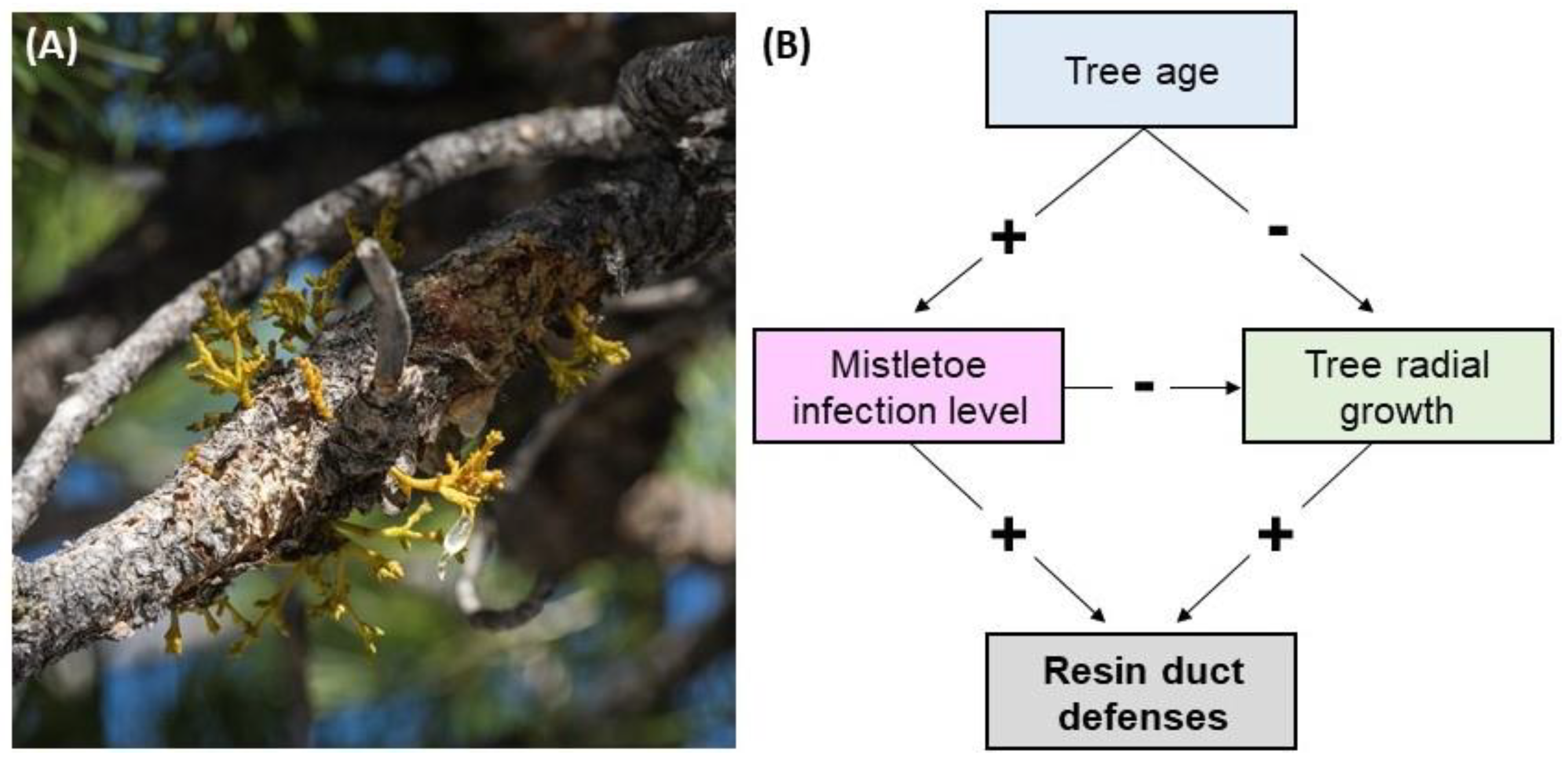
:1. Introduction
2. Materials and Methods
2.1. Study Site and Data Collection
2.2. Data Analyses
3. Results and Discussion
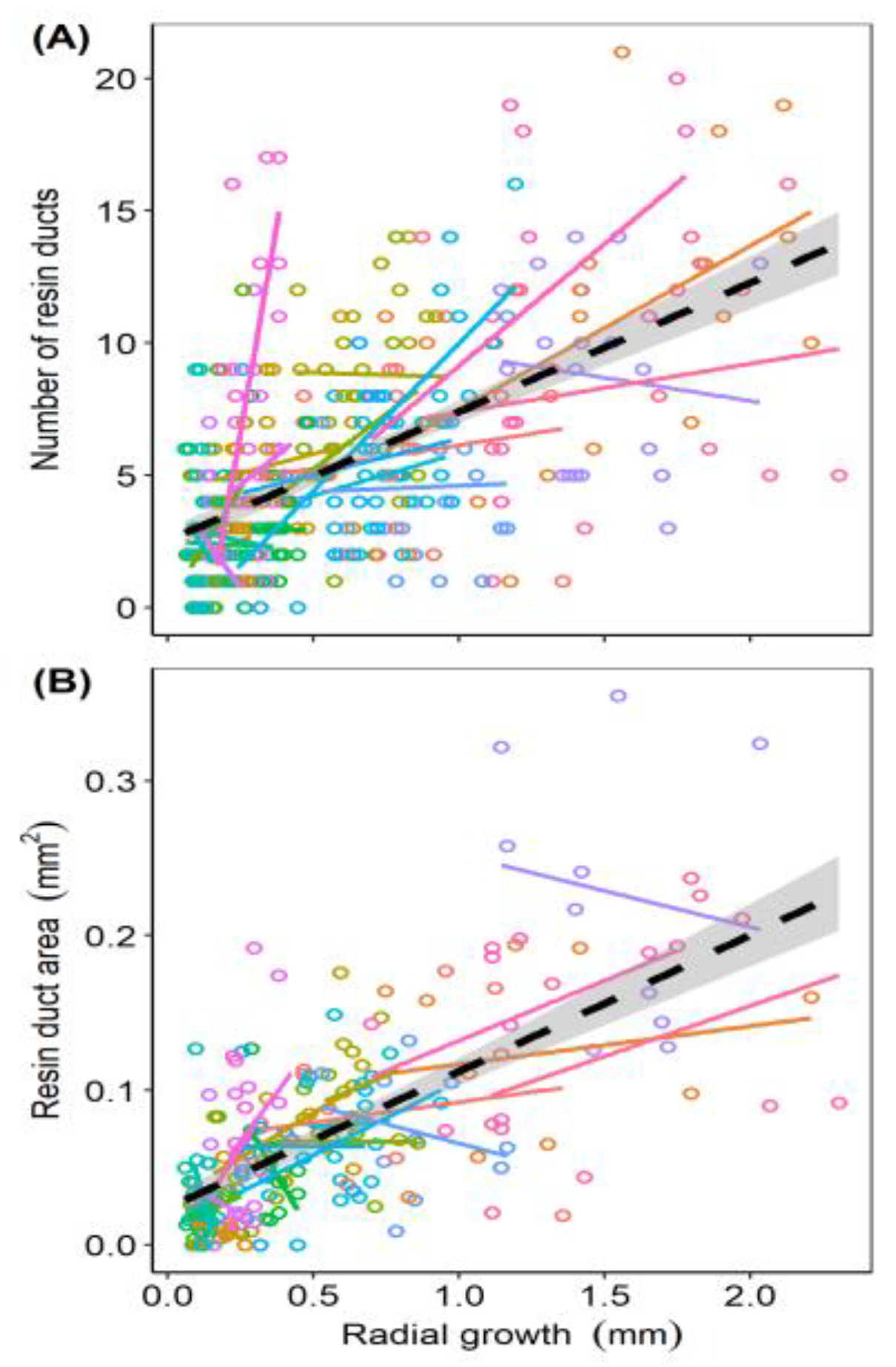
3.1. Relationship of Number and Area of Resin Ducts
3.2. Prediction 1: Resin Duct Defenses and Tree Growth
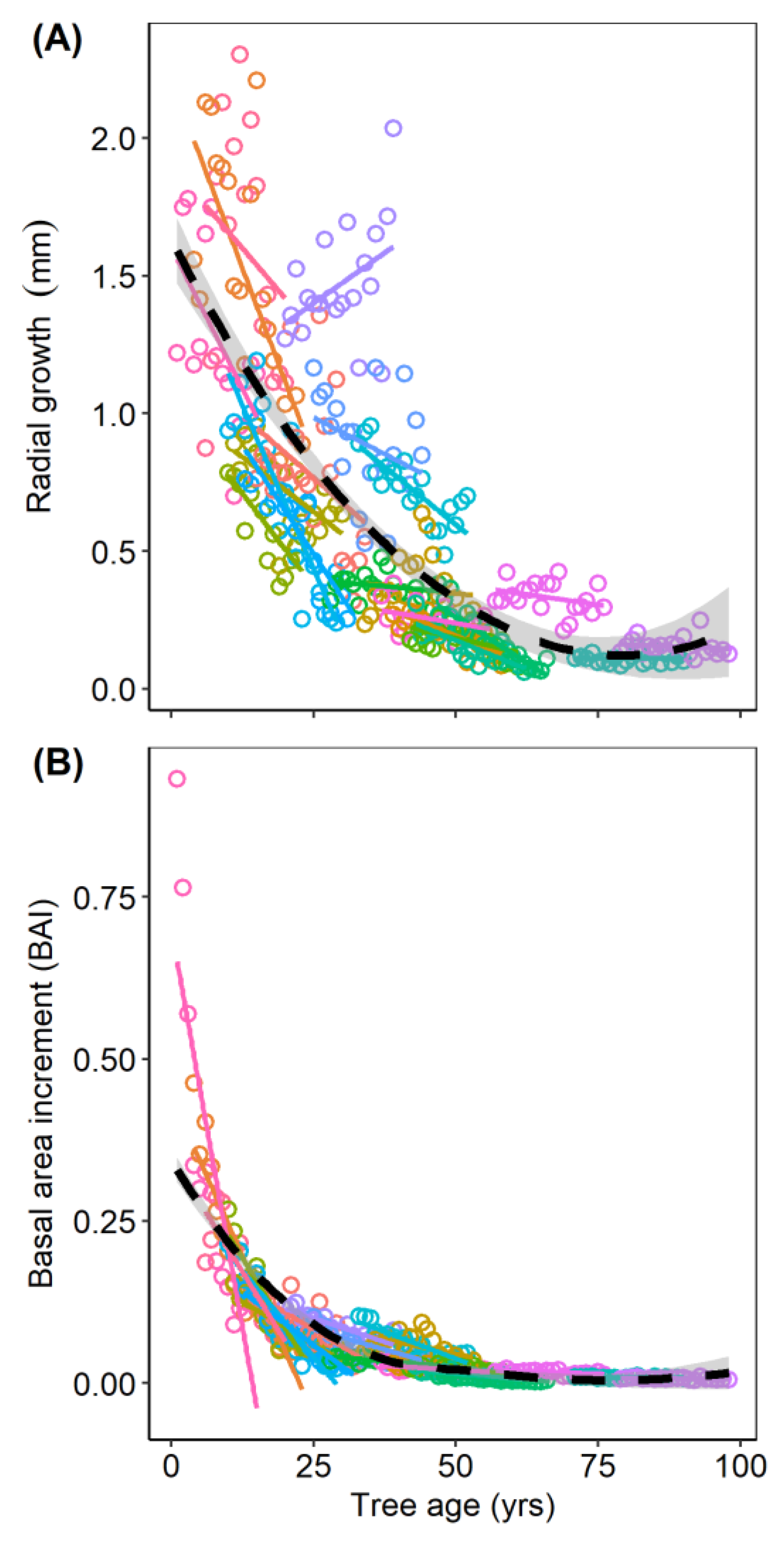
3.3. Prediction 2: Influence of Tree Age on Growth
3.4. Prediction 3: Influence of Tree Age on Mistletoe Infection
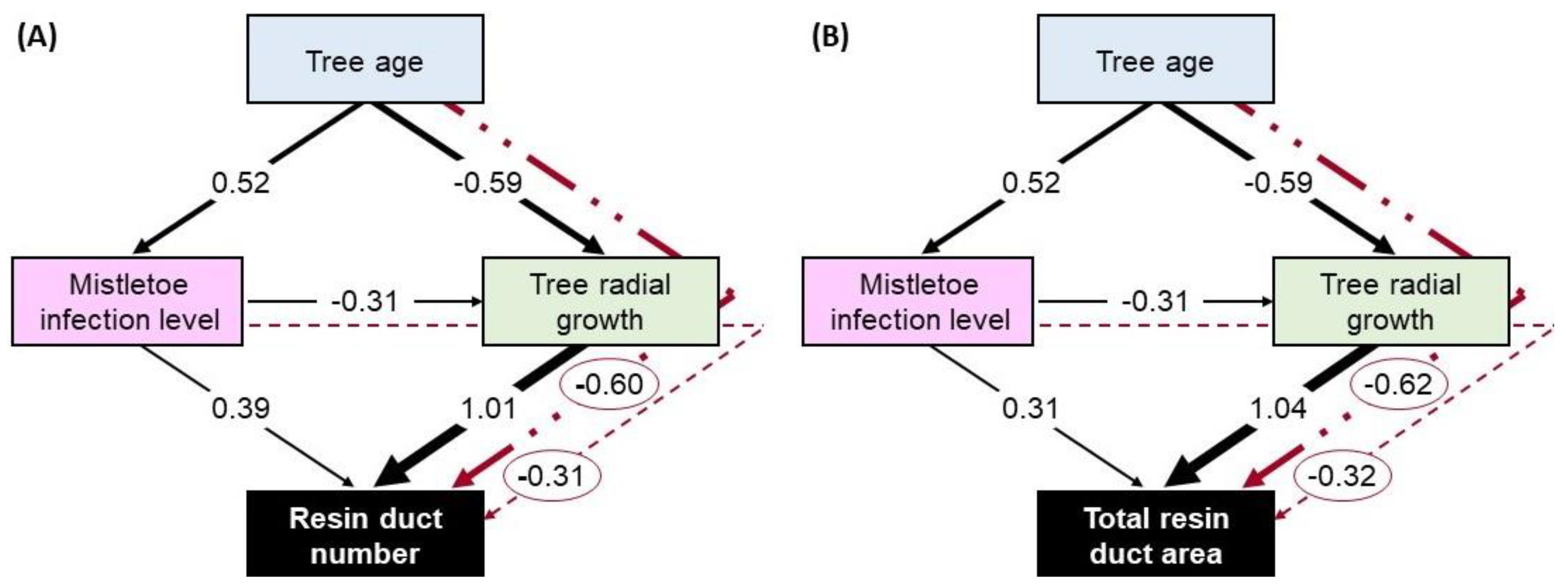
3.5. Hypothesized Direct and Indirect Effects of Mistletoe on Resin Duct Defenses
4. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Trapp, S.; Croteau, R. Defensive resin biosynthesis in conifers. Annu. Rev. Plant Biol. 2001, 52, 689–724. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, V.R.; Krokene, P.; Christiansen, E.; Krekling, T. Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytol. 2005, 167, 353–376. [Google Scholar] [CrossRef] [PubMed]
- Kolosova, N.; Bohlmann, J. Conifer defense against insects and fungal pathogens. In Global Change and Arctic Terrestrial Ecosystems; Springer: Berlin/Heidelberg, Germany, 2012; pp. 85–109. [Google Scholar]
- Kane, J.M.; Kolb, T.E. Importance of resin ducts in reducing ponderosa pine mortality from bark beetle attack. Oecologia 2010, 164, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Ferrenberg, S.; Kane, J.M.; Mitton, J.B. Resin duct characteristics associated with tree resistance to bark beetles across lodgepole and limber pines. Oecologia 2014, 174, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Hood, S.; Sala, A.; Heyerdahl, E.K.; Boutin, M. Low-severity fire increases tree defense against bark beetle attacks. Ecology 2015, 96, 1846–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampedro, L.; Moreira, X.; Zas, R. Costs of constitutive and herbivore-induced chemical defences in pine trees emerge only under low nutrient availability. J. Ecol. 2011, 99, 818–827. [Google Scholar] [CrossRef] [Green Version]
- Moreira, X.; Zas, R.; Solla, A.; Sampedro, L. Differentiation of persistent anatomical defensive structures is costly and determined by nutrient availability and genetic growth-defence constraints. Tree Physiol. 2015, 35, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Westbrook, J.W.; Walker, A.R.; Neves, L.G.; Munoz, P.; Resende, M.F., Jr.; Neale, D.B.; Peter, G.F. Discovering candidate genes that regulate resin canal number in Pinus taeda stems by integrating genetic analysis across environments, ages, and populations. New Phytol. 2015, 205, 627–641. [Google Scholar] [CrossRef] [Green Version]
- Ferrenberg, S. Landscape features and processes influencing forest pest dynamics. Curr. Landsc. Ecol. Rep. 2016, 1, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Moreira, X.; Mooney, K.A.; Rasmann, S.; Petry, W.K.; Carrillo-Gavilán, A.; Zas, R.; Sampedro, L. Trade-offs between constitutive and induced defences drive geographical and climatic clines in pine chemical defences. Ecol. Lett. 2014, 17, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Ferrenberg, S.; Langenhan, J.M.; Loskot, S.A.; Rozal, L.M.; Mitton, J.B. Resin monoterpene defenses decline within three widespread species of pine (Pinus) along a 1530-m elevational gradient. Ecosphere 2017, 8, e01975. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-González, C.; López-Goldar, X.; Zas, R.; Sampedro, L. Neutral and climate-driven adaptive processes contribute to explain population variation in resin duct traits in a mediterranean pine species. Front. Plant Sci. 2019, 10, 1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boege, K.; Marquis, R.J. Facing herbivory as you grow up: The ontogeny of resistance in plants. Trends Ecol. Evol. 2005, 20, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Barton, K.E.; Koricheva, J. The ontogeny of plant defense and herbivory: characterizing general patterns using meta-analysis. Am. Nat. 2010, 175, 481–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redmond, M.D.; Davis, T.S.; Ferrenberg, S.; Wion, A.P. Resource allocation trade-offs in a mast-seeding conifer: Piñon pine prioritizes reproduction over defence. AoB Plants 2019, 11, plz070. [Google Scholar] [CrossRef]
- Ferrenberg, S.; Kane, J.M.; Langenhan, J.M. To grow or defend? Pine seedlings grow less but induce more defences when a key resource is limited. Tree Physiol. 2015, 35, 107–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonello, P.; Gordon, T.R.; Herms, D.A.; Wood, D.L.; Erbilgin, N. Nature and ecological implications of pathogen-induced systemic resistance in conifers: A novel hypothesis. Physiol. Mol. Plant Pathol. 2006, 68, 95–104. [Google Scholar] [CrossRef]
- Wallis, C.; Eyles, A.; Chorbadjian, R.; Gardener, B.M.; Hansen, R.; Cipollini, D.; Herms, D.A.; Bonello, P. Systemic induction of phloem secondary metabolism and its relationship to resistance to a canker pathogen in Austrian pine. New Phytol. 2008, 177, 767–778. [Google Scholar] [CrossRef]
- Eyles, A.; Bonello, P.; Ganley, R.; Mohammed, C. Induced resistance to pests and pathogens in trees. New Phytol. 2010, 185, 893–908. [Google Scholar] [CrossRef]
- Meinzer, F.C.; Woodruff, D.R.; Shaw, D.C. Integrated responses of hydraulic architecture, water and carbon relations of western hemlock to dwarf mistletoe infection. Plant Cell Environ. 2004, 27, 937–946. [Google Scholar] [CrossRef]
- Rigling, A.; Eilmann, B.; Koechli, R.; Dobbertin, M. Mistletoe-induced crown degradation in Scots pine in a xeric environment. Tree Physiol. 2010, 30, 845–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCambridge, W.F.; Hawksworth, F.G.; Edminster, C.B.; Laut, J.G. Ponderosa Pine Mortality Resulting from a Mountain Pine Beetle Outbreak; Research Paper RM-RP-235; U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1982; 7p.
- Scott, J.M.; Mathiasen, R.L. Assessing growth and mortality of bristlecone pine infected by dwarf mistletoe using dendrochronology. For. Sci. 2012, 58, 366–376. [Google Scholar] [CrossRef]
- Hawksworth, F.G.; Geils, B.W. How long do mistletoe-infected ponderosa pines live? West. J. Appl. For. 1990, 5, 47–48. [Google Scholar] [CrossRef]
- Mathiasen, R.L.; Hawksworth, F.G.; Edminster, C.B. Effects of dwarf mistletoe on growth and mortality of Douglas-fir in the Southwest. Great Basin Nat. 1990, 50, 173–179. [Google Scholar]
- Filip, G.M.; Colbert, J.J.; Shaw, C.G., III; Hessburg, P.F.; Hosman, K.P. Influence of dwarf mistletoe and western spruce budworm on growth and mortality of Douglas-fir in unmanaged stands. For. Sci. 1993, 39, 465–477. [Google Scholar]
- Hawksworth, F.G.; Wiens, D. Dwarf mistletoes: Biology, pathology, and systematics. In Agricultural Handbook 709; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1996; 410p. [Google Scholar]
- Dobbertin, M.; Rigling, A. Pine mistletoe (Viscum album ssp. austriacum) contributes to Scots pine (Pinus sylvestris) mortality in the Rhone valley of Switzerland. For. Pathol. 2006, 36, 309–322. [Google Scholar] [CrossRef]
- Mehl, H.K.; Mori, S.R.; Frankel, S.J.; Rizzo, D.M. Mortality and growth of dwarf mistletoe-infected red and white fir and the efficacy of thinning for reducing associated losses. For. Pathol. 2013, 43, 193–203. [Google Scholar] [CrossRef]
- Agne, M.C.; Shaw, D.C.; Woolley, T.J.; Queijeiro-Bolaños, M.E. Effects of dwarf mistletoe on stand structure of lodgepole pine forests 21–28 years post-mountain pine beetle epidemic in central Oregon. PLoS ONE 2014, 9, e107532. [Google Scholar] [CrossRef] [Green Version]
- Bilgili, E.; Öztürk, M.; Coskuner, K.A.; Baysal, I.; Serdar, B.; Yavuz, H.; Eroglu, M.; Usta, Y. Quantifying the effect of pine mistletoe on the growth of Scots pine. For. Pathol. 2018, 48, e12435. [Google Scholar] [CrossRef]
- González-Elizondo, M.; Flores-Villegas, M.Y.; Álvarez-Zagoya, R.; González-Elizondo, M.S.; Márquez-Linares, M.A.; Quiñonez-Barraza, S.; Mathiasen, R.L. Effects of Mexican dwarf mistletoe (Arceuthobium vaginatum subsp. vaginatum) on the growth of Pinus cooperi in Durango, Mexico—A case study. For. Pathol. 2018, 49, e12473. [Google Scholar]
- Klutsch, J.G.; Erbilgin, N. Dwarf mistletoe infection in jack pine alters growth–defense relationships. Tree Physiol. 2018, 38, 1538–1547. [Google Scholar] [CrossRef] [PubMed]
- Stanton, S. Effects of dwarf mistletoe on climate response of mature ponderosa pine trees. Tree Ring Res. 2007, 63, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Ciesla, W.M. Report on the Health of Colorado’s Forests: Continuing Challenges for Colorado’s Forests: Recurring & Emerging Threats: 10th Anniversary Report; Colorado State Forest Service: Fort Collins, CO, USA, 2010. [Google Scholar]
- Geils, B.W.; Mathiasen, R.L. Intensification of dwarf mistletoe on southwestern Douglas-fir. For. Sci. 1990, 36, 955–969. [Google Scholar]
- Duhl, T.R.; Gochis, D.; Guenther, A.; Ferrenberg, S.; Pendall, E. Emissions of BVOC from lodgepole pine in response to mountain pine beetle attack in high and low mortality forest stands. Biogeosciences 2013, 10, 483–499. [Google Scholar] [CrossRef] [Green Version]
- Mitton, J.B.; Ferrenberg, S.M. Mountain pine beetle develops an unprecedented summer generation in response to climate warming. Am. Nat. 2012, 179, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Hawksworth, F.G.; Hinds, T.E. Effects of dwarf mistletoe on immature lodgepole pine stands in Colorado. J. For. 1964, 62, 27–32. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Tinevez, J.Y. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676. [Google Scholar] [CrossRef] [Green Version]
- Chhin, S.; Hogg, E.H.; Lieffers, V.J.; Huang, S. Growth-climate relationships vary with height along the stem in lodgepole pine. Tree Physiol. 2010, 30, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; Mérian, P.; Qeadan, F.; Zang, C. dplR: Dendrochronology Program Library in R. R package version 1.7.0. 2019. Available online: https://CRAN.R-project.org/package=dplR (accessed on 21 July 2019).
- Stephenson, N.L.; Das, A.J.; Condit, R.; Russo, S.E.; Baker, P.J.; Beckman, N.G.; Coomes, D.A.; Lines, E.R.; Morris, W.K.; Rüger, N.; et al. Rate of tree carbon accumulation increases continuously with tree size. Nature 2014, 507, 90–93. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Barton, B.K. MuMIn: Multi-Model Inference. R Package Version 1.15. 6. 2016. Available online: https://cran.r-project.org/web/packages/MuMIn/MuMIn.pdf (accessed on 2 January 2020).
- Wheeler, B.; Torchiano, M. lmPerm: Permutation tests for linear models. R Package Vers. 2010, 1, 1–2. [Google Scholar]
- McKelvey, R.D.; Zavoina, W. A statistical model for the analysis of ordinal level dependent variables. J. Math. Sociol. 1975, 4, 103–120. [Google Scholar] [CrossRef]
- Beaujean, A.A.; Beaujean, M.A.A. Package ‘BaylorEdPsych’. 2015. Available online: https://cran.r-project.org/web/packages/BaylorEdPsych/BaylorEdPsych.pdf (accessed on 2 January 2020).
- Windmeijer, F.A. Goodness-of-fit measures in binary choice models1. Econ. Rev. 1995, 14, 101–116. [Google Scholar] [CrossRef]
- Rosseel, Y. Lavaan: An R package for structural equation modeling and more. Version 0.5–12 (BETA). J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Koricheva, J. Meta-analysis of sources of variation in fitness costs of plant antiherbivore defenses. Ecology 2002, 83, 176–190. [Google Scholar] [CrossRef]
- Stamp, N. Out of the quagmire of plant defense hypotheses. Q. Rev. Boil. 2003, 78, 23–55. [Google Scholar] [CrossRef] [Green Version]
- Enquist, B.J.; West, G.B.; Charnov, E.L.; Brown, J.H. Allometric scaling of production and life-history variation in vascular plants. Nature 1999, 401, 907–911. [Google Scholar] [CrossRef]
- Weiner, J.; Thomas, S.C. The nature of tree growth and the “age-related decline in forest productivity”. Oikos 2001, 94, 374–376. [Google Scholar] [CrossRef] [Green Version]
- Sala, A.; Carey, E.V.; Callaway, R.M. Dwarf mistletoe affects whole-tree water relations of Douglas fir and western larch primarily through changes in leaf to sapwood ratios. Oecologia 2001, 126, 42–52. [Google Scholar] [CrossRef]
- Mathiasen, R.L. Dwarf mistletoes in forest canopies. Northwest Sci. 1996, 70, 61–71. [Google Scholar]
- Barbosa, J.M.; Sebástian-González, E.; Asner, G.P.; Knapp, D.E.; Anderson, C.; Martin, R.E.; Dirzo, R. Hemiparasite-host plant interactions in a fragmented landscape assessed via imaging spectroscopy and LiDAR. Ecol. Appl. 2016, 26, 55–66. [Google Scholar] [CrossRef] [PubMed]
- MacRaild, L.M.; Radford, J.Q.; Bennett, A.F. Non-linear effects of landscape properties on mistletoe parasitism in fragmented agricultural landscapes. Landsc. Ecol. 2010, 25, 395–406. [Google Scholar] [CrossRef]
- Goodsman, D.W.; Lusebrink, I.; Landhäusser, S.M.; Erbilgin, N.; Lieffers, V.J. Variation in carbon availability, defense chemistry and susceptibility to fungal invasion along the stems of mature trees. New Phytol. 2013, 197, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Ferrenberg, S.; Mitton, J.B. Smooth bark surfaces can defend trees against insect attack: Resurrecting a ‘slippery’ hypothesis. Funct. Ecol. 2014, 28, 837–845. [Google Scholar] [CrossRef]
- Quiring, D.T. Rapid change in suitability of white spruce for a specialist herbivore, Zeiraphera canadensis, as a function of leaf age. Can. J. Zool. 1992, 70, 2132–2138. [Google Scholar] [CrossRef]
- Boege, K.; Barton, K.E.; Dirzo, R. Influence of tree ontogeny on plant-herbivore interactions. In Size-and Age-Related Changes in Tree Structure and Function; Meinzer, F.C., Lachenbruch, B., Dawson, T.E., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 193–214. [Google Scholar]
- Zhao, S.; Erbilgin, N. Larger resin ducts are linked to the survival of lodgepole pine trees during mountain pine beetle outbreak. Front. Plant Sci. 2019, 10, 1459. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrenberg, S. Dwarf Mistletoe Infection Interacts with Tree Growth Rate to Produce Opposing Direct and Indirect Effects on Resin Duct Defenses in Lodgepole Pine. Forests 2020, 11, 222. https://doi.org/10.3390/f11020222
Ferrenberg S. Dwarf Mistletoe Infection Interacts with Tree Growth Rate to Produce Opposing Direct and Indirect Effects on Resin Duct Defenses in Lodgepole Pine. Forests. 2020; 11(2):222. https://doi.org/10.3390/f11020222
Chicago/Turabian StyleFerrenberg, Scott. 2020. "Dwarf Mistletoe Infection Interacts with Tree Growth Rate to Produce Opposing Direct and Indirect Effects on Resin Duct Defenses in Lodgepole Pine" Forests 11, no. 2: 222. https://doi.org/10.3390/f11020222
APA StyleFerrenberg, S. (2020). Dwarf Mistletoe Infection Interacts with Tree Growth Rate to Produce Opposing Direct and Indirect Effects on Resin Duct Defenses in Lodgepole Pine. Forests, 11(2), 222. https://doi.org/10.3390/f11020222