The Structure of Northern Siberian Spruce–Scots Pine Forests at Different Stages of Post-Fire Succession

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Stand and Lower Layers Sampling
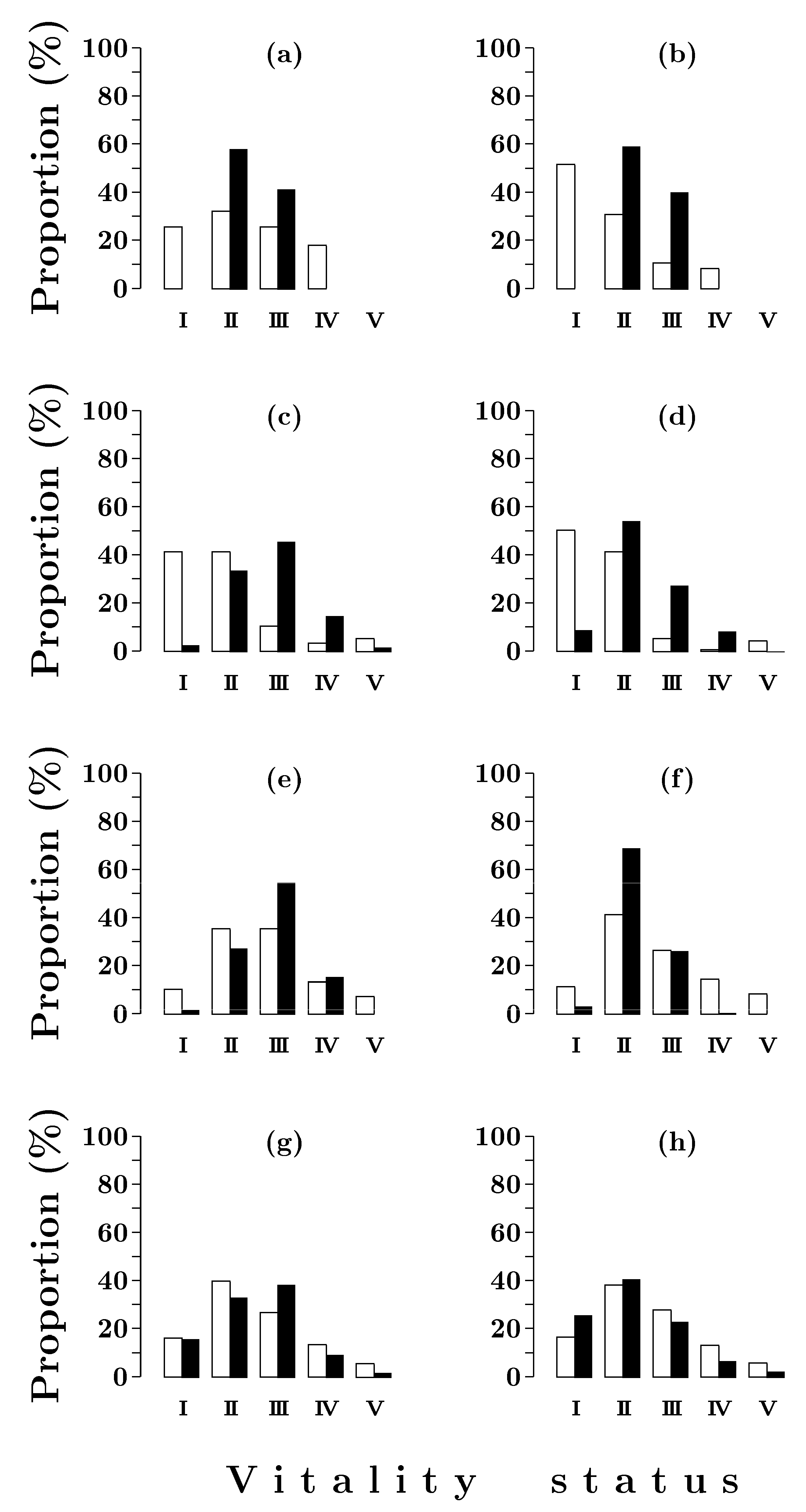
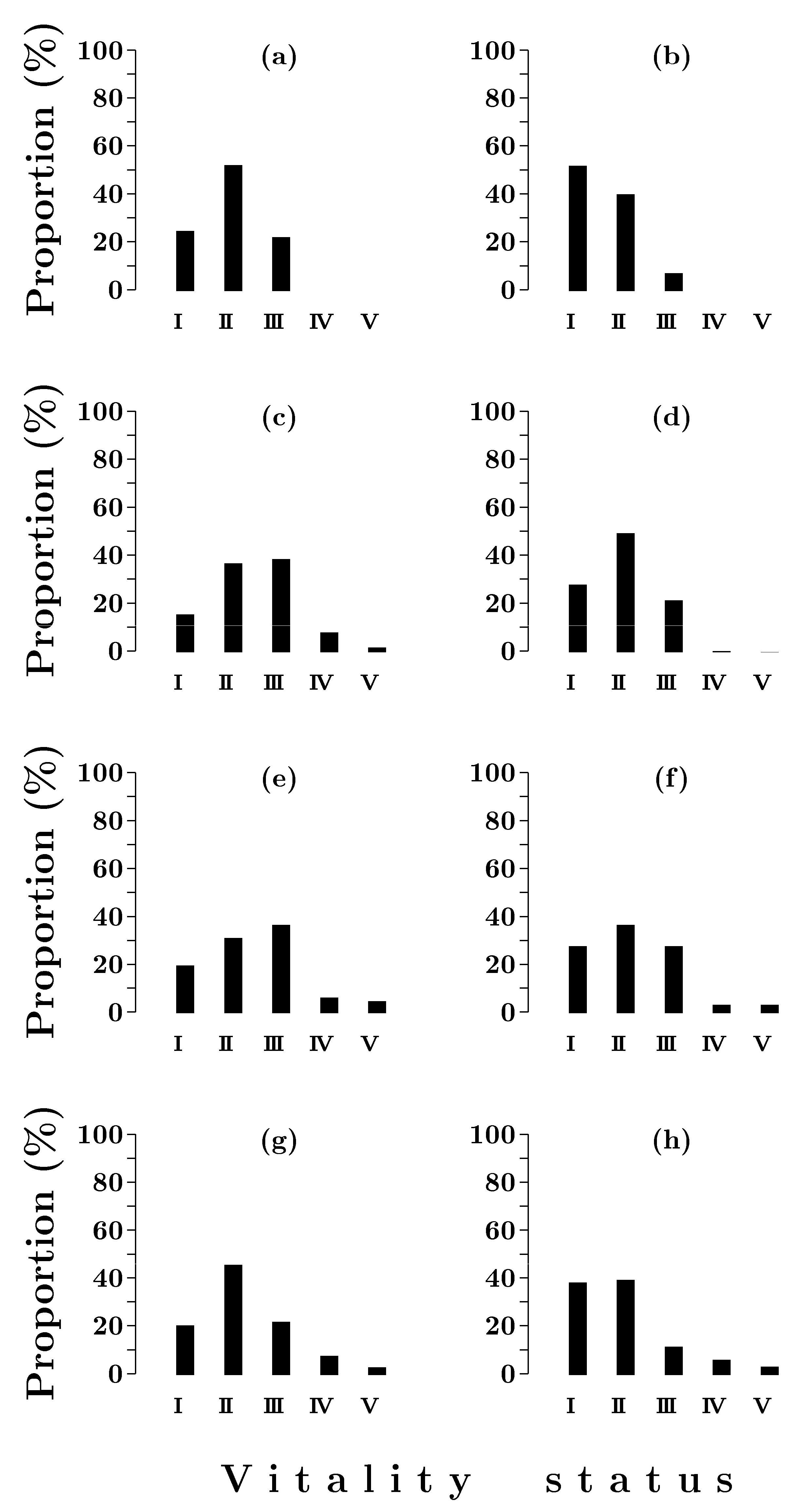
2.3. Tree Vitality Classification
2.4. Stand Vitality Index
2.5. Pielou’s Evenness Index for the Ground Vegetation
2.6. Statistical Analysis
3. Results
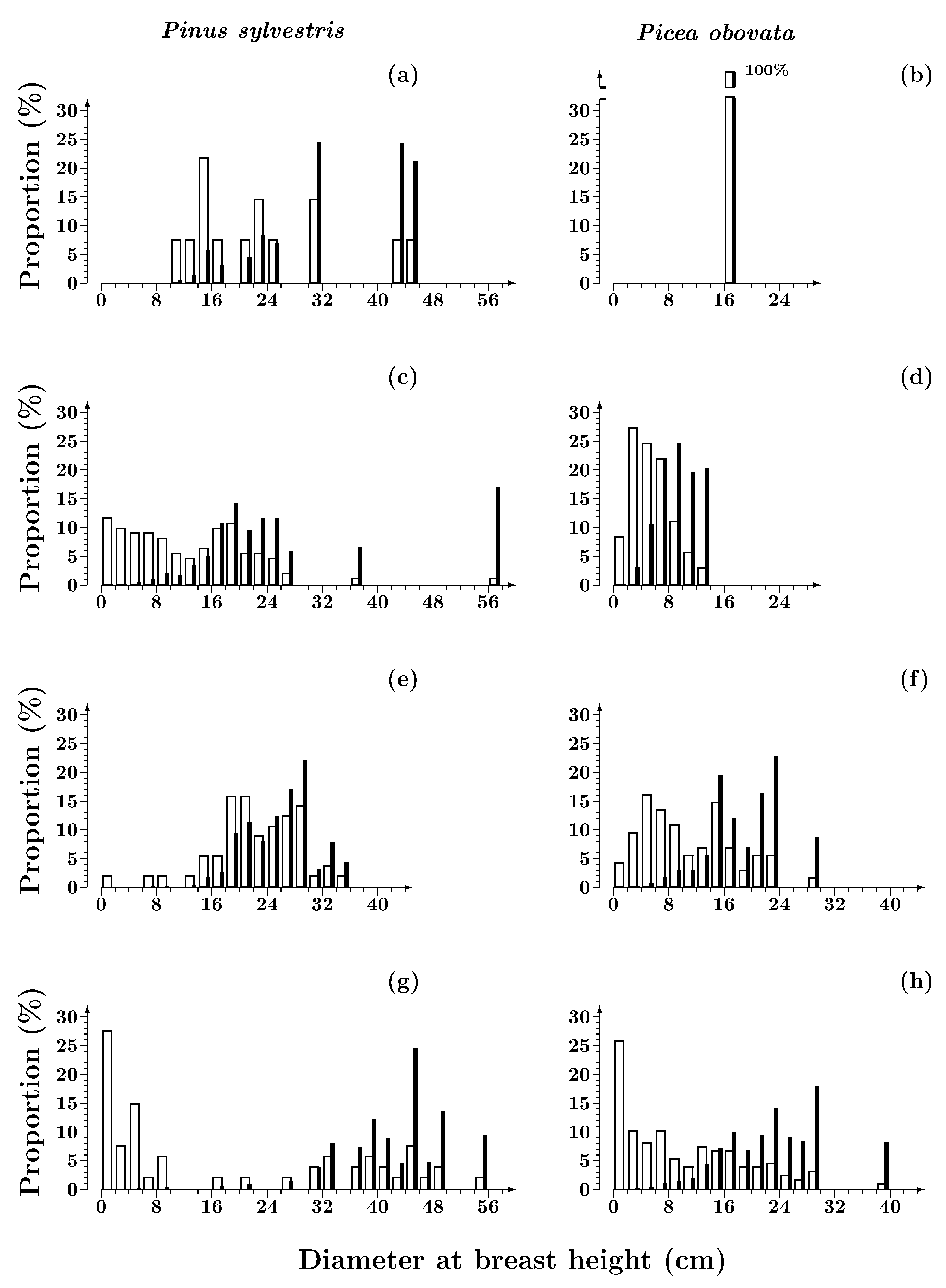
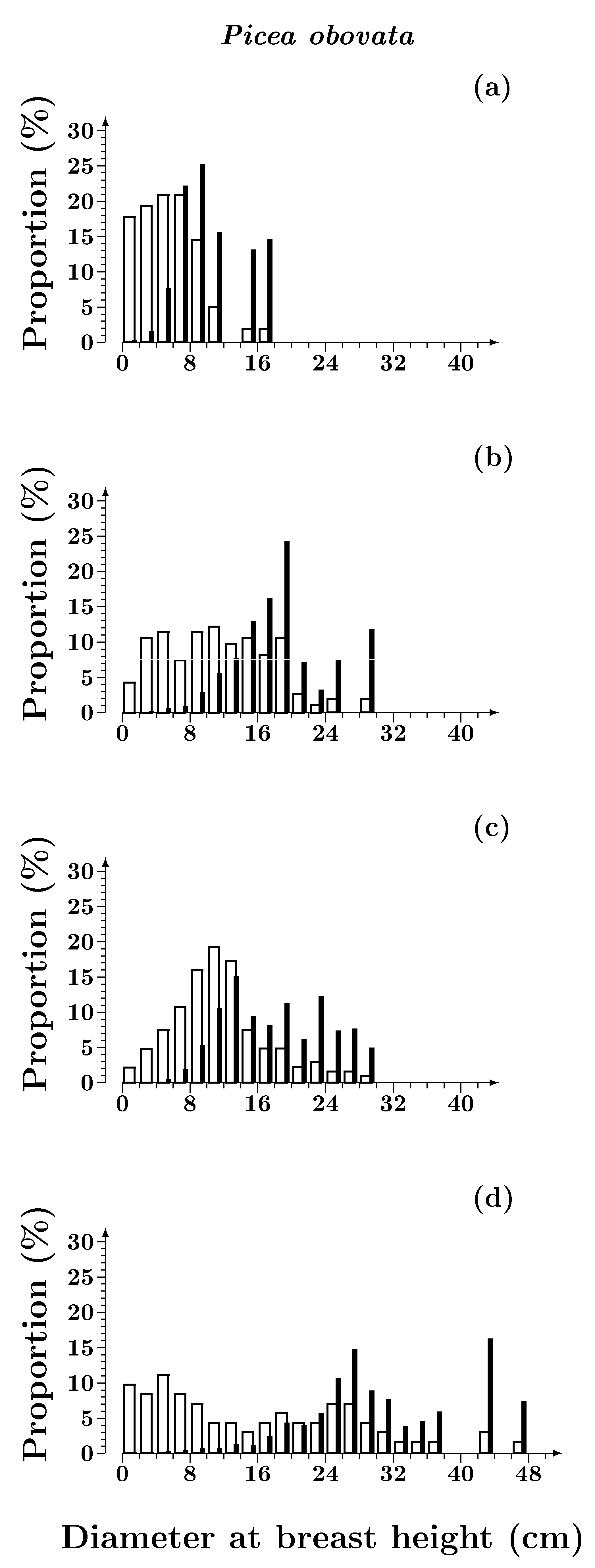
3.1. Structure of Scots Pine and Siberian Spruce Stands
3.1.1. Size Structure
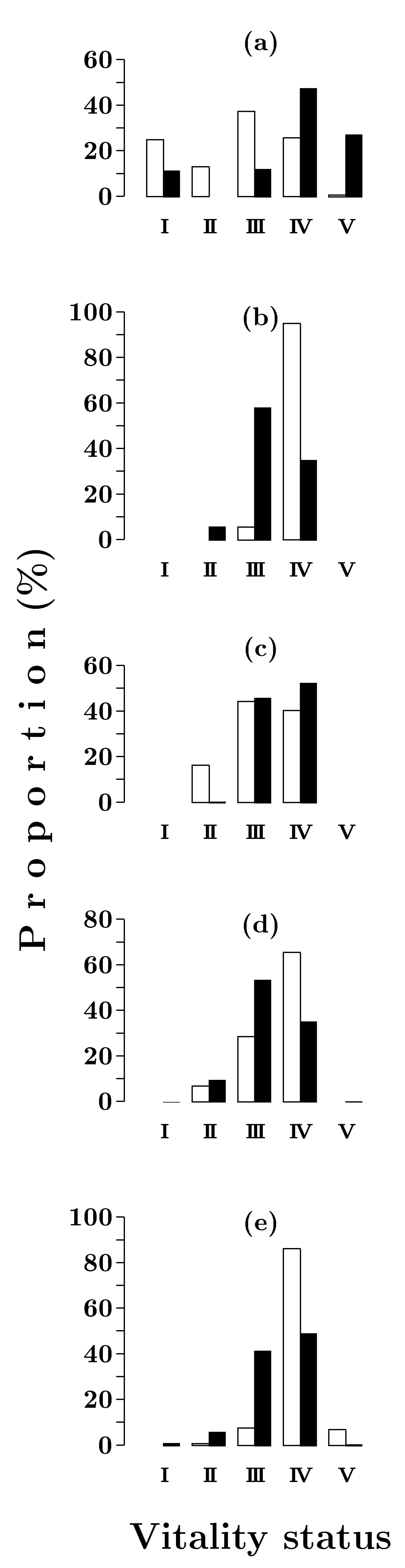
3.1.2. Vitality Structure
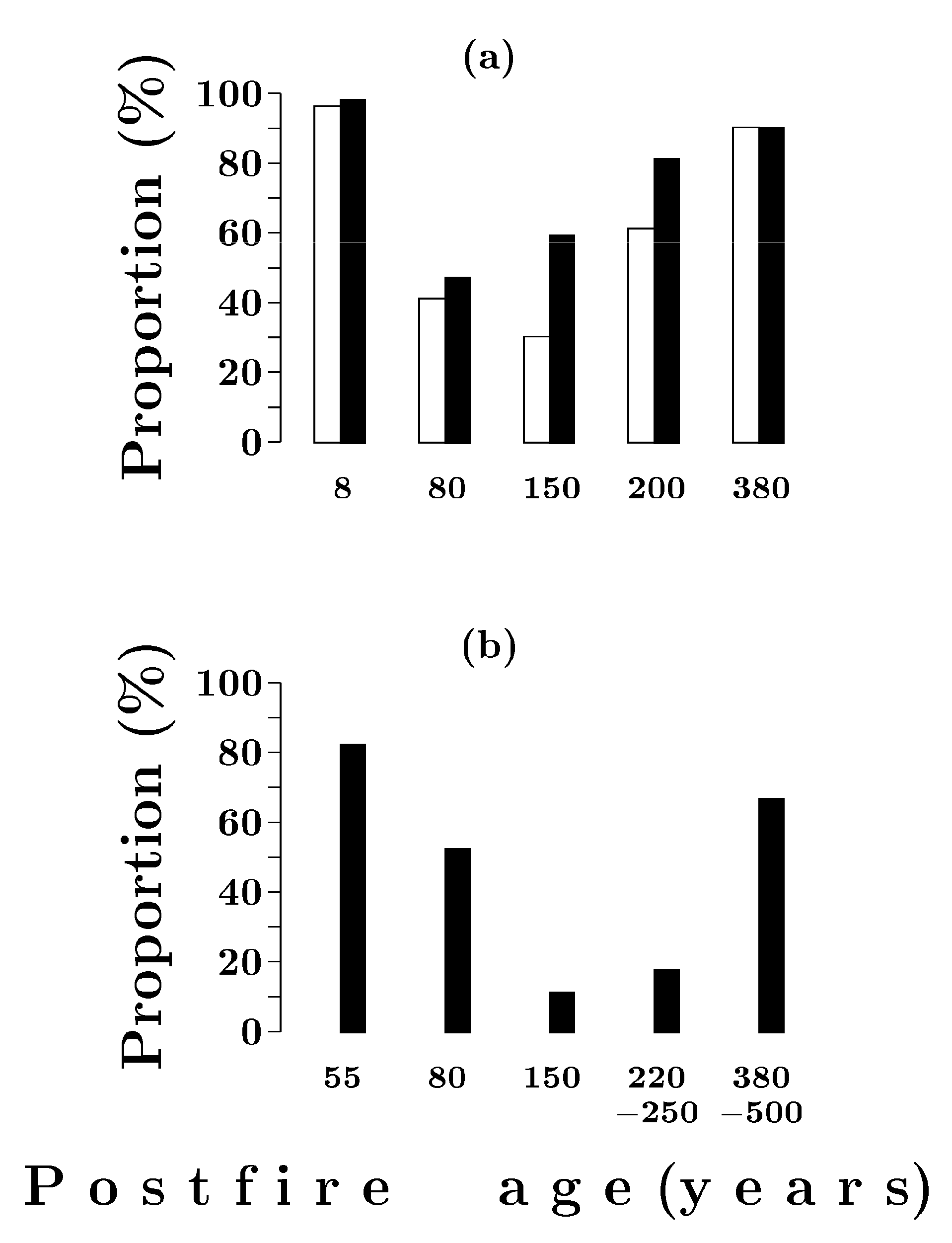
3.2. Natural Regeneration of Scots Pine and Siberian Spruce
3.3. Species Composition of the Ground Cover
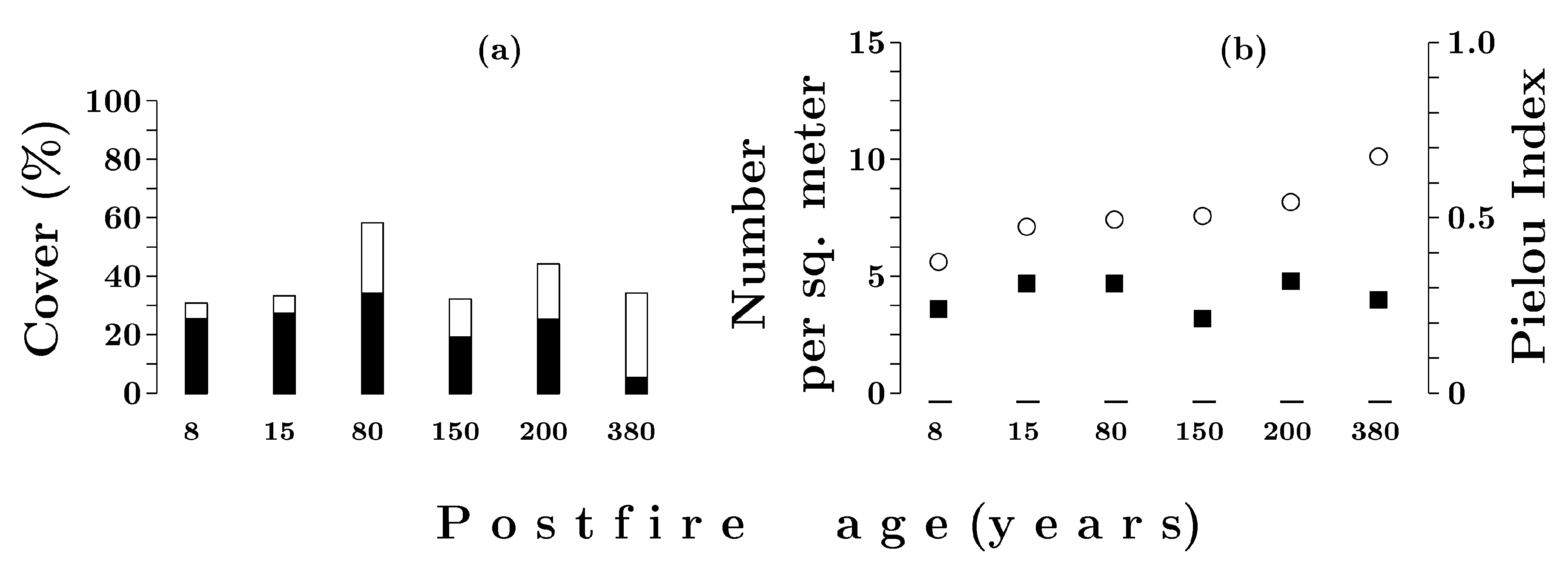
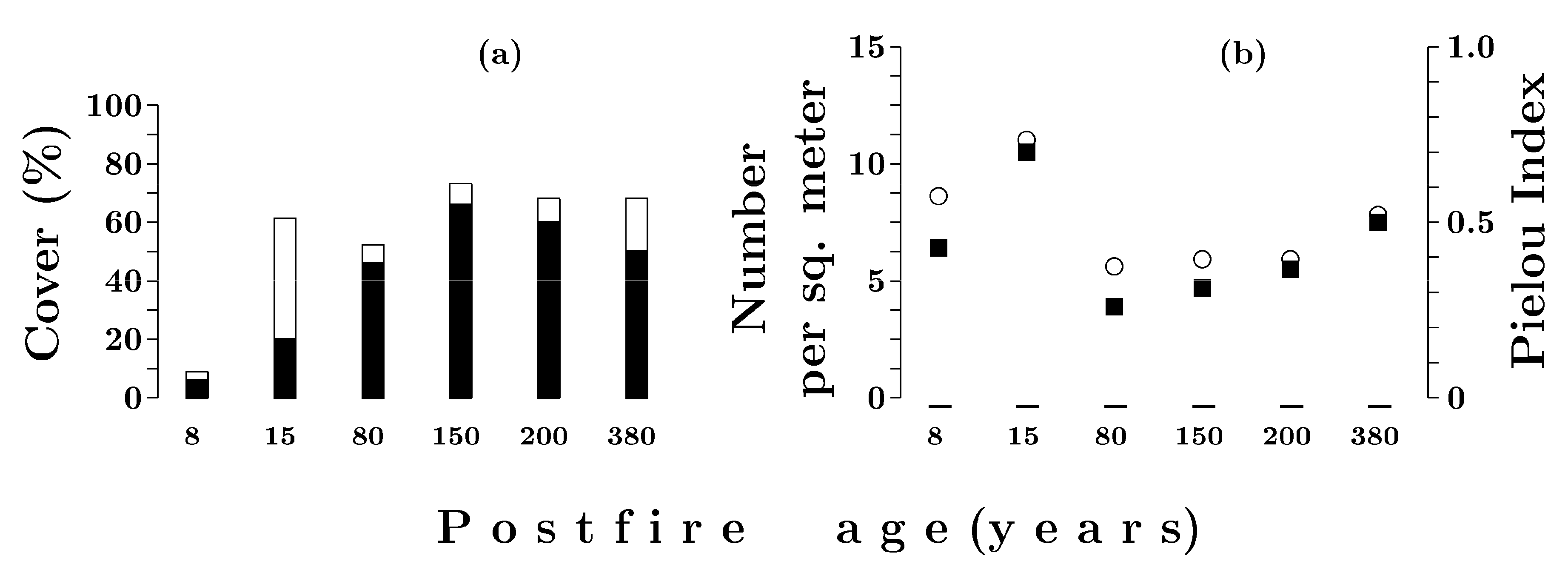
3.3.1. Dwarf Shrub and Herb Layer
3.3.2. Moss and Lichen Layer
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Esseen, P.-A.; Ehnström, B.; Ericson, L.; Sjöberg, K. Boreal forests. Ecol. Bull. 1997, 46, 16–47. [Google Scholar]
- Gauthier, S.; Bernier, P.; Kuuluvainen, T.; Shvidenko, A.Z.; Schepaschenko, D.G. Boreal forest health and global change. Science 2015, 349, 819–822. [Google Scholar] [CrossRef] [PubMed]
- Kuuluvainen, T.; Wallenius, T.H.; Kauhanen, H.; Aakala, T.; Mikkola, K.; Demidova, N.; Ogibin, B. Episodic, patchy disturbances characterize an old-growth Picea abies dominated forest landscape in northeastern Europe. For. Ecol. Manag. 2014, 320, 96–103. [Google Scholar] [CrossRef]
- Kuuluvainen, T. Forest management and biodiversity conservation based on natural ecosystem dynamics in northern Europe: The complexity challenge. AMBIO 2009, 38, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Kuuluvainen, T.; Gauthier, S. Young and old forest in the boreal: Critical stages of ecosystem dynamics and management under global change. For. Ecosyst. 2018, 5, 26. [Google Scholar] [CrossRef]
- Zackrisson, O. The influence of forest fires in the North Swedish boreal forest. Oikos 1977, 29, 22–32. [Google Scholar] [CrossRef]
- Angelstam, P.; Andersson, L. Estimates of the needs for forest reserves in Sweden. Scand. J. For. Res. Suppl. 2001, 3, 38–51. [Google Scholar] [CrossRef]
- Gromtsev, A. Natural disturbance dynamics in the boreal forests of European Russia: A review. Silva Fenn. 2002, 36, 41–55. [Google Scholar] [CrossRef] [Green Version]
- Wallenius, T.H.; Kauhanen, H.H.; Herva, H.; Pennanen, J. Long fire cycle in northern boreal Pinus forests in Finnish Lapland. Can. J. For. Res. 2010, 40, 2027–2035. [Google Scholar] [CrossRef]
- Goldammer, J.G.; Furyaev, V.V. (Eds.) Fire in Ecosystems of Boreal Eurasia; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; 524p. [Google Scholar] [CrossRef]
- Korovin, G.N. Analysis of the distribution of forest fires in Russia. In Fire in Ecosystems of Boreal Eurasia; Goldammer, J.G., Furyaev, V.V., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; pp. 112–128. [Google Scholar]
- Linder, P.; Elfving, B.; Zackrisson, O. Stand structure and successional trends in virgin boreal forest reserves in Sweden. For. Ecol. Manag. 1997, 98, 17–33. [Google Scholar] [CrossRef]
- Boulanger, Y.; Gauthier, S.; Burton, P.J. A refinement of models projecting future Canadian fire regimes using homogeneous fire regime zones. Can. J. For. Res. 2014, 44, 365–376. [Google Scholar] [CrossRef]
- Harper, K.A.; Bergeron, Y.; Gauthier, S.; Drapeau, P. Post-fire development of canopy structure and composition in black spruce forests of Abitibi, Québec: A landscape scale study. Silva Fenn. 2002, 36, 249–263. [Google Scholar] [CrossRef] [Green Version]
- Auvinen, A.-P.; Hildén, M.; Toivonen, H.; Primmer, E.; Niemelä, J.; Aapala, K.; Bäck, S.; Härmä, P.; Ikävalko, J.; Järvenpää, E.; et al. Evaluation of the Finnish National Biodiversity Action Plan 1997–2005. Monographs of Boreal Environment Research; Finnish Environment Institute: Helsinki, Finland, 2007; pp. 1–55. [Google Scholar]
- Bergeron, Y.; Fent, N.J. Boreal forests of eastern Canada revisited: Old growth, nonfire disturbances, forest succession, and biodiversity. Botany 2012, 90, 509–523. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Siitonen, J. Fennoscandian boreal forests as complex adaptive systems. Properties, management challenges and opportunities. In Managing Forests as Complex Adaptive Systems. Building Resilience to the Challenge of Global Change; Messier, C., Puettmaan, K.J., Coates, K.D., Eds.; Earthscan: New York, NY, USA, 2013; pp. 244–268. [Google Scholar]
- Stavrova, N.I.; Gorshkov, V.V.; Katyutin, P.N. Structure formation of forest tree species coenopopulations during post-fire recovery of northern taiga forest. Trans. Karelian Res. Cent. Russ. Acad. Sci. Ser. Biogeogr. 2016, 3, 10–28, (In Russian with English Summary). [Google Scholar]
- Korchagin, A.A. The impact of fires on vegetation and vegetation recovery after fires in the European North. Proc. Bot. Inst. USSR Acad. Sci. Ser. III 1954, 9, 75–149. (In Russian) [Google Scholar]
- Sirén, G. The development of spruce forest on raw humus sites in Northern Finland and its ecology. Acta For. Fenn. 1955, 62. [Google Scholar] [CrossRef] [Green Version]
- Dyrenkov, S.A. Structure and Dynamics of the Boreal Spruce Forest; Nauka: Leningrad, USSR, 1984; 174p. (In Russian) [Google Scholar]
- Steijlen, I.; Zackrisson, O. Long-term regeneration dynamics and successional trends in northern Swedish coniferous forest stand. Can. J. Bot. 1987, 65, 839–848. [Google Scholar] [CrossRef]
- Ågren, J.; Zackrisson, O. Age and size structure of Pinus sylvestris populations on mires in Central and Northern Sweden. J. Ecol. 1990, 78, 1049–1062. [Google Scholar] [CrossRef]
- Engelmark, O. Early post-fire tree regeneration in a Picea-Vaccinium forest in northern Sweden. J. Veg. Sci. 1993, 4, 791–794. [Google Scholar] [CrossRef]
- Lähde, E.; Laiho, O.; Norokorpi, Y.; Saksa, T. Structure and yield of all-sized and even-sized conifer-dominated stands on fertile sites. Ann. For. Sci. 1994, 51, 97–109. [Google Scholar] [CrossRef] [Green Version]
- Hörnberg, G.; Ohlson, M.; Zackrisson, O. Stand dynamics, regeneration patterns and long-term continuity in boreal old-growth Picea abies swamp-forests. J. Veg. Sci. 1995, 6, 291–298. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Syrjänen, K.; Kalliola, R. Structure of a pristine Picea abies forest in northeastern Europe. J. Veg. Sci. 1998, 9, 563–574. [Google Scholar] [CrossRef]
- Volkov, A.D. Comparative assessment of the ecological role and biological specificity of indigenous and derived forests in the North-West of the taiga zone of Russia. In Proceedings of the Primary Forests of the Taiga Zone of Europe: Current State and Conservation Problems: Mater, International Scientific-Practical Conference, Petrozavodsk, Russia, 6–8 July 1999; pp. 9–16. (In Russian). [Google Scholar]
- Kuuluvainen, T.; Mäki, J.; Karjalainen, L.; Lehtonen, H. Tree age distributions in old-growth forest sites in Vienansalo wilderness, eastern Fennoscandia. Silva Fenn. 2002, 36, 169–184. [Google Scholar] [CrossRef] [Green Version]
- Regel, K. Die Pflanzendecke der Halbinsel Kola. Memories de la Faculte des Sciences de L’Universite de Lithuanie. Tail II. Lapponia Ponoensis und Lapponia Imandrae; L′Universite de Lithuanie: Kaunas, Lithuania, 1927; pp. 164–293. [Google Scholar]
- Nekrasova, T.P. The relationship of pine and spruce in the forests of the Kola Peninsula. In Forests of the Kola Peninsula and Their Renewal; USSR Academy of Sciences: Moscow, USSR, 1961; pp. 63–70. (In Russian) [Google Scholar]
- Nicklasson, M.; Hörnberg, G.; Zackrisson, O. Age structure and disturbance history in boreal Pinus-Picea forest, N. Sweden; Department of Forest Vegetation Ecolology, Swedish University of Agricultural Sciences: Umeaa, Sweden, 1998; 23p. [Google Scholar]
- Stavrova, N.I.; Gorshkov, V.V.; Mishko, A.E. Ontogenesis of Siberian spruce (Picea obovata Ledeb.) in North taiga shrub-green-moss pine-spruce forests. Bot. J. 2017, 102, 163–185, (In Russian with English Summary). [Google Scholar]
- Bebiya, S.M. Differentiation of trees in the forest, their classification and determination of the life state of stands. Lesovedenie 2000, 4, 35–43, (In Russian with English Summary). [Google Scholar]
- Torlopova, N.V.; Ilchukov, S.V. Vital state of native pine forests at the Pechora-Ilych biosphere reserve. Lesovedenie 2003, 3, 34–40, (In Russian with English Summary). [Google Scholar]
- Stavrova, N.I.; Gorshkov, V.V.; Katyutin, P.N. Dynamics of vitality structure of Picea obovata and Betula pubescens coenopopulations during post fire succession in northern taiga spruce forest. Botanicheskii Zhurnal 2003, 95, 1550–1565, (In Russian with English Summary). [Google Scholar]
- World Reference Base for Soil Resources 2014, International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Report No. 106; Food and Agriculture Organization: Rome, Italy, 2015.
- Pereverzev, V.N. Forest Soils of the Kola Peninsula; Nauka: Moscow, Russia, 2004; 232p, (In Russian with English Summary). [Google Scholar]
- Storozhenko, V.G. Dating of decomposition of fir windfall. Ekologiya 1990, 6, 66–69, (In Russian with English Summary). [Google Scholar]
- Alekseev, V.A. Some problems of diagnosis and classification of forest ecosystems damaged by air pollution. In Forest Ecosystems and Atmospheric Pollution; Alekseev, V.A., Ed.; Nauka: Leningrad, USSR, 1990; pp. 38–54. [Google Scholar]
- Yarmishko, V.T.; Gorshkov, V.V.; Stavrova, N.I. Vitality structure of Pinus sylvestris L. in forest communities with varying degrees and types of anthropogenic disturbance. Plant Resour. 2003, 39, 1–19, (In Russian with English Summary). [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing Company: Oxford, UK, 2004; 256p. [Google Scholar]
- Bailey, R.L.; Dell, T.R. Quantifying diameter distributions with Weibull function. For. Sci. 1973, 19, 97–104. [Google Scholar]
- Maltamo, M.; Kangasb, A.; Uutterac, J.; Torniainenc, T.; Saramäki, J. Comparison of percentile based prediction methods and the Weibull distribution in describing the diameter distribution of heterogeneous Scots pine stands. For. Ecol. Manag. 2000, 133, 263–274. [Google Scholar] [CrossRef]
- Mulverhill, C.; Coops, N.C.; White, J.C.; Tompalski, P.; Marshall, P.L.; Bailey, T. Enhancing the estimation of stem-size distributions for unimodal and bimodal stands in a boreal mixedwood forest with airborne laser scanning data. Forests 2018, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Cao, L.; Mulverhill, C.; Liu, H.; Pang, Y.; Li, Z. Prediction of Diameter Distributions with Multimodal Models Using LiDAR Data in Subtropical Planted Forests. Forests 2019, 10, 125. [Google Scholar] [CrossRef] [Green Version]
- Melekhov, I.S. The Impact of Fires on Forest; State Forestry Publishing House: Moscow: Leningrad, Russia, 1948; 126p. (In Russian) [Google Scholar]
- Sannikov, S.N.; Sannikova, N.S.; Petrova, I.V. Outlines of Theory of Forest Populational Biology; Ural Branch of the Russian Academy of Sciences: Ekaterinburg, Russia, 2012; 272p, (In Russian with English Summary). [Google Scholar]
- Steijlen, I.; Nilsson, M.-C.; Zackrisson, O. Seed regeneration of Scots pine in boreal forests stands dominated by lichen and feather moss. Can. J. For. Res. 1995, 25, 713–723. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Juntunen, P. Propeties and importance of tree regeneration microhabitats in a small windthrow gap in a boreal Pinus sylvestris dominated forest. J. Veg. Sci. 1998, 9, 551–562. [Google Scholar] [CrossRef]
- Nguyen-Xuan, T.; Bergeron, Y.; Simard, D.; Fyles, J.W.; Paré, D. The importance of forest floor disturbance in the early regeneration patterns of the boreal forest of western and Central Quebec: A wildfire versus logging comparison. Can. J. For. Res. 2000, 30, 1353–1364. [Google Scholar] [CrossRef]
- Grenfell, R.; Aakala, T.; Kuuluvainen, T. Microsite occupancy and the spatial structure of understorey regeneration in three late-successional Norway spruce forests in northern Europe. Silva Fenn. 2011, 45, 1093–1110. [Google Scholar] [CrossRef] [Green Version]
- Leemans, R. Canopy gaps and establishment patterns of spruce (Picea abies L. Karst.) in two old-growth coniferous forests in central Sweden. Vegetatio 1991, 93, 157–165. [Google Scholar] [CrossRef]
- Korotkov, V.N.; Morozov, A.S.; Yaroshenko, A.Y. Mosaic organization and spontaneous dynamics of quasiclimax boreal forests. In Eastern European Forests: History in the Holocene and Modernity; Smirnova, O.V., Ed.; Nauka: Moscow, Russia, 2004; Volume 2, pp. 330–346. (In Russian) [Google Scholar]
- Smirnova, O.V.; Toropova, N.A. Succession and climax as the ecosystem process. Biol. Bull. Rev. 2008, 128, 129–144, (In Russian with English Summary). [Google Scholar]
- Kuuluvainen, T. Gap disturbance, ground microtopography, and the regeneration dynamics of boreal forests in Finland: A review. Ann. Zool. Fenn. 1994, 31, 35–51. [Google Scholar]
- Stavrova, N.I.; Kalimova, I.B.; Gorshkov, V.V.; Drozdova, I.V.; Alekseeva-Popova, N.V.; Bakkal, I.Y. Long-term postfire changes of soil characteristics in dark coniferous forests of the European North. Eurasian Soil Sci. 2019, 52, 218–227. [Google Scholar] [CrossRef]
- Sannikov, S.N.; Sannikova, N.S. Ecology of Natural Regeneration of Pine under the Forest Canopy; Gorchakovsky, P.L., Ed.; Nauka: Moscow, USSR, 1985; 149p, (In Russian with English Summary). [Google Scholar]
- Hofgaard, A. 50 years of change in a Swedish boreal old-growth Picea abies forests. J. Veg. Sci. 1993, 4, 773–782. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Kalmari, R. Regeneration microsites of Picea abies seedlings in a windthrow area of a boreal old-growth forests in Finland. Ann. Bot. Fenn. 2003, 40, 401–413. [Google Scholar]
- Juntunen, V.; Neuvonen, S. Natural regeneration of Scots pine and Norway spruce close to the timberline in northern Finland. Silva Fenn. 2006, 40, 443–458. [Google Scholar] [CrossRef] [Green Version]
- Zielonka, T. When does dead wood turn into a substrate for spruce replacement? J. Veg. Sci. 2006, 17, 739–746. [Google Scholar] [CrossRef]
- Sannikov, S.N.; Sannikova, N.S.; Petrova, I.V. Natural Forest Regeneration in Western Siberia (Ecological-Geograpfical Essay); Ural Branch of the Russian Academy of Sciences: Ekaterinburg, Russia, 2004; 198p, (In Russian with English Summary). [Google Scholar]
- Ulanova, N.G. The effect of windthrow on forests at different spatial scales: A review. For. Ecol. Manag. 2000, 135, 155–167. [Google Scholar] [CrossRef]
- Ahlström, M.A.; Lundqvist, L. Stand development during 16–57 years in partially harvested sub-alpine uneven-aged Norway spruce stands reconstructed from increment cores. For. Ecol. Manag. 2015, 350, 81–86. [Google Scholar] [CrossRef]
- Hett, J.M.; Loucks, O.L. Age structure of balsam fir and eastern hemlock. J. Ecol. 1976, 64, 1029–1044. [Google Scholar] [CrossRef]
- McCarthy, J.W.; Weetman, G. Age and size structure of gap dynamic, old-growth boreal forest stand in Newfoundland. Silva Fenn. 2006, 40, 209–230. [Google Scholar] [CrossRef] [Green Version]
- Lundqvist, L. Tamm Review: Selection system reduces long-term volume growth in Fennoscandic uneven-aged Norway spruce forests. For. Ecol. Manag. 2017, 391, 362–375. [Google Scholar] [CrossRef]
- Ushakova, G.I. Biogeocenotic features of spruce and birch trees in rare-coniferous spruce forests of the Far North and their role in migration of elements. In Fertility of Soil and Productivity of Phytocenoses; KSC of the USSR Academy of Sciences: Apatity, USSR, 1991; pp. 55–72. (In Russian) [Google Scholar]
- Orlova, M.A.; Lukina, N.V.; Smirnov, V.E.; Isaeva, L.G. Fertility of soils of birch forests at the northern margin of their distribution. Pochvovedenie 2014, 3, 327–339. [Google Scholar]
- Pereverzev, V.N. Soil Formation on Loose and Crystalline Rocks in Northern Fennoscandia; Kola Scientific Center RAS: Apatity, Russia, 2013; 158p. (In Russian) [Google Scholar]
- Gorshkov, V.V.; Bakkal, I.J. Recovery of the ground layer in spruce (Picea obovata Ledeb.) forests. In Dynamics of Forest Communities in the North-West of Russia; Yarmishko, V.T., Ed.; VVM: St.-Petersburg, Russia, 2009; pp. 197–204. (In Russian) [Google Scholar]
- Gorshkov, V.V.; Bakkal, I.J. Features of post-fire recovery dynamics of communities with domination of lichens. News Samara Sci. Cent. RAS 2012, 14, 1223–1227. (In Russian) [Google Scholar]
- Pushkina, N.M. Regeneration of vegetation on forest fire areas. Works Lapland State Reserve 1960, IV, 5–124, (In Russian with English Summary). [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Site | Time Since Last Fire (Years) | Species | Basal Area (m2 ha−1) | Density (Trees ha−1) | Age (Years) | DBH (cm) | Tree Height (m) |
|---|---|---|---|---|---|---|---|
| L4 | 8 | P * | 3.4 | 79 | 102 | 23.5 | 13.7 |
| S * | 0.14 | 6 | 120 | 17.0 | 10.5 | ||
| B * | 0.1 | 22 | 82 | 7.7 | 5.0 | ||
| L9 | 15 | P * | 5.3 | 50 | No data | 31.4 | 15.0 |
| S * | 1.7 | 33 | No data | 21.5 | 14.0 | ||
| B * | 0.01 | 27 | No data | 6.0 | 5.2 | ||
| L7 | 80 | P * | 1.1 | 13 | 220 | 32.0 | 18.0 |
| P | 11.3 | 587 | 76 | 15.6 | 13.3 | ||
| S | 0.7 | 160 | 58 | 12.3 | 0.7 | ||
| B | 5.5 | 1262 | 77 | 7.4 | 9.1 | ||
| L8 | 150 | P | 16.4 | 387 | 106 | 23.2 | 16.2 |
| S | 6.0 | 433 | 125 | 13.3 | 13.3 | ||
| B | 3.6 | 307 | No data | 12.2 | 10.7 | ||
| L5 | 200 | P * | 3.05 | 20 | 252 | 44.1 | 17.1 |
| P | 7.1 | 173 | 140 | 22.8 | 14.1 | ||
| S | 4.1 | 333 | 135 | 12.5 | 13.7 | ||
| B | 1.5 | 400 | 86 | 6.9 | 6.3 | ||
| L3 | 380 | P | 4.7 | 66 | 306 | 30.1 | 16.8 |
| S | 4.4 | 182 | 215 | 16.4 | 13.9 | ||
| B | 3.3 | 352 | 156 | 11.1 | 9.7 |
| Time Since Last Fire (Years) | N | Mean, cm | S.D. | V, % | Range, cm | S | K | BC |
|---|---|---|---|---|---|---|---|---|
| Scots pine (Siberian spruce–Scots pine forests) | ||||||||
| 8 | 14 * | 23.4 | 10.4 | 45 | 34.0 | No data | No data | No data |
| 80 | 113 | 11.8 | 7.7 | 65 | 27.3 | 0.18 | −1.17 | 0.5399 |
| 150 | 58 | 22.3 | 6.3 | 28 | 32.3 | −0.74 | 1.30 | 0.3466 |
| 200 | 50 | 21.5 | 9.7 | 45 | 41.0 | 0.27 | –0.10 | 0.4588 |
| 380 | 55 | 18.4 | 18.5 | 101 | 53.7 | 0.50 | −1.50 | 0.7466 |
| Siberian spruce (Siberian spruce–Scots pine forests) | ||||||||
| 80 | 62 | 5.4 | 2.8 | 52 | 12.8 | 0.69 | 0.24 | 0.4349 |
| 150 | 76 | 10.8 | 6.5 | 60 | 27.8 | 0.52 | −0.52 | 0.4879 |
| 200 | 179 | 9.2 | 5.7 | 63 | 29.0 | 0.96 | 1.17 | 0.4590 |
| 380 | 141 | 9.9 | 8.6 | 87 | 38.7 | 0.84 | −0.04 | 0.5638 |
| Siberian spruce (Siberian spruce forests) | ||||||||
| 55 | 58 | 5.2 | 3.0 | 56 | 11.0 | 0.35 | –0.84 | 0.4828 |
| 80 | 63 | 5.3 | 3.3 | 63 | 15.3 | 0.91 | 1.11 | 0.4291 |
| 150 | 126 | 11.2 | 6.2 | 56 | 27.6 | 0.37 | –0.45 | 0.4334 |
| 220 | 152 | 11.6 | 5.3 | 45 | 29.4 | 0.86 | 1.04 | 0.4242 |
| 250 | 92 | 11.6 | 7.3 | 63 | 27.8 | 0.24 | –0.94 | 0.4893 |
| 380 | 74 | 15.7 | 11.8 | 75 | 46.1 | 0.60 | –0.50 | 0.5176 |
| 500 | 92 | 10.6 | 9.3 | 88 | 38.0 | 1.19 | 0.84 | 0.6130 |
| Time Since Last Fire (Years) | Chi-Squared Test | Distribution Parameters | ||||
|---|---|---|---|---|---|---|
| (χ2) | d.f. | p-Value | Lower Threshold | Shape | Scale | |
| Scots pine (Siberian spruce–Scots pine forests) | ||||||
| 80 (DBH < 12 cm) | 9.31 | 4 | 0.06 | – | 1.55 | 5.71 |
| 80 (DBH > 12 cm) | 5.14 | 3 | 0.16 | 11.8 | 1.91 | 7.96 |
| 150 | 7.47 | 9 | 0.59 | – | 4.08 | 24.38 |
| 200 | 13.74 | 12 | 0.32 | – | 2.34 | 24.10 |
| 380(DBH > 26 cm) | 1.86 | 4 | 0.86 | 20.5 | 3.13 | 21.63 |
| Siberian spruce (Siberian spruce–Scots pine forests) | ||||||
| 80 | 2.20 | 3 | 0.53 | – | 2.02 | 6.14 |
| 150 | 9.61 | 9 | 0.38 | – | 1.73 | 12.09 |
| 200 | 10.93 | 10 | 0.36 | – | 1.63 | 10.22 |
| 380 | 20.68 | 14 | 0.11 | – | 1.11 | 10.26 |
| 380(DBH > 10 cm) | 3.17 | 7 | 0.87 | 10.4 | 1.38 | 9.24 |
| Siberian spruce (Siberian spruce forests) | ||||||
| 55 | 4.47 | 3 | 0.22 | – | 1.86 | 5.90 |
| 80 | 4.30 | 4 | 0.37 | – | 1.66 | 5.96 |
| 150 | 15.70 | 10 | 0.11 | – | 1.81 | 12.59 |
| 220 | 13.67 | 9 | 0.13 | – | 2.30 | 13.15 |
| 250 | 14.61 | 11 | 0.20 | – | 1.52 | 12.76 |
| 380 | 15.50 | 15 | 0.42 | – | 1.24 | 16.70 |
| 500 | 11.88 | 12 | 0.46 | – | 1.14 | 11.15 |
| Time Since Last Fire (Years) | Pinus Sylvestris | Picea Obovata | ||||
|---|---|---|---|---|---|---|
| Trees (Individuals > 1.3 m High) | Seedlings (Individuals < 1.3 m High) | Trees (Individuals > 1.3 m high) | Seedlings (Individuals < 1.3 m High) | |||
| by Volume | by Number | by Number | By Volume | By Number | by Number | |
| Siberian spruce–Scots pine forest | ||||||
| 8 | – | – | 0.53 | – | – | 0.33 |
| 80 | 0.79 | 0.62 | 0.14 | 0.60 | 0.60 | 0.35 |
| 150 | 0.82 | 0.75 | 0.35 | 0.61 | 0.50 | 0.28 |
| 200 | 0.54 | 0.52 | 0.24 | 0.65 | 0.48 | 0.36 |
| 380 | 0.57 | 0.57 | 0.15 | 0.68 | 0.59 | 0.31 |
| Siberian spruce forest | ||||||
| 80 | – | – | – | 0.85 | 0.72 | 0.19 |
| 150 | – | – | – | 0.71 | 0.59 | 0.19 |
| 220 | – | – | – | 0.72 | 0.59 | 0.16 |
| 380 | – | – | – | 0.73 | 0.63 | 0.35 |
| Species | Time Since Last Fire (Years) | |||||
|---|---|---|---|---|---|---|
| 8 | 15 | 80 | 150 | 200 | 380 | |
| M ± S.E. | M ± S.E. | M ± S.E. | M ± S.E. | M ± S.E. | M ± S.E. | |
| Avenella flexuosa (L.) Drej. | 1.1 ± 0.1 | 0.2 ± 0.05 | 0.3 ± 0.1 | <0.1 | 0.3 ± 0.1 | 0.2 ± 0.05 |
| Calluna vulgaris (L.) Hull | 0.2 ± 0.05 | 1.3 ± 0.3 | 0 | 0 | 0.1 ± 0.1 | <0.1 |
| Empetrum hermaphroditum Hagerup | 0 | 0.5 ± 0.1 | 0.8 ± 0.4 | 4.7± 0.8 | 10.8 ± 0.8 | 17 ± 1 |
| Linnaea borealis L. | 0.5 ± 0.1 | 0 | 0 | 0 | 0.5 ± 0.1 | 0.1 ± 0.1 |
| Lycopodium annotinum L. | 0 | 0 | 0.6 ± 0.3 | 0 | 0 | 0.1 ± 0.1 |
| Melampyrum pratense L. | 0.3 ± 0.05 | 0 | 0.1 ± 0.05 | 0 | 0.2±0.1 | <0.1 |
| Vaccinium myrtillus L | 26 ± 2 | 28 ± 1 | 35 ± 2 | 20 ± 2 | 26 ± 1 | 6.1 ± 0.6 |
| Vaccinium vitis-idaea L. | 2.1 ± 0.5 | 3.0 ± 0.2 | 11 ± 2 | 7 ± 1 | 4.9 ± 0.7 | 8.6 ± 0.8 |
| Vaccinium uliginosum L. | 0.2 ± 0.05 | 0.2 ± 0.2 | 11 ± 2 | 0.3 ± 0.2 | 0.5 ± 0.3 | 0.3 ± 0.1 |
| Species | Time Since Last Fire (Years) | |||||
|---|---|---|---|---|---|---|
| 8 | 15 | 80 | 150 | 200 | 380 | |
| M ± S.E. | M ± S.E. | M ± S.E. | M ± S.E. | M ± S.E. | M ± S.E. | |
| Cladonia cornuta (L.) Hoffm. | 0 | 1.0 ± 0.5 | 0 | <0.1 | <0.1 | 0.1 ± 0.1 |
| Cladonia crispata (Ach.) Flot. | 0 | 0.5 ± 0.2 | <0.1 | 0.1 ± 0.1 | <0.1 | 0.3 ± 0.1 |
| Cladonia deformis (L.) Hoffm. | 0 | 3.2 ± 0.8 | 0 | <0.1 | 0 | 0.1 ± 0.1 |
| Cladonia gracilis (L.) Willd. | 0 | 0.3 ± 0.3 | <0.1 | <0.1 | 0.2 ± 0.1 | 1.0 ± 0.2 |
| Cladonia mitis Sandst. | 0 | 1.1 ± 0.4 | 0 | 0.5 ± 0.4 | 0.4 ± 0.1 | 1.4 ± 0.3 |
| Cladonia rangiferina (L.) Web. | 0 | <0.1 | 1.9 ± 0.8 | 1.2 ± 0.2 | 1.8 ± 0.4 | 4.6 ± 0.7 |
| Cladonia stellaris (Opiz) Pouzar&Vezda | 0 | 0 | 0.5 ± 0.4 | 0.5 ± 0.2 | 0.5 ± 0.2 | 8 ± 2 |
| Cladonia uncialis (L.)Weber ex FH Wigg. | 0 | 0 | 0 | 0.2 ± 0.05 | 0.1 ± 0.1 | 0.4 ± 0.2 |
| Nephroma arcticum (L.) Torss. | 0 | 0 | 0 | 2 ± 1 | 2 ± 1 | 0.5 ± 0.3 |
| Peltigera aphthosa (L.) Willd. | 0 | 0 | 0 | 0 | 0.2 ± 0.1 | 1.1 ± 0.05 |
| Trapeliopsis granulosa (Hoffm.) Lumbsch | 0.3 ± 0.05 | 1.1 ± 0.6 | 0 | 0 | 0 | 0.1 ± 0.1 |
| Primary thalli | 0.9 ± 0.05 | 37 ± 4 | <0.1 | 0 | 0 | <0.1 |
| Dicranum polysetum Sw. | 0.3 ± 0.05 | 4.0 ± 0.3 | 0.1 ± 0.05 | 0.3 ± 0.02 | 0.4 ± 0.1 | 0.3 ± 0.1 |
| Dicranum scoparium Hedw. | 0.8 ± 0.05 | 0.4 ± 0.05 | 1.1 ± 0.2 | 4 ± 1 | 6.1 ± 0.9 | 7 ± 1 |
| Hylocomium splendens (Hedw.) B. S. G. | 0 | 0 | 6 ± 1.5 | 8 ± 2 | 1.9 ± 0.7 | 0.9 ± 0.5 |
| Pleurozium schreberi (Willd. ex Brid.) Mitt. | 0 | 2.5 ± 0.7 | 41 ± 3 | 55 ± 4 | 52 ± 2 | 38 ± 3 |
| Pohlia nutans (Hedw.) Lindb. | 4 ± 1 | 12 ± 2 | 0 | 0 | 0.1 ± 0.05 | <0.1 |
| Polytrichum juniperinum Hedw. | 0.4 ± 0.05 | 0.4 ± 0.05 | 0 | 0 | 0 | 0.1 ± 0.1 |
| Polytrichum piliferum Hedw. | 0 | 0.3 ± 0.1 | 0 | 0.1 ± 0.1 | 0 | <0.1 |
| Ptilidium pulcherrimum (Weber) Hampe | 0.1 ± 0.05 | 0 | 0 | 0 | 0.3 ± 0.3 | 0.5 ± 0.1 |
| Hepaticae | 1.0 ± 0.1 | 0.7 ± 0.3 | 0.1 ± 0.05 | 1.1 ± 0.4 | 2.4 ± 0.3 | 3.2±0.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stavrova, N.I.; Gorshkov, V.V.; Katjutin, P.N.; Bakkal, I.J. The Structure of Northern Siberian Spruce–Scots Pine Forests at Different Stages of Post-Fire Succession. Forests 2020, 11, 558. https://doi.org/10.3390/f11050558
Stavrova NI, Gorshkov VV, Katjutin PN, Bakkal IJ. The Structure of Northern Siberian Spruce–Scots Pine Forests at Different Stages of Post-Fire Succession. Forests. 2020; 11(5):558. https://doi.org/10.3390/f11050558
Chicago/Turabian StyleStavrova, Natalia I., Vadim V. Gorshkov, Paul N. Katjutin, and Irina Ju. Bakkal. 2020. "The Structure of Northern Siberian Spruce–Scots Pine Forests at Different Stages of Post-Fire Succession" Forests 11, no. 5: 558. https://doi.org/10.3390/f11050558
APA StyleStavrova, N. I., Gorshkov, V. V., Katjutin, P. N., & Bakkal, I. J. (2020). The Structure of Northern Siberian Spruce–Scots Pine Forests at Different Stages of Post-Fire Succession. Forests, 11(5), 558. https://doi.org/10.3390/f11050558