Modelling the Crown Profile of Western Hemlock (Tsuga heterophylla) with a Combination of Component and Aggregate Measures of Crown Size

Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Analyses
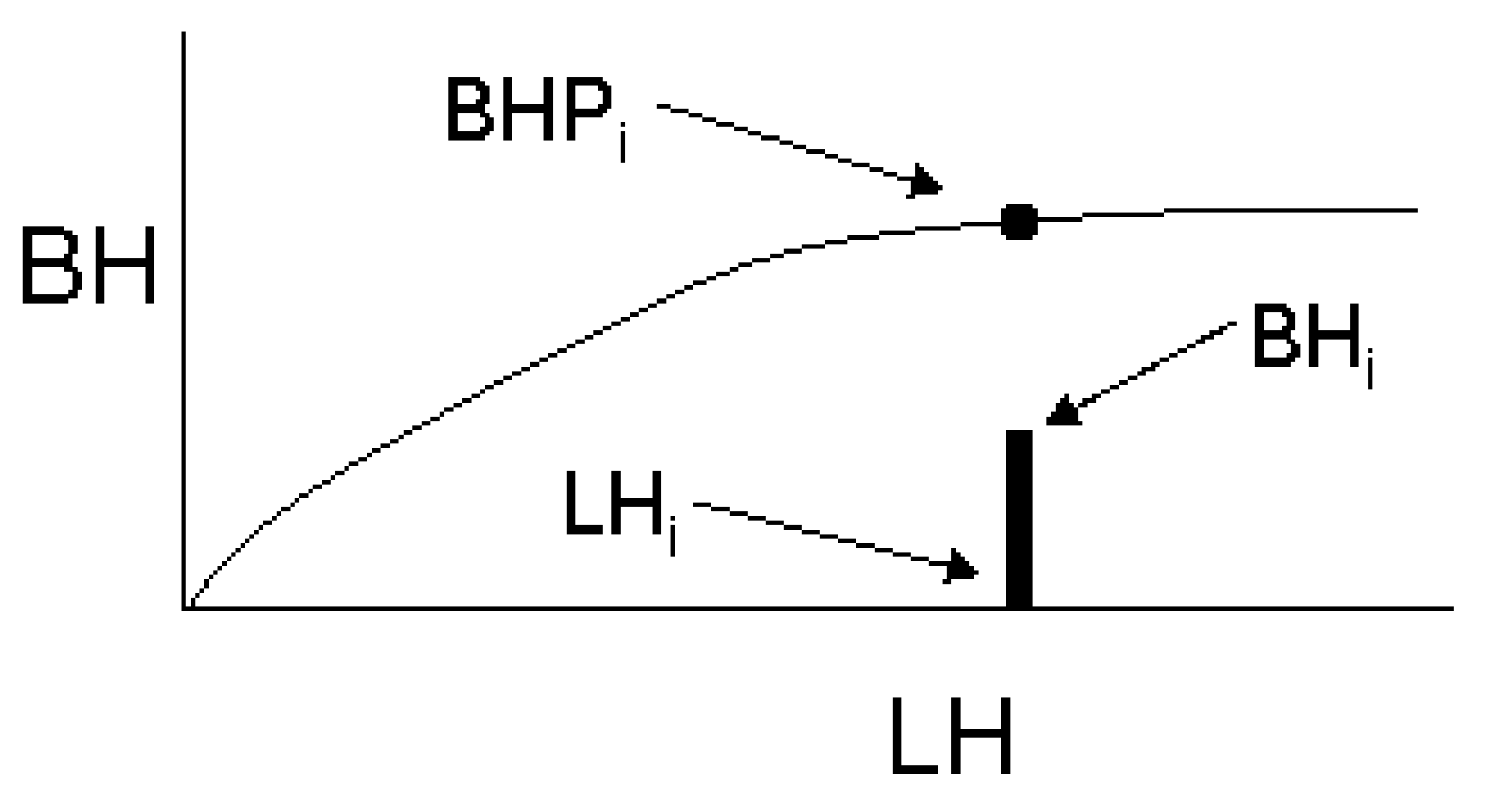
2.2.1. Modelling the Component Measures
2.2.2. Adjusting the Crown Profile with Ground Estimates of Crown Projection Area (CPAG)
3. Results
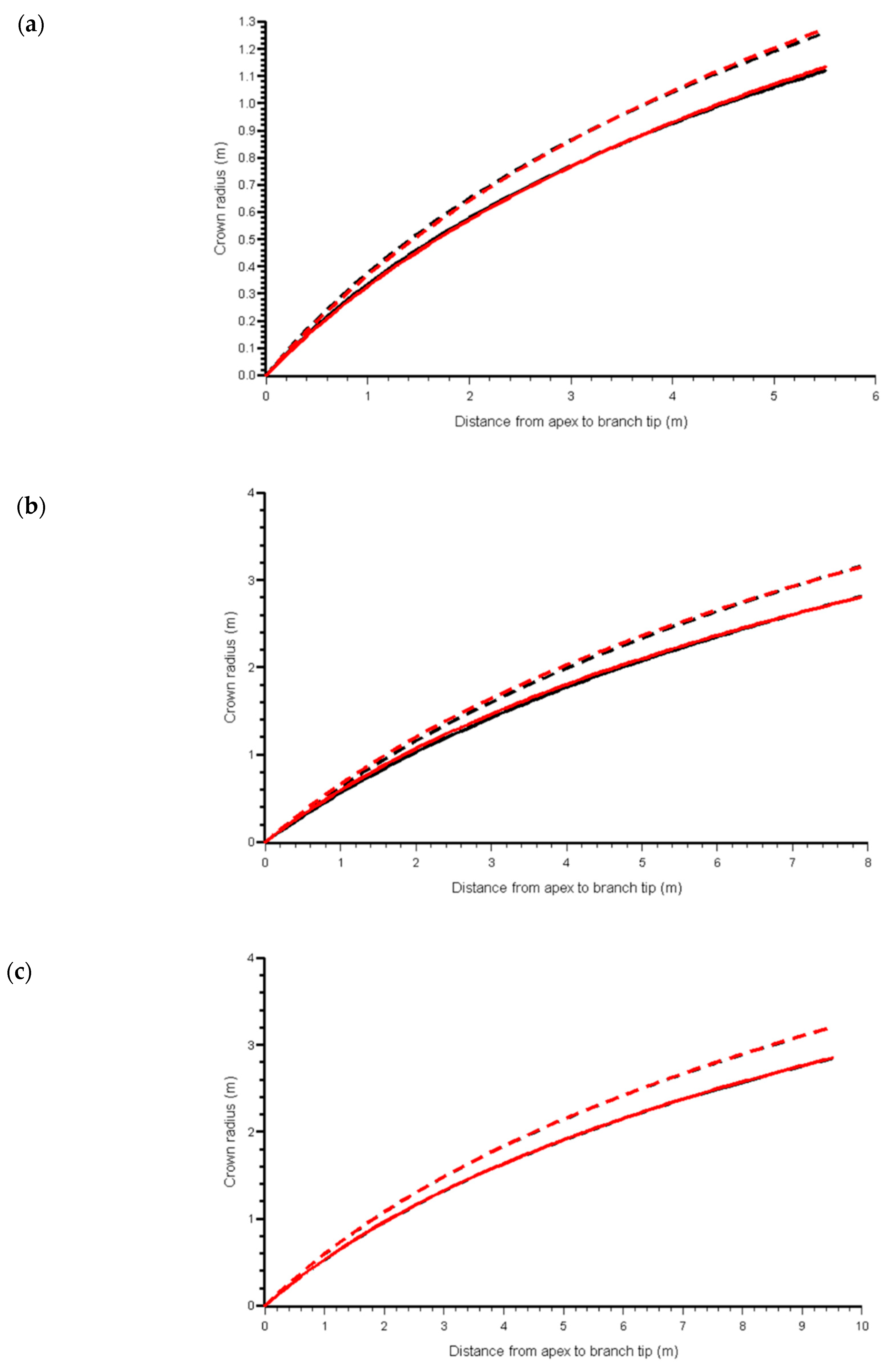
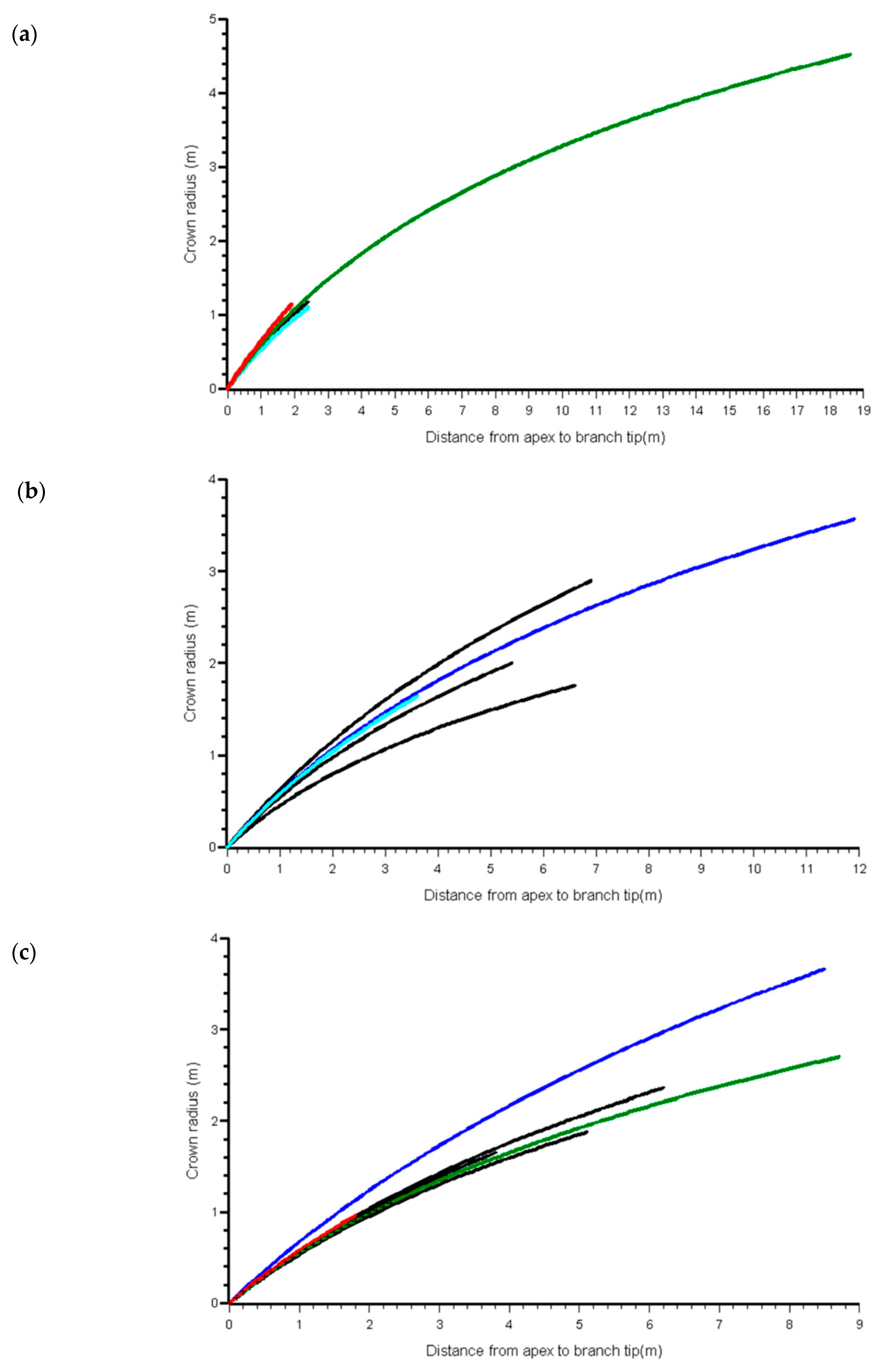
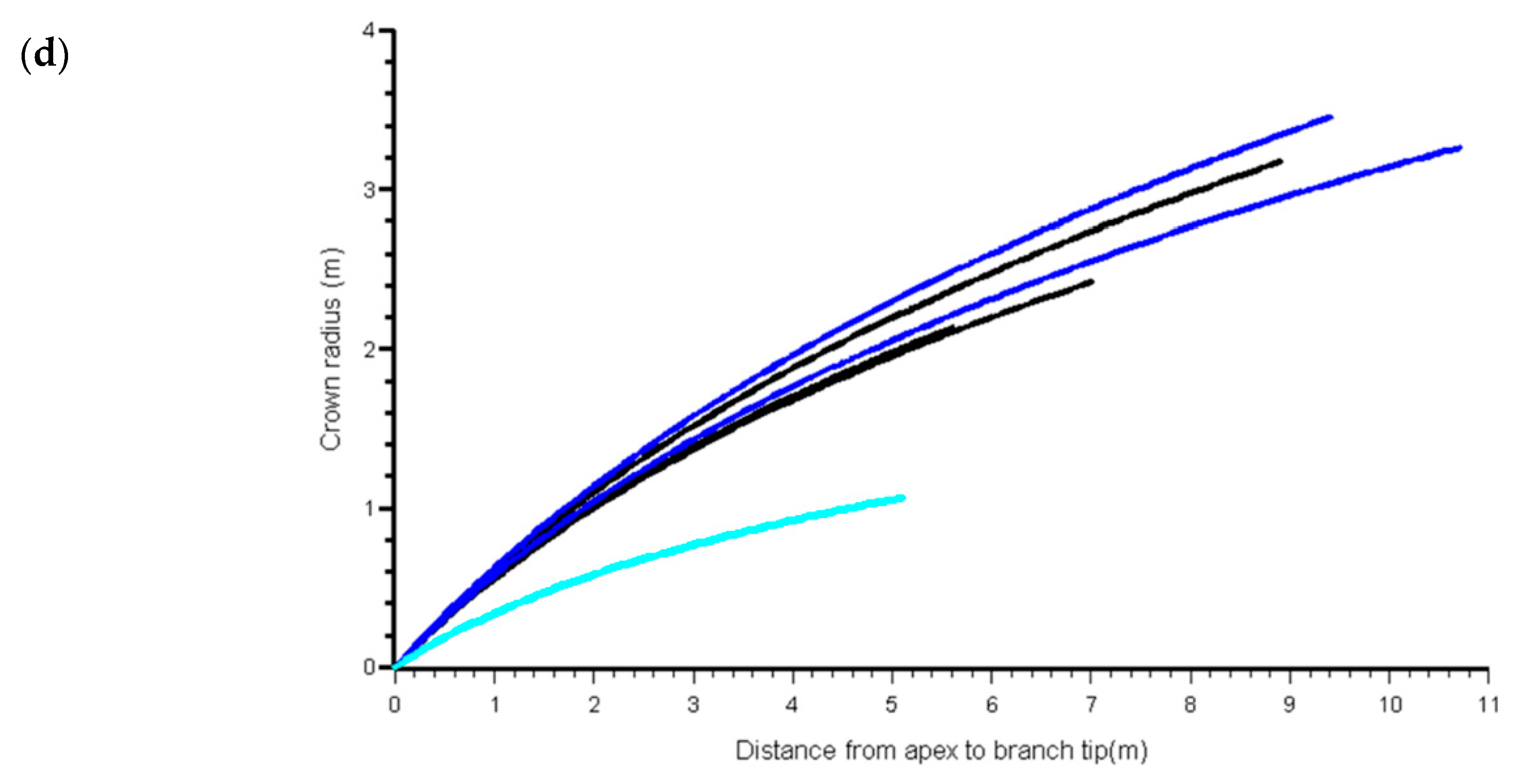
3.1. Component Measures
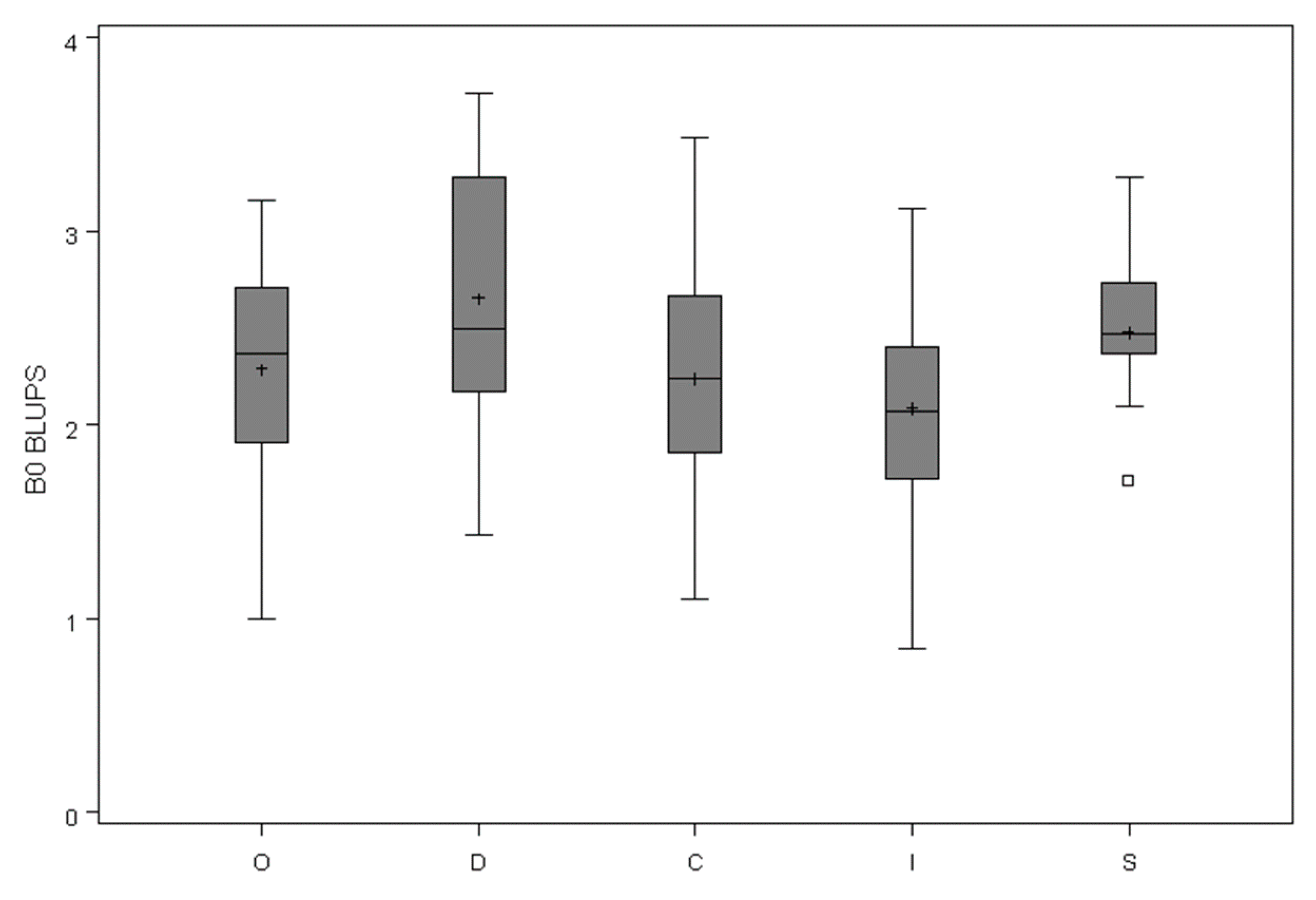
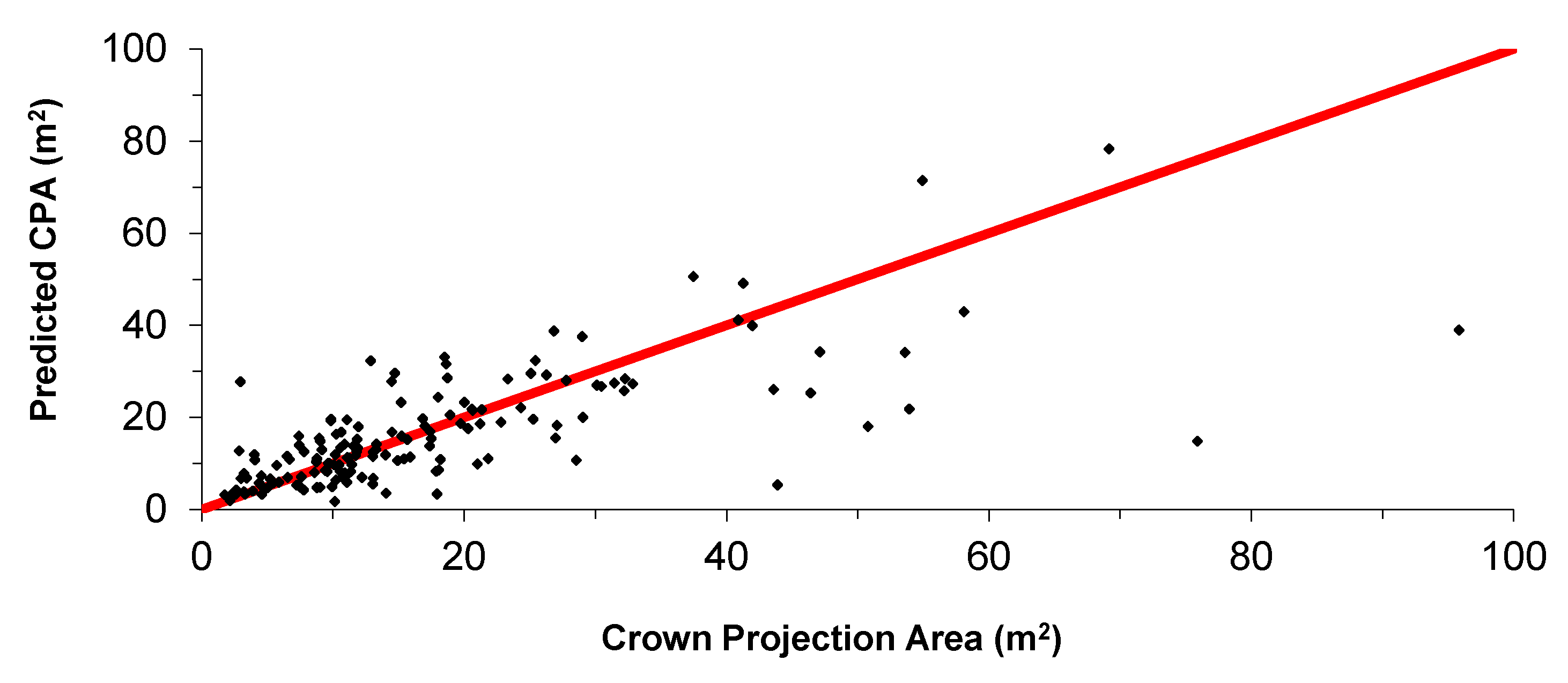
3.2. Adjusting the Crown Profile with Ground Estimates of Crown Projection Area (CPAG)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stenberg, P.; Kuuluvainen, T.; Kellomäki, S.; Grace, J.C.; Jokela, E.J.; Gholz, H.L. Crown structure, light interception and productivity of pine trees and stands. Ecol. Bull. 1994, 43, 20–34. [Google Scholar]
- Brunner, A. A light model for spatially explicit forest stand models. For. Ecol. Manag. 1998, 7, 19–46. [Google Scholar] [CrossRef]
- Brunner, A.; Nigh, G. Light absorption and bole volume growth of individual Douglas-fir trees. Tree Physiol. 2000, 20, 323–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentine, H.T.; Mäkelä, A. Bridging process-based and empirical approaches to modeling tree growth. Tree Physiol. 2005, 25, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Rudnicki, M.; Silins, U.; Lieffers, V.J.; Josi, G. Measure of simultaneous tree sways and estimation of crown interactions among a group of trees. Trees 2001, 15, 83–90. [Google Scholar] [CrossRef]
- Mitchell, K.J. Dynamics and simulated yield of Douglas-fir. For. Sci. Monogr. 1975, 17, 39. [Google Scholar]
- Larson, P.R. Stem form development of forest trees. For. Sci. Monogr. 1963, 5, 42. [Google Scholar] [CrossRef]
- Houllier, F.; Leban, J.-M.; Cohn, F. Linking growth modelling to timber quality assessment for Norway spruce. For. Ecol. Manag. 1995, 74, 91–102. [Google Scholar] [CrossRef]
- Mansfield, S.D.; Parish, R.; Goudie, J.W.; Kang, K.-Y.; Ott, P. The effects of crown ratio on the transition from juvenile to mature wood production in lodgepole pine in western Canada. Can. J. For. Res. 2007, 37, 1450–1459. [Google Scholar] [CrossRef]
- Hann, D.W.; Hester, A.S.; Olsen, C.L. ORGANON user’s manual: Edition 5.1 Southwest Oregon version and western Willamette Valley version; Oregon State University: Corvallis, OR, USA, 1995; 113p. [Google Scholar]
- Wykoff, W.R.; Crookston, N.L.; Stage, A.R. User’s guide to the stand prognosis model. USDA For. Serv. Gen. Tech. Rep. 1982, INT-133, 112. [Google Scholar]
- Mäkelä, A. Modeling structural-functional relationships in whole-tree growth: Resource allocation. In Process Modeling of Forest Growth Responses to Environmental Stress; Dixon, R.K., Meldahl, R.S., Ruark, G.A., Warren, W.G., Eds.; Timber Press: Portland, OR, USA, 1990; pp. 81–95. [Google Scholar]
- Sievänen, R.; Mäkelä, A.; Nikinmaa, E.; Korpilahti, E. Preface to “Helsinki Workshop on Functional-Structural Tree Models”. Silva Fenn. 1997, 31, 237–238. [Google Scholar]
- Cluzeau, C.; Le Goff, N.; Ottorini, J.-M. Development of primary branches and crown profile of Fraxinus excelsior. Can. J. For. Res. 1994, 24, 2315–2323. [Google Scholar] [CrossRef]
- Crecente-Campo, F.; Marshall, P.; LeMay, V.; Dieguez-Aranda, U. A crown profile model for Pinus radiata D. Don in northwestern Spain. For. Ecol. Manag. 2009, 257, 2370–2379. [Google Scholar] [CrossRef]
- Baldwin, V.C., Jr.; Peterson, K.D. Predicting the crown shape of loblolly pine trees. Can. J. For. Res. 1997, 27, 102–107. [Google Scholar] [CrossRef]
- Gill, S.J.; Biging, G.S. Autoregressive moving average models of conifer crown profiles. J. Agric. Biol. Environ. Stat. 2002, 7, 558–573. [Google Scholar] [CrossRef]
- Sheng, Y.W.; Gong, P.; Biging, G.S. Model-based conifer canopy surface reconstruction from photographic imagery: overcoming the occlusion, foreshortening, and edge effects. Photogramm. Eng. Remote Sens. 2003, 69, 249–258. [Google Scholar] [CrossRef] [Green Version]
- Martin-Ducup, O.; Schneider, R.; Fournier, R.A. A method to quantify canopy changes using multi-temporal terrestrial lidar data: Tree response to surrounding gaps. Agric. For. Meteorol. 2017, 237–238, 184–195. [Google Scholar]
- Dalponte, M.; Coomes, D.A. Tree-centric mapping of forest carbon density from airborne laser scanning and hyperspectral data. Methods Ecol. Evol. 2016, 7, 1236–1245. [Google Scholar] [CrossRef] [Green Version]
- Roeh, R.L.; Maguire, D.A. Crown profile models based on branch attributes in coastal Douglas-fir. For. Ecol. Manag. 1997, 96, 77–100. [Google Scholar] [CrossRef]
- Nepal, S.; Somers, G.; Caudill, S. A stochastic frontier model for fitting tree crown shape in loblolly pine. J. Agric. Biol. Environ. Stat. 1996, 1, 336–353. [Google Scholar] [CrossRef]
- Cochrane, L.A.; Ford, E.D. Growth of a Sitka spruce plantation: Analysis and stochastic description of the development of the branching structure. J. Appl. Ecol. 1978, 15, 227–244. [Google Scholar] [CrossRef]
- Valentine, H.T.; Tritton, L.M.; Furnival, G.M. Subsampling trees for biomass, volume, or mineral content. For. Sci. 1984, 30, 673–681. [Google Scholar]
- Halle, F.; Oldeman, R.A.A.; Tomlinson, P.B. Tropical trees and forests: An Architectural Analysis; Springer: New York, NY, USA, 1978; 444p. [Google Scholar]
- Tomlinson, P.B. Architecture of tropical plants. Ann. Rev. Ecol. Syst. 1987, 18, 1–21. [Google Scholar] [CrossRef]
- Goudie, J.W.; Parish, R.; Antos, J.A. Foliage biomass and specific leaf area equations at the branch, annual shoot and whole-tree levels for lodgepole pine and white spruce in British Columbia. For. Ecol. Manag. 2016, 361, 286–297. [Google Scholar] [CrossRef]
- Gregoire, T.G. Design-based and model-based inference in survey sampling: Appreciating the difference. Can. J. For. Res. 1998, 28, 1429–1447. [Google Scholar] [CrossRef]
- Burkhart, H.E.; Tomé, M. Modeling Forest Trees and Stands; Springer: New York, NY, USA, 2012; 457p. [Google Scholar]
- Husch, B.; Miller, C.I.; Beers, T.W. Forest Mensuration, 2nd ed.; Ronald Press: New York, NY, USA, 1972; 410p. [Google Scholar]
- Walters, J. The branch arrangement of western hemlock (Tsuga heterophylla [Raf.] Sarg.). Univ. B. C. Fac. For. Res. Note 1960, 29. [Google Scholar]
- Mitchell, K.J.; Cameron, I.R. Managed stand yield tables for coastal Douglas-fir: initial density and precommercial thinning. B.C. Min. For. Res. Branch Land Manag. Rep. 1985, 31, 69. [Google Scholar]
- SAS. SAS/STAT Version 9; SAS Institute Inc.: Cary, NC, USA, 2008. [Google Scholar]
- Schabenberger, O.; Pierce, F.J. Contemporary Statistical Models for the Plant and Soil Sciences; CRC Press: Boca Raton, FL, USA, 2002; 738p. [Google Scholar]
- Leites, L.P.; Robinson, A.P. Improving taper equations of loblolly pine with crown dimensions in a mixed-effects modeling framework. For. Sci. 2002, 50, 204–212. [Google Scholar]
- Hann, D.W. An adjustable predictor of crown profile for stand-grown Douglas-fir trees. For. Sci. 1999, 45, 217–225. [Google Scholar]
- Wang, C.; Wu, B.; Chen, Y.; Qi, Y. Development of crown profile models for Chinese Fir using non-linear mixed-effects modelling. Nat. Environ. Pollut. Tech. 2019, 18, 349–1361. [Google Scholar]
- Marshall, D.D.; Johnson, G.P.; Hann, D.W. Crown profile equations for stand- grown western hemlock trees in northwestern Oregon. Can. J. For. Res. 2003, 33, 2059–2066. [Google Scholar] [CrossRef] [Green Version]
- Schoettle, A.E.; Smith, W.K. Interrelation between shoot characteristics and solar irradiance in the crown of Pinus contorta ssp. latifolia. Tree Physiol. 1991, 9, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Rouvinen, S.; Kuuluvainen, T. Structure and asymmetry of tree crowns in relation to local competition in a natural mature Scots pine forest. Can. J. For. Res. 1997, 27, 890–902. [Google Scholar] [CrossRef]
- Grace, J.C.; Pont, D.; Goulding, C.J. Modelling branch development for forest management. N. Z. J. For. Sci. 1999, 29, 391–408. [Google Scholar]
- Cannell, M.G.R.; Bowler, K.C. Spatial arrangement of lateral buds at the time that they form on leaders of Picea and Larix. Can. J. For. Res. 1978, 8, 129–137. [Google Scholar] [CrossRef]
- Doruska, P.F.; Burkhart, H.E. Modeling the diameter and locational distribution of branches within crowns of loblolly pine in unthinned plantations. Can. J. For. Res. 1994, 24, 2362–2376. [Google Scholar] [CrossRef]
- Gao, H.; Dong, L.; Fengri, L. Modeling variation in crown profile with tree status and cardinal directions for planted Larix olgensis Henry trees in northeast China. Forests 2017, 8, 139–154. [Google Scholar]
- Skatter, S.; Kucera, B. The cause of the prevalent directions of the spiral grain patterns in conifers. Trees 1998, 12, 265–273. [Google Scholar] [CrossRef]
- Achim, A.; Gardiner, B.; Leban, J.-M.; Daquitane, R. Predicting the branching properties of Sitka spruce grown in Great Britain. N. Z. J. For. Sci. 2006, 36, 246–264. [Google Scholar]
- Raulier, F.; Ung, C.-H.; Ouellet, D. Influence of social status on crown geometry and volume increment in regular and irregular black spruce stands. Can. J. For. Res. 1996, 26, 1742–1753. [Google Scholar] [CrossRef]
- Kantola, A.; Mäkelä, A. Crown development in Norway spruce [Picea abies (L.) Karst]. Trees 2004, 18, 408–421. [Google Scholar] [CrossRef]
- Gregoire, T.G.; Valentine, H.T. A sampling strategy to estimate the area and perimeter of irregularly shaped planar regions. For. Sci. 1995, 41, 470–476. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Number of Trees | Number of Branches | Age (years) | Height (m) | Crown Length (m) | Crown Projection Area (m2) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| min | avg | max | min | avg | max | min | avg | max | min | avg | max | |||
| G86 | 20 | 129 | 9 | 41 | 91 | 6.3 | 21.0 | 38.5 | 3.9 | 10.5 | 23.6 | 2.8 | 18.8 | 69.2 |
| W98 | 85 | 878 | 12 | 47 | 111 | 6.9 | 25.6 | 45.1 | 3.7 | 12.8 | 27.5 | 1.7 | 20.0 | 95.8 |
| P99 | 48 | 609 | 16 | 18 | 19 | 7.8 | 12.4 | 16.0 | 5.5 | 9.8 | 13.3 | 5.9 | 13.2 | 24.3 |
| Source | Internode Selection | Branch Selection | Details |
|---|---|---|---|
| G86 | Systematic | LNG2 | Collected in five locations by the former MacMillan Bloedel Ltd. on Vancouver Island in 1986 |
| W98 | Stratified Random | PPS3 | Collected in four stands of different ages in four geographic locations in coastal BC in 1998 |
| P99 | Stratified Random | PPS3 | Collected from a hemlock progeny trial on two sites in coastal BC in 1999 |
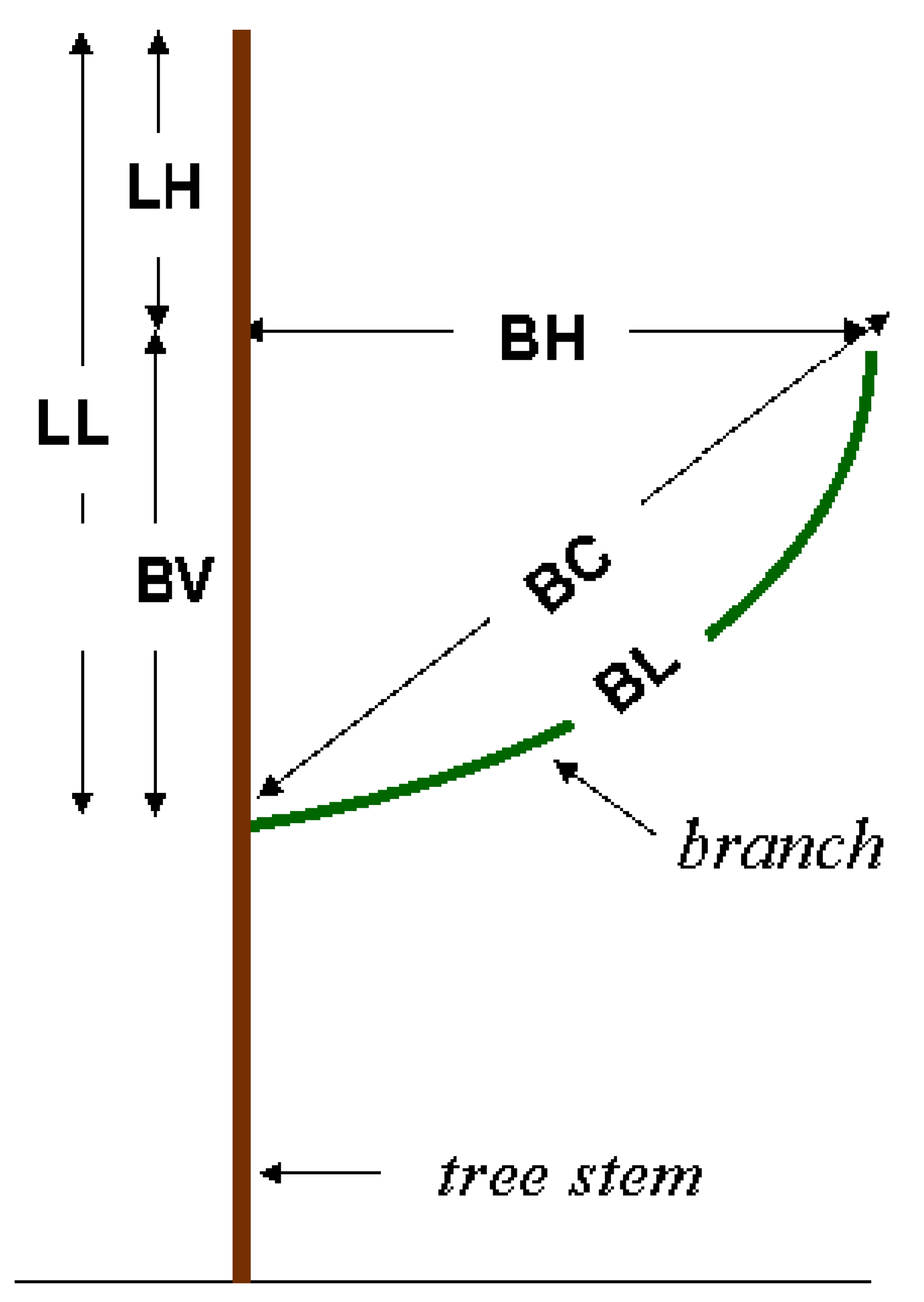
| Variable | Definition |
|---|---|
| BC | Chord length from the branch origin to the branch tip. |
| BL | Total length of the branch along the main branch axis. |
| BH | Horizontal distance from the stem to the branch tip. |
| BV | Vertical distance from the branch origin to the projection of the branch tip. |
| LL | Vertical distance from the branch origin to the apex of the tree. |
| LH | Vertical distance from the branch tip to the apex of the tree. |
| b0 | b1 | b2 | s02 | s12 | s2 | −2 Log Likelihood | AIC | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Model | Estimate | SE | Estimate | SE | Estimate | SE | Estimate | SE | Estimate | SE | Estimate | SE | ||
| ML | 2.40 | 0.138 | 4.00 | 0.309 | 0.49 | 0.092 | 1.648 | 0.193 | 0.056 | 0.002 | 266.5 | 278.5 | ||
| MP | 2.44 | 0.113 | 4.04 | 0.252 | 1.09 | 0.020 | 0.55 | 0.078 | 1.188 | 0.243 | 0.054 | 0.002 | 239.7 | 253.7 |
| MB0* | 2.27 | 0.123 | 1.09 | 0.021 | 0.43 | 0.135 | 0.059 | 0.002 | 310.6 | 322.6 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cameron, I.R.; Parish, R.; Goudie, J.W.; Bealle Statland, C.A. Modelling the Crown Profile of Western Hemlock (Tsuga heterophylla) with a Combination of Component and Aggregate Measures of Crown Size. Forests 2020, 11, 281. https://doi.org/10.3390/f11030281
Cameron IR, Parish R, Goudie JW, Bealle Statland CA. Modelling the Crown Profile of Western Hemlock (Tsuga heterophylla) with a Combination of Component and Aggregate Measures of Crown Size. Forests. 2020; 11(3):281. https://doi.org/10.3390/f11030281
Chicago/Turabian StyleCameron, Ian R., Roberta Parish, James W. Goudie, and Catherine A. Bealle Statland. 2020. "Modelling the Crown Profile of Western Hemlock (Tsuga heterophylla) with a Combination of Component and Aggregate Measures of Crown Size" Forests 11, no. 3: 281. https://doi.org/10.3390/f11030281
APA StyleCameron, I. R., Parish, R., Goudie, J. W., & Bealle Statland, C. A. (2020). Modelling the Crown Profile of Western Hemlock (Tsuga heterophylla) with a Combination of Component and Aggregate Measures of Crown Size. Forests, 11(3), 281. https://doi.org/10.3390/f11030281