1. Introduction
Wind loads often induce tree failure, and many studies have quantified the effects of wind events on forests. In an extensive review, Everham and Brokaw [
1] synthesized the findings of 42 studies of tree failure during catastrophic wind events to compile a database of wind resistance ratings for different species in temperate and tropical climates. However, their synthesis only included studies on forest- or plantation-grown trees, which resulted in two important limitations. First, the form of open-grown trees differs from that of forest- or plantation-grown trees [
2]; secondly, ornamental species common in residential landscapes are typically absent from natural forest stands or plantations. Studies have examined the wind-induced failure of open-grown trees in temperate [
3] and tropical and subtropical climates [
4,
5,
6,
7], but predicting failure is problematic because empirical data on relevant parameters are sparse.
Hurricanes annually impact communities along the Atlantic and Gulf Coasts in the United States, sometimes causing widespread and costly damage. Hurricanes often induce tree failure, and studies have (i) quantified the cost of tree damage and debris [
8], (ii) evaluated the likelihood of tree failure during catastrophic wind events [
1,
4,
5,
6,
7,
9], (iii) assessed damaged trees [
10] and property [
11,
12], (iv) considered their effect on urban tree populations [
12], and (v) gauged perceptions of the urban forest following a hurricane [
13]. However, predicting the likelihood of tree failure during storms remains a challenge.
Many factors influence the likelihood of tree failure. Factors can be categorized as intrinsic or extrinsic to the tree. Intrinsic factors include size, wood density, and the presence or absence of structural defects (e.g., decay, weakly attached branches) and foliage. Studies have consistently reported a greater likelihood of failure for taller trees with larger trunk diameter for trees in both cooler [
3,
9,
14,
15] and warmer climates [
4,
11]. Larger trees experience greater drag and drag-induced bending moments because of a greater crown area, a higher center of pressure, and greater exposure to the wind. The likelihood of tree failure tends to be lower with denser wood [
7,
11,
15,
16,
17,
18]. Leafless trees experience considerably less drag than in-leaf trees [
19], which presumably explains the lower likelihood of failure in catastrophic wind events of dormant trees [
9] or trees that readily shed leaves [
4,
20]. An integral part of visual tree assessment is identifying the presence and severity of structural defects [
21,
22,
23,
24,
25], but only severe defects, rather than simply the presence of defects, have been linked to increased likelihood of tree failure [
9].
Extrinsic factors, such as site conditions, can influence and even mask species-specific intrinsic factors because they influence growth form. For several species, the form of open-grown individuals differed substantially from that of forest- or plantation-grown trees [
2]. However, within a particular site, many studies have demonstrated that the likelihood of failure varies among species [
1,
3,
4,
6,
7,
9,
11,
20]. Studies have also shown instances where native species were more resilient to wind than nonnative species [
5,
12,
26] especially in hurricane-prone regions, but the converse has also been documented [
11,
27].
In developed landscapes, likelihood of failure was lower when trees had a greater volume of soil for roots [
11,
12], and among trees growing in clusters [
12]. In developed landscapes, management practices may also influence the likelihood of failure in catastrophic wind events, but a consensus is lacking among the few studies that have investigated this effect. Some studies have found that pruned trees were less likely to fail than trees that had not been pruned [
5,
11,
12], but one [
9] reported a minimal effect of pruning on the likelihood of failure. It is also important to remember that the effects of management may be negligible when winds are catastrophic and sufficient to cause even structurally sound trees to fail.
Given the complexity of modeling tree failure in catastrophic wind events, species failure profiles can be useful. Failure profiles commonly involve an evaluation of structural and environmental factors that lead to tree failure. For example, Edberg and Berry [
28] developed a failure profile for coast live oak in California (
Quercus agrifolia Née) using data from the California Tree Failure Report Program (CTFRP). Structural factors included the location of the failure (trunk; branch; root; above, at, or below ground level); environmental factors included precipitation, wind speed, soil, and topography. They also considered the size of the tree, stand characteristics, common defects, cultural practices, and the presence of decay [
28]. The most common type of failure found in coast live oak (83%) was related to decay found in the lower trunk and roots, resulting from saturated soil conditions [
28]. Edberg et al. [
29] also used data from the CTFRP to develop a failure profile for Monterey pine (
Pinus radiata D. Don)and found that 75% of failures were of the heavy lateral limbs; of these, 74% failed away from the branch attachment, suggesting that wood strength or load distribution, not a weak branch attachment, was the cause.
In the United States, hurricanes regularly occur in Florida, providing ample opportunities to develop species failure profiles [
5,
10,
11,
12]. Since climate change models predict an increase in hurricane frequency and severity, the objectives of this study were to determine (i) factors that influenced the likelihood of failure, and (ii) what type of failure was most common.
2. Materials and Methods
Hurricane Irma reached coastal Florida as a category 3 hurricane on 10 September 2017. As it moved northward to Naples, Florida, USA (26.1420° N, 81.7948° W), the storm diminished in strength, with the southern and eastern portions of the city experiencing category 2 hurricane winds and the northern portion of the city experiencing category 1 winds [
30]. According to the Saffir–Simpson scale, category 2 hurricanes will cause “many shallowly rooted trees [to] be snapped or uprooted and block numerous roads” and category 1 hurricanes will cause “large branches of trees [to] snap and shallowly rooted trees may be toppled” [
31]. Prior to Hurricane Irma, Naples had a complete inventory of 19,638 public trees, which included 79 species. The current study focused on a subset of the inventory that included city-owned street trees in the public right-of-way, but excluded trees in parks and medians (i.e., the strip of unpaved land that separates lanes of opposing traffic on a divided highway).
Following the hurricane and initial cleanup, from 30 October 2017, through 3 November 2017, the city of Naples conducted an assessment of trees in the public right-of-way. The assessment was carried out by a Florida Forest Service Urban Forest Strike Team (UFST). The UFST consisted of four arboricultural professionals who were divided into two crews. Each crew had one assessor with the International Society of Arboriculture’s (ISA) Tree Risk Assessment Qualification (TRAQ). All four assessors were ISA-Certified Arborists with previous experience or training in post-disaster tree risk assessment and incident command systems [
32]. The crews located trees using smartphones and the city’s inventory, which included species, trunk diameter 1.4 m above ground (DBH), height, and GPS coordinates in a GIS-based data collection app (Collector, Esri, Redlands, CA, USA).
The crews conducted a level 1 limited visual assessment [
25] of each tree, using a one-year timeframe as the inspection period. The crews performed a walk-by visual inspection (partial or 360-degree) of trees from ground level to identify obvious defects without using any inspection tools. For each tree, crews assessed the three components of a risk rating: likelihood of failure, likelihood of impact, and severity of consequences [
24,
25]. From the components, the crews assigned each tree a damage category corresponding to mitigation options. Damage categories included the following: “n/a”—missing tree, “none”—no damage present, “minor”—minimal corrective pruning needed, “significant”—major corrective pruning needed, and “whole tree”—severely damaged crown or whole-tree failure, requiring tree removal.
From the population of assessed trees (n = 5094 individuals; n = 38 species), we excluded any species with fewer than 30 individuals, paring the sample to n = 4194 individuals of 15 species. We used descriptive statistics to describe the proportion of individuals in each species and the range of individual DBHs. We used ordinal logistic regression to investigate whether measured parameters (species, DBH, height, presence of utility lines) influenced the proportion of individuals in each damage category. We considered the presence of utility lines a surrogate for pruning, assuming that individuals near utility lines would, on average, be pruned more frequently than individuals away from utility lines. We computed survivorship as the proportion of each species that remained standing following the hurricane. We used SPSS (v 26.0, IBM Corporation, Armonk, NY, USA) for ordinal regression models and Excel (Microsoft Corporation, Redmond, Washington, DC, USA) to compute odds ratios and descriptive statistics. We made statistical inferences at a 0.05 level of significance.
3. Results
Table 1 lists 15 species with at least 30 individuals assessed after Hurricane Irma in Naples, including the number of individuals assessed and each species’ proportion of the total sample. Nearly half of all individuals were
Swietenia mahogani (L.) Jacq., and more than 70% of the sample was composed of three species:
S. mahogani,
Ficus retusa L., and
Quercus virginiana Mill. In contrast, individuals of the eight least-common species made up less than 10% of the sample (
Table 1).
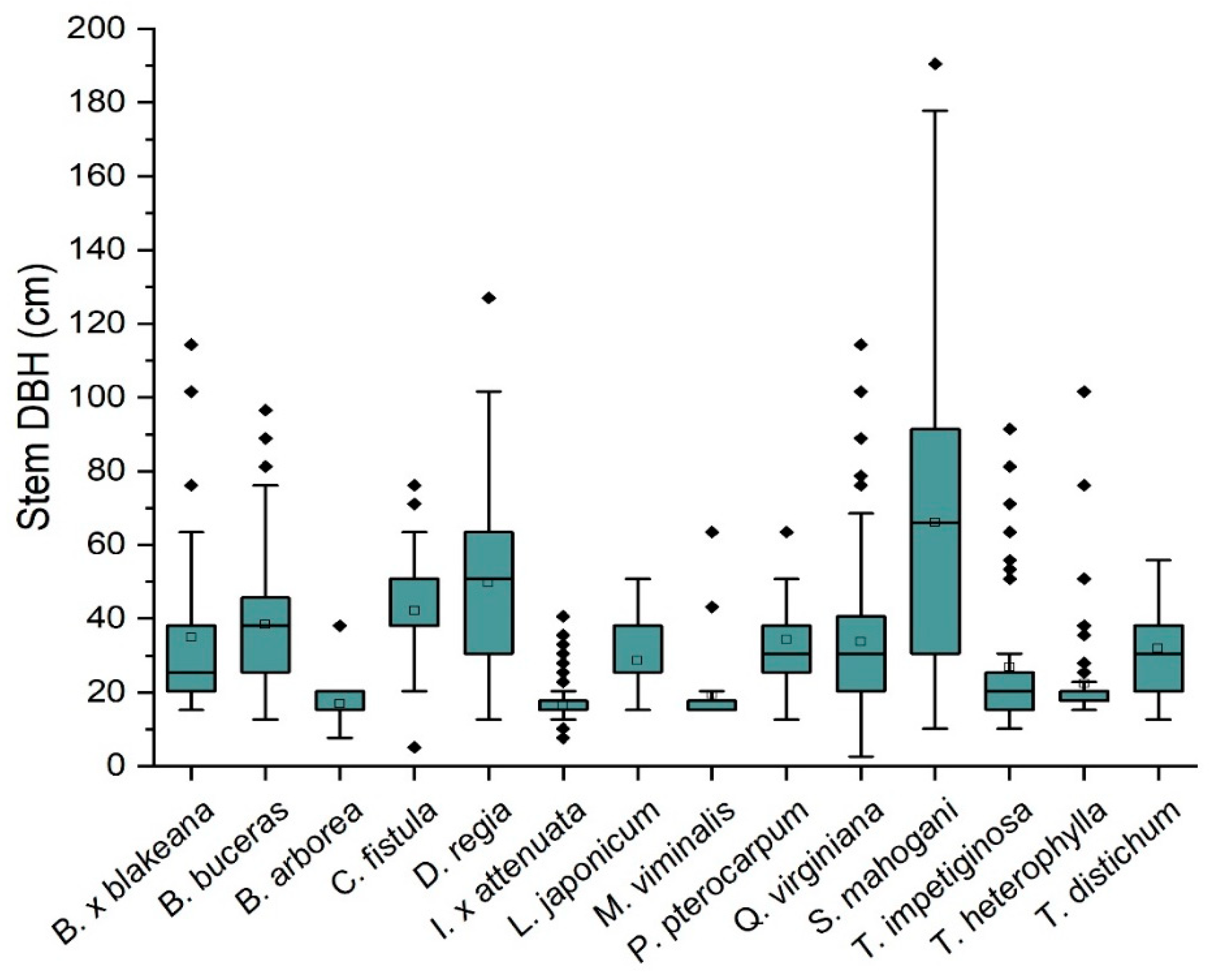
Figure 1 shows the distribution of DBH for each species, except
F.
retusa, in
Table 1. We did not include
F.
retusa in the latter analyses because of its habit of growing aerial roots, which created uniquely large DBH values. The median DBH for species ranged from 15.2 cm for
Bulnesia arborea (Jacq.) Engl.,
Ilex × attenuata Ashe, and
Melaleuca viminalis (Sol. ex Gaertn.) Byrnes. to 66.0 cm for
S. mahogani. The interquartile range of DBH was greatest for
S. mahogani (61.0 cm), nearly twice the range of the species with the next largest value (34.3 cm),
Delonix regia (Bojer ex Hook.) Raf. Species, with the smallest interquartile range (2.5 cm) found for the species
M.
viminalis,
I.
× attenuata, and
Tabebuia heterophylla (DC.) Britton.
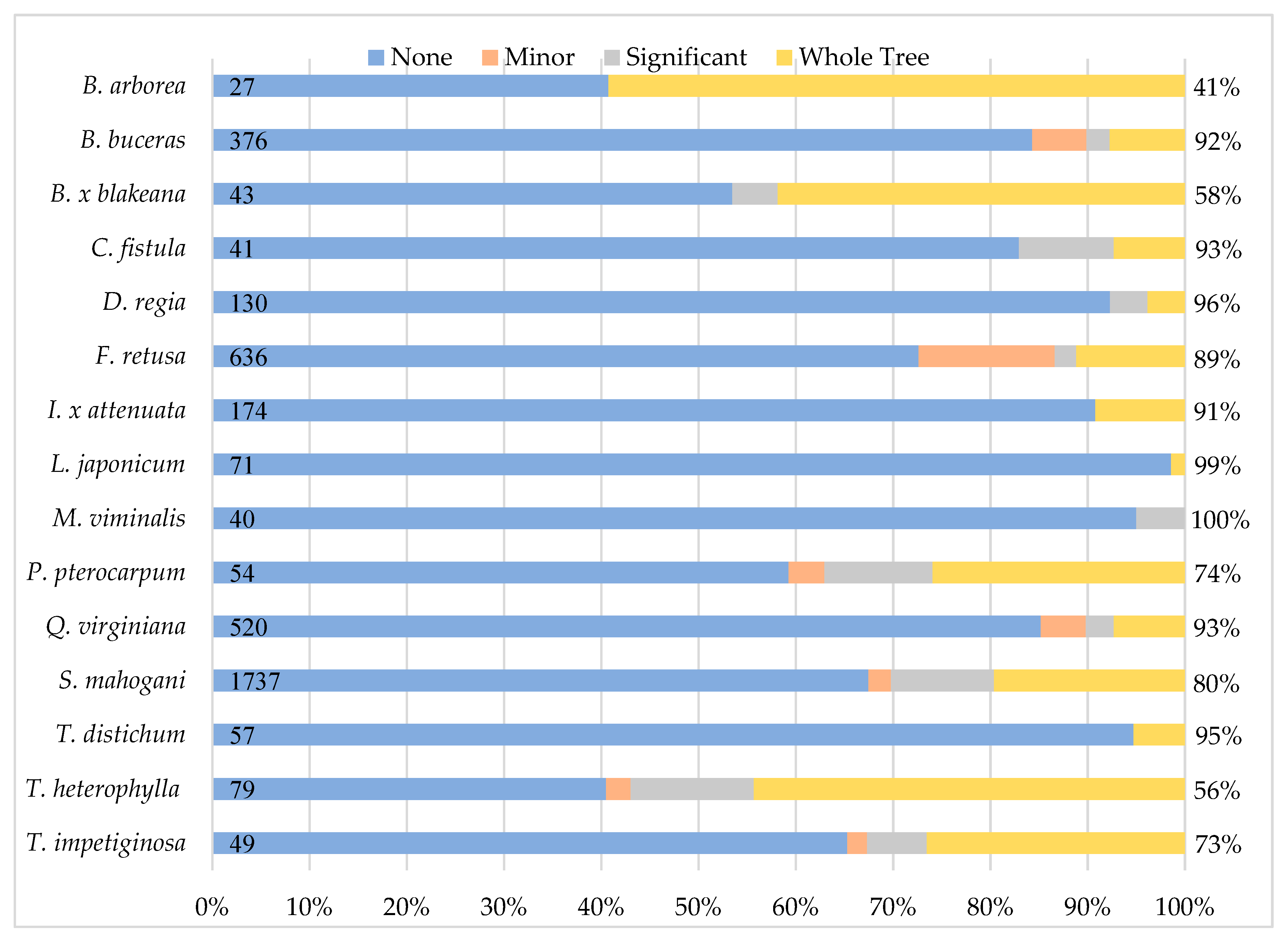
Of 4194 individuals assessed before Hurricane Irma, 162 were not found during the post-hurricane assessments, and we excluded them from further analysis. Of the remaining 4034 individuals, 74% sustained no damage, 4% sustained only minor damage, 6% sustained significant damage and 15% were whole-tree failures or individuals with sufficient crown damage to justify whole tree removal. Survivorship and the proportion of individuals in each damage category varied among species (
Figure 2). More than 90% of individuals of eight species (
Bucida buceras L.,
Cassia fistula L.,
D.
regia,
I.
× attenuata,
Ligustrum japonicum Thunb.,
M.
viminalis,
Q.
virginiana,
Taxodium distichum L.) survived the hurricane. In contrast, less than 60% of individuals of three species (
B.
arborea,
Bauhinia × blakeana Dunn,
T.
heterophylla) survived the hurricane.
Species differences also existed with respect to the proportion of individuals in each damage category (
Figure 2). For five species (
D.
regia,
I.
× attenuata,
L.
japonicum,
M.
viminalis,
T.
distichum), more than 90% of individuals sustained no damage. For an additional four species (
B.
buceras,
C.
fistula,
F.
retusa,
Q.
virginiana), more than 80% of individuals sustained no or minor damage. The remaining six species fared worse. For
B.
arborea and
T.
heterophylla, more than 50% of individuals sustained significant damage or whole-tree failure; for the remaining four species (
B.
blakeana,
Peltophorum pterocarpum (DC.) Backer ex K.Heyne,
S.
mahogany,
Tabebuia impetiginosa (Mart. ex DC.) Standl.), at least 30% of individuals sustained significant damage or whole-tree failure.
Table 2 includes the output from the ordinal logistic regression. We excluded
F.
retusa from the model because of its uniquely large DBH values and used
Q.
virginiana as the comparison species because previous studies have shown it to be a storm-resistant species [
5,
11,
12]. Species, DBH, and the presence of utility lines influenced the proportion of individuals in each damage category. For each cm increase in DBH, the odds of an individual moving to a lesser damage category increased by a factor of 1.01. Similarly, individuals near utility lines were 9.09 times more likely to be in a lower damage category than individuals not near a utility line. Compared to
Q.
virginiana, individuals of six species (
B.
× blakeana, B.
arborea, P.
pterocarpum, S.
mahogany, T.
heterophylla, and
T.
impetiginosa) were significantly more likely to be in a higher damage category, and individuals of two species (
I.
× attenuata,
L.
japonicum) were significantly more likely to be in a lower damage category. There was weaker evidence (
p = 0.07) that
M.
viminalis and
T.
distichum were more likely to be in a lower damage category than individuals of
Q.
virginiana.
4. Discussion
Hurricanes and other catastrophic wind events will continue to threaten urban forests. Because most climate models predict the increasing frequency and severity of such storms—especially in coastal regions like the Atlantic and Gulf Coasts in the United States—the economic and human costs associated with storm-induced tree failures will likely increase in the future. At the same time, however, communities and homeowners continue to plant trees to capitalize on the many benefits they provide to residents. Understanding the factors that influence the likelihood of failure and the degree of damage sustained by amenity trees is integral to sustainable communities.
Our work considered two intrinsic factors that influence the likelihood of tree failure (species and size), as well as the presence of utility lines, which we assumed reflected the effect of pruning. However, we did not consider the effect of structural defects, adding uncertainty to our findings, even though presence/absence was not a factor in [
9]; nor did we investigate the effect of extrinsic factors such as wind speed. A careful spatial analysis would help to correlate local wind speed data with the likelihood of failure or damage, but a more comprehensive sample will be necessary to include other relevant factors like species, size, previous maintenance, and presence/severity of structural defects.
We expected species to influence the proportion of individuals in each damage category because many previous studies have shown differences in survivorship between species in forests and plantations [
1,
16,
20,
33,
34], as well as urban forests [
3,
4,
6,
7,
10,
12]. Our data include open-grown individuals of eight species (
B.
arborea,
B.
× blakeana,
C.
Fistula,
F.
retusa,
I.
× attenuata,
L.
japonicum,
P.
pterocarpum,
T.
impetiginosa) not previously studied, and seven species (
B.
buceras,
D.
regia, M.
viminalis, Q.
virginiana,
S.
mahogani,
T.
distichum,
T.
heterophylla) included in previous studies [
5,
11,
12]. Consistent with previous regional studies of open-grown trees [
5,
7,
11,
12], our data revealed species differences, although we cannot rule out the possibility that such differences were the result of differences in unmeasured variables, such as the presence of defects.
Categorizing individuals in terms of damage—in addition to survivorship—was important, because it may help communities prepare not only for the proportion of trees that may be removed following a storm, but also the volume of debris that needs to be managed. Measuring survivorship alone will likely underestimate the volume of debris that accumulates from individuals that sustained significant damage but remained standing and would not be removed following a storm.
Despite this limitation, we computed survivorship to more readily compare our data with previous studies that reported survivorship of open-grown species from seven previous hurricanes in the region [
5,
11,
12].
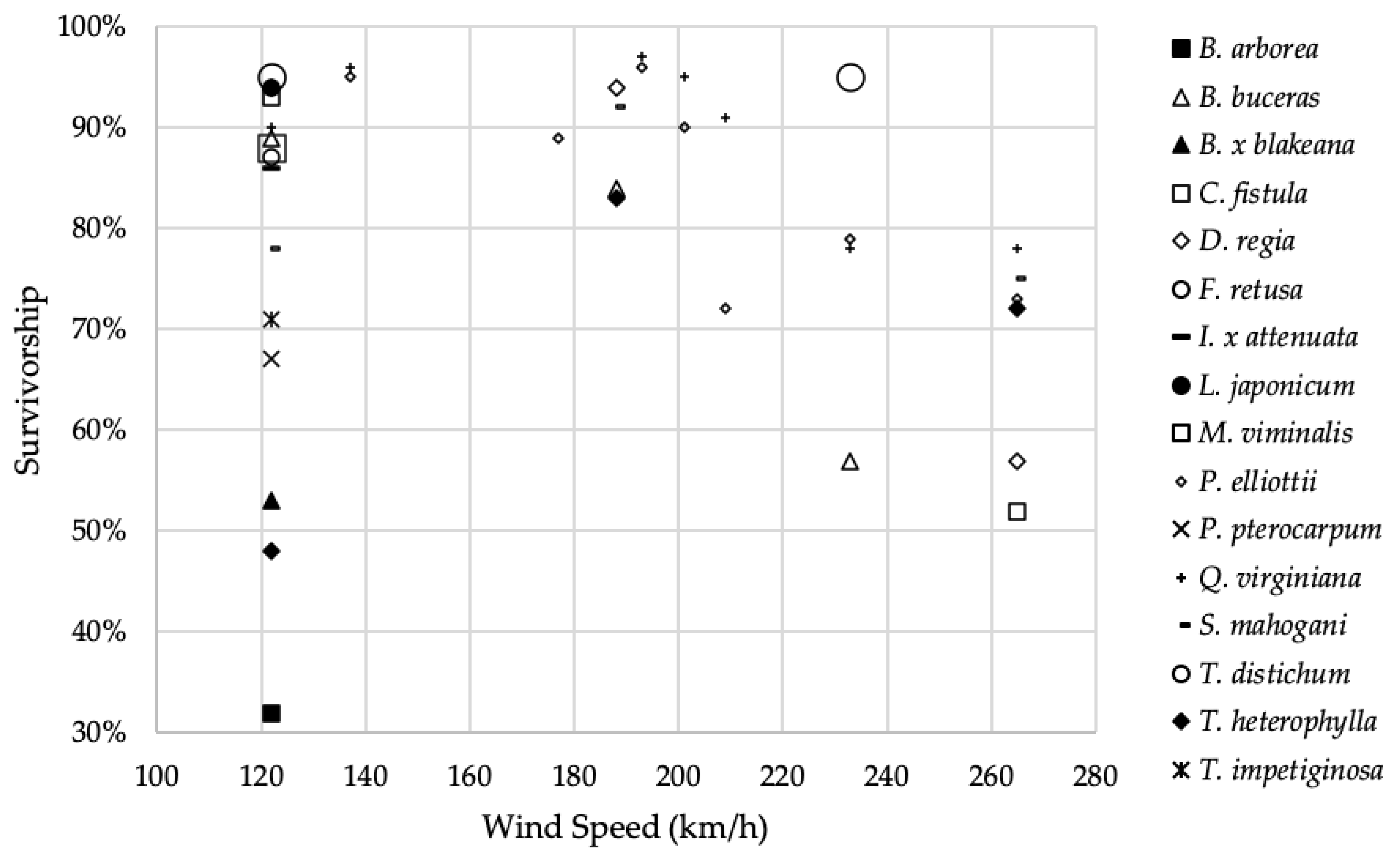
Figure 3 shows survivorship by species for all eight hurricanes, ranging in sustained wind speed from 122 km/h to 265 km/h. Generally, there is a decrease in survivorship in the reported species as wind speed increases. For example, the survivorship of
B.
buceras was noticeably higher following Hurricane Irma (122 km/h) than Hurricane Charley (233 km/h). A similar overall trend can be noted among the seven data sets featuring
Pinus elliottii Engelm. In light of this, variability among species within each hurricane and within a species among hurricanes is evident. For example, the survivorship of
T.
heterophylla was noticeably lower following Hurricane Irma (122 km/h) than hurricanes Andrew (265 km/h) or Georges (188 km/h). The survivorship of
Q.
virginiana, with a reputation among arborists and urban foresters as a particularly resilient species, was high throughout the range of wind speeds. Similarly,
T.
distichum survivorship was nearly identical for Hurricanes Irma and Charley, despite their differing intensities. Such disparities underscore the inherent variability of survivorship, which depends on many intrinsic and extrinsic factors.
Inherent variability was also evident with the effect of DBH on the proportion of individuals in each damage category. Previous studies from the region [
5,
7,
11] and of other sub-tropical [
6] and temperate climates [
1,
3] have shown that the likelihood of failure or the severity of damage is greater for larger trees. Following Hurricane Irma, however, larger trees in Naples were less likely to have sustained greater damage. It was unclear why this occurred, although it was possible that there was a confounding interaction between species and DBH. It is also possible that the effect was an artifact of the damage rating system: the relative severity of a given limb falling decreased with increased crown size and complexity. Although the effect of DBH was statistically significant, the odds ratio was comparatively small. Applying the odds ratio in practice, a tree would have to be 75 cm smaller in DBH to have twice the likelihood of being in the next highest damage category.
Consistent with a previous study in Florida which indicated that pruned trees were less likely to sustain damage [
5], trees near utility lines in Naples were less likely to be in a more severe damage category. We assumed that trees near utility lines were more likely to have been pruned than trees not in proximity to utility lines, which would remove defective branches from parts of the crown near the lines. Repeated pruning may also have reduced the crown area which, in turn, would reduce the wind-induced bending moment [
19].
5. Conclusions
Our findings contribute to a growing body of knowledge that describes the likelihood of failure and the severity of damage to open-grown trees in subtropical climates. However, even in a hurricane-prone region such as Florida, which has provided many opportunities to study wind-induced tree failure, a more comprehensive meta-analysis would be necessary to investigate the interactions between intrinsic (size, species, presence of defects, tree maintenance history, and maintenance of infrastructure like sidewalks and roads that often damages roots and induces windthrow) and extrinsic (wind speed, sheltering, soil texture and volume) factors that influence wind-induced tree failures and damage. Such work is challenging, however. Unlike forest or plantation settings, wind-induced damage in residential settings must be cleaned up relatively quickly, often precluding the collection of detailed measurements of individual trees.
Species selection is an important aspect of urban forest management; in communities that experience regular and severe storms, arborists and urban foresters should consider which species have a greater likelihood of failure or damage. However, restricting selection to species that have consistently demonstrated storm resilience would unnecessarily limit species diversity. Maintaining a resilient and diverse urban forest will require additional knowledge of species performance in storms. It will also require careful planning to provide shelter for less-resilient species and sound arboricultural practice to reduce wind loads and minimize the likelihood of defects that compromise structural integrity.

 ,
, 



{kind=link}
{kind=link}
{kind=link}