Physiological and Biochemical Traits in Korean Pine Somatic Embryogenesis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Plant Material and Embryogenic Culture
2.2. Maturation of Somatic Embryos
2.3. Microscopic Observation
2.4. Sample Collection
2.5. FW, DW and Water Content of EMs
2.6. Physiological Determinations
2.6.1. Soluble Sugar Content
2.6.2. Starch Content
2.6.3. Soluble Protein Content
2.6.4. Polyphenol Content
2.6.5. Proline Content
2.6.6. Antioxidant Enzyme Activities
2.6.7. Statistical Analysis
3. Results
3.1. Characterization of Analyzed Tissues
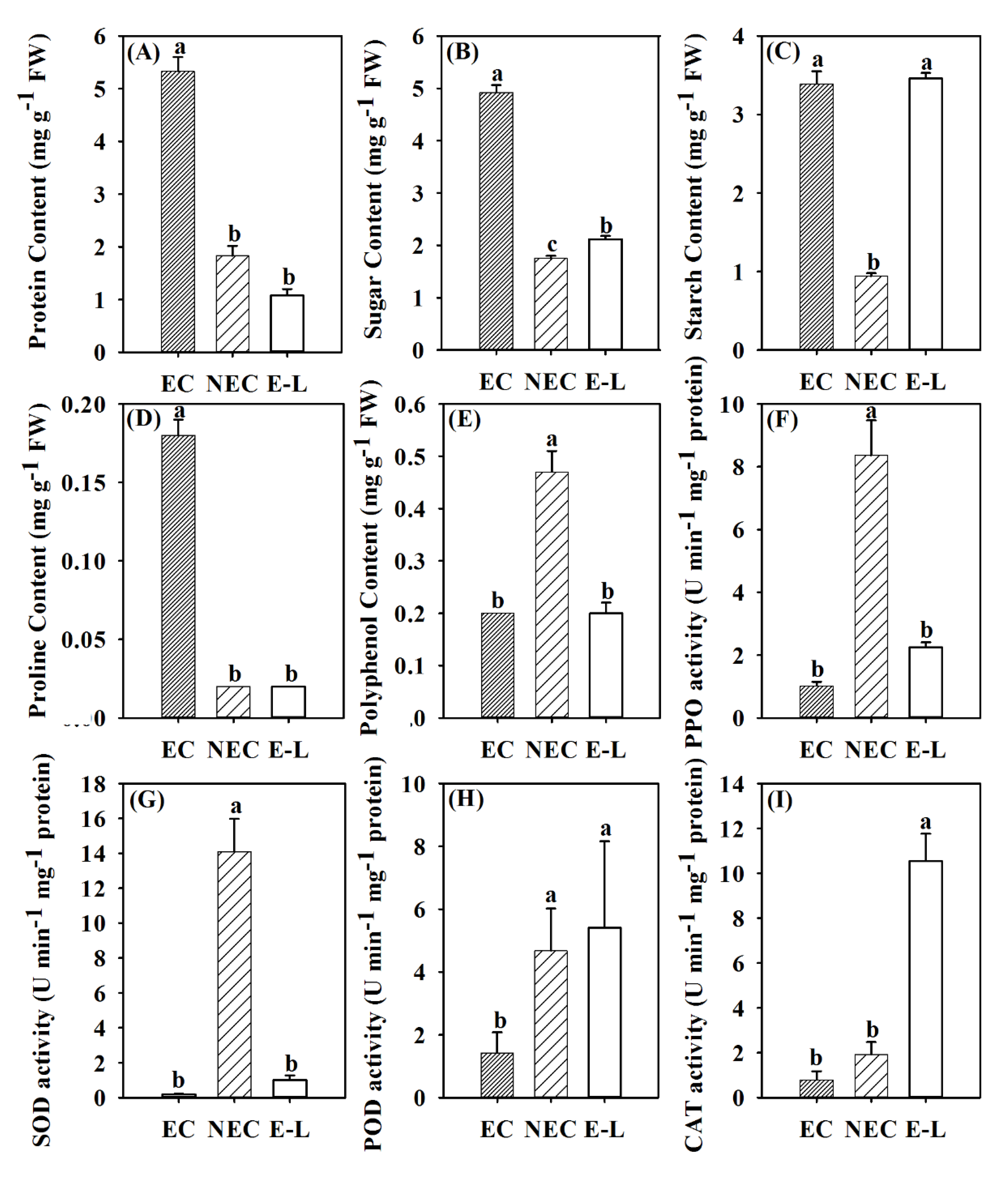
3.2. Physiological Differences in Different Types of Callus
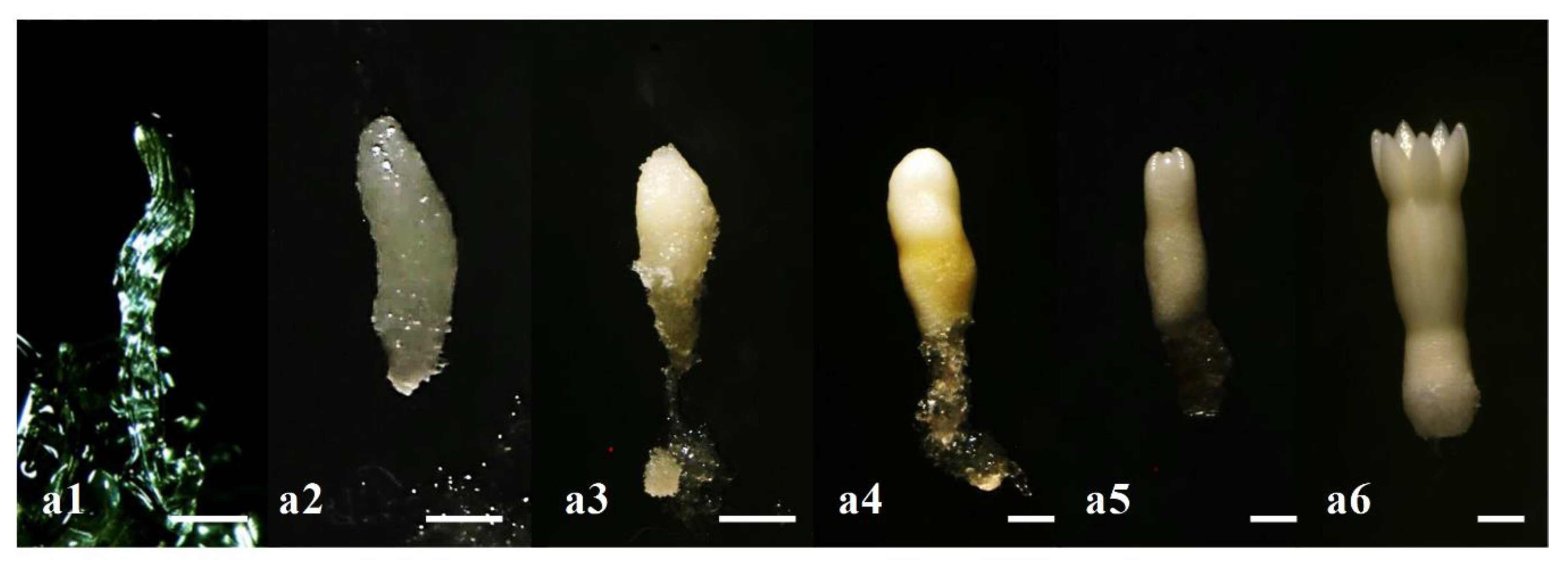
3.3. Morphological Changes during Somatic Embryogenesis in Korean Pine
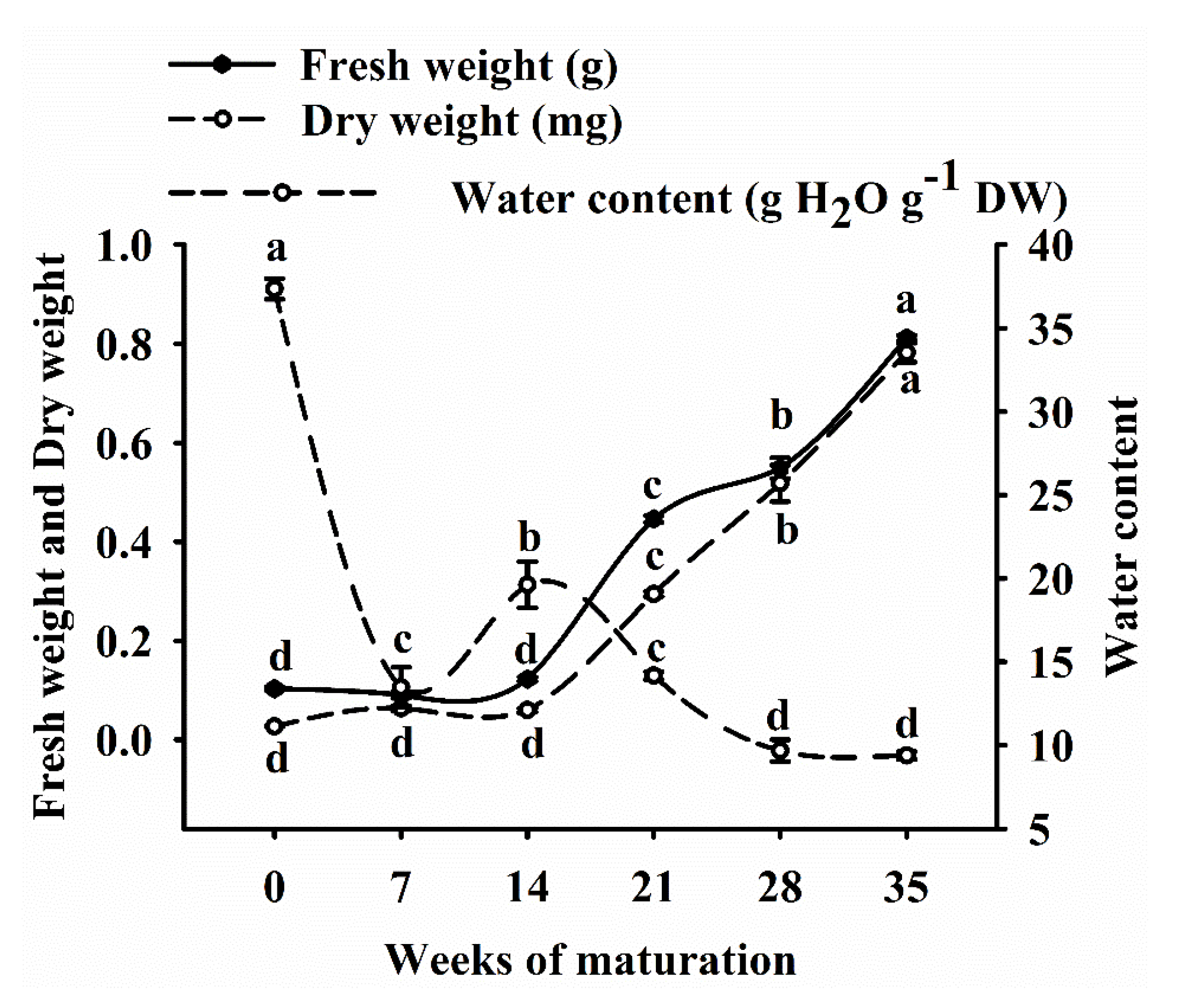
3.4. Fresh Weight, Dry Weight and Water Content of EC
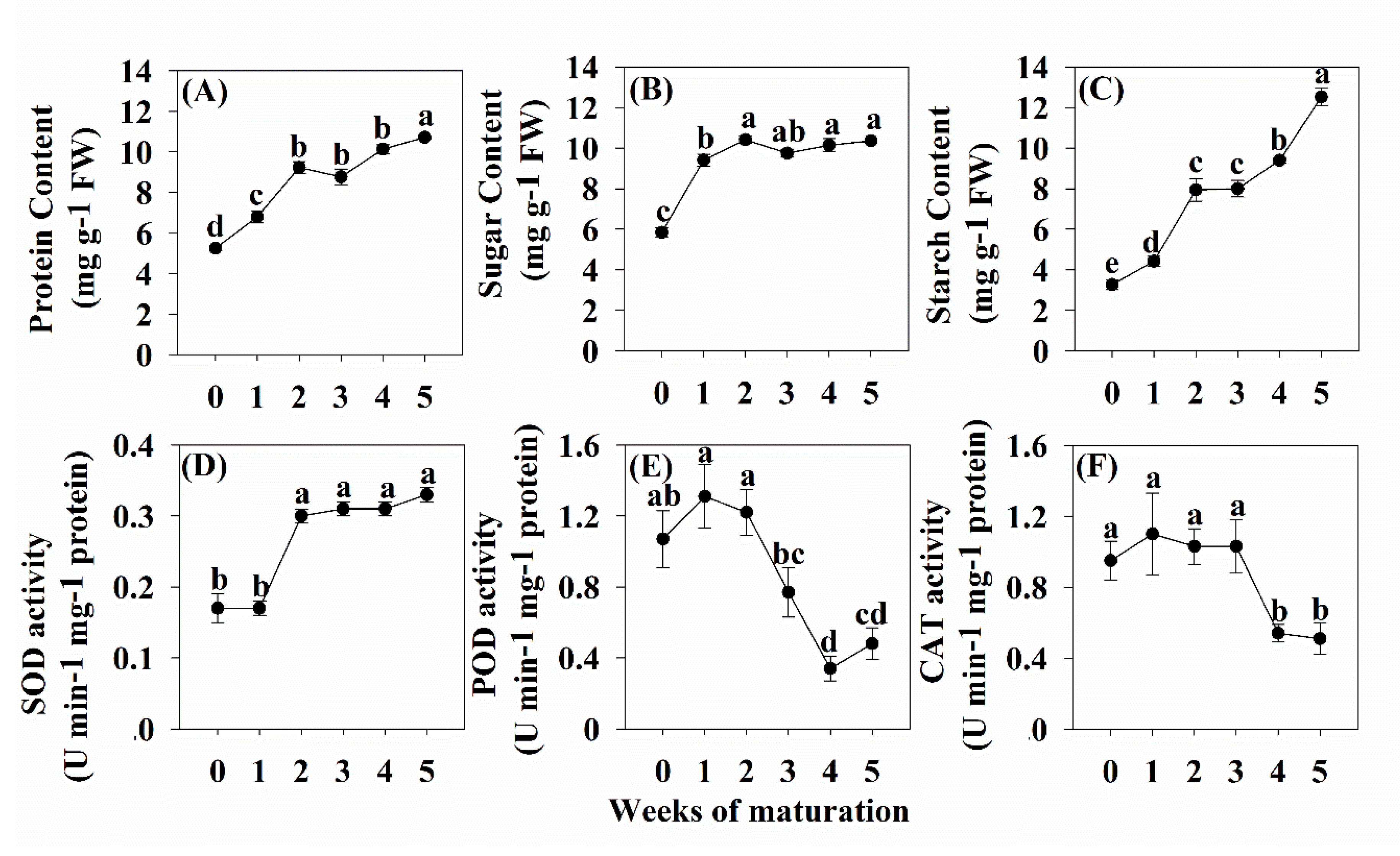
3.5. Levels of Soluble Sugar, Soluble Protein and Starch
3.6. Levels of SOD, POD and CAT Activity
4. Discussion
4.1. Physiological Differences between Different Types of Callus
4.2. Physiological Changes during Somatic Embryo Development of Korean Pine
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SE(s) | Somatic embryo(s) |
| EC | Embryogenic callus |
| 2,4-D | 2.4-dichlorophenoxyacetic acid |
| SOD | Superoxide dismutase |
| POD | Peroxidase |
| CAT | Catalase |
| NEC | Non-embryonic |
| E-L | The callus which lost somatic embryogenesis potential |
| PPO | Polyphenol oxidase |
| FW | Fresh weight |
| DW | Dry weight |
| NAA | 1-Naphthaleneacetic acid |
| 6-BA | 6-benzyl amino-purine acid |
| CH | Casein hydrolysate |
| ABA | Abscisic acid |
| NBT | Nitroblue tetrazolium |
References
- Llebres, M.T.; Pascual, M.B.; Debille, S.; Trontin, J.F.; Harvengt, L.; Avila, C.; Canovas, F.M. The role of arginine metabolic pathway during embryogenesis and germination in maritime pine (Pinus pinaster Ait.). Tree Physiol. 2018, 38, 471–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.; He, H.; Dai, L.; Yang, J. Effects of human disturbances on Korean pine coverage and age structure at a landscape scale in Northeast China. Ecol. Eng. 2014, 71, 375–379. [Google Scholar] [CrossRef]
- Egertsdotter, U. Automation and scale up of somatic embryogenesis for commercial plant production, with emphasis on Conifers. Front Plant Sci. 2019, 10, 109. [Google Scholar] [CrossRef] [Green Version]
- Lelu-Walter, M.A.; Thompson, D.; Harvengt, L.; Sanchez, L.; Toribio, M.; Pâques, L.E. Somatic embryogenesis in forestry with a focus on Europe: State-of-the-art, benefits, challenges and future direction. Tree Genet. Genomes 2013, 9, 883–899. [Google Scholar] [CrossRef]
- Guan, Y.; Li, S.G.; Fan, X.F.; Su, Z.H. Application of somatic embryogenesis in woody plants. Front Plant Sci. 2016, 7, 938. [Google Scholar] [CrossRef] [Green Version]
- Bozhkov, P.; Ahn, I.; Park, Y. Two alternative pathways of somatic embryo origin from polyembryonic mature stored seeds of Pinus koraiensis Sieb et Zucc. Can. J. Bot. 1997, 75, 509–512. [Google Scholar] [CrossRef]
- Gao, F.; Shen, H.L.; Liu, C.P.; Wang, Y.; Zhang, P.; Yang, L. Optimization of culture conditions and selection of suitable explants for callus induction from mature embryo of Pinus koraiensis. J. Nanjing For. Univ. 2016, 41, 43–50. [Google Scholar]
- Klubicová, K.; Uvácková, L.; Danchenko, M.; Nemecek, P.; Skultéty, L.; Salaj, J.; Salaj, T. Insights into the early stage of Pinus nigra Arn. somatic embryogenesis using discovery proteomics. J. Proteom. 2017, 169, 99–111. [Google Scholar] [CrossRef]
- Silveira, V.; De Vita, A.M.; Macedo, A.F.; Dias, M.F.R.; Floh, E.I.S.; Santa-Catarina, C. Morphological and polyamine content changes in embryogenic and non-embryogenic callus of sugarcane. Plant Cell Tissue Organ 2013, 114, 351–364. [Google Scholar] [CrossRef]
- Neves, N.; Segura-Nieto, M.; Blanco, M.A.; Sánchez, M.; González, A.; González, J.L.; Castillo, R. Biochemical characterization of embryogenic and non-embryogenic calluses of sugarcane. In Vitro Cell Dev. Plant 2003, 39, 343–345. [Google Scholar] [CrossRef]
- Gao, F.; Pen, C.X.; Wang, H.; Shen, H.L.; Yand, L. Selection of culture conditions for callus induction and proliferation by somatic embryogenesis of Pinus koraiensis. J. For. Res. 2020. [Google Scholar] [CrossRef]
- Mahmud, I.; Shrestha, B.; Boroujerdi, A.; Chowdhury, K. NMR-based metabolomics profile comparisons to distinguish between embryogenic and non-embryogenic callus tissue of sugarcane at the biochemical level. In Vitro Cell. Dev. Plant. 2015, 51, 340–349. [Google Scholar] [CrossRef]
- Maadon, S.N.; Rohani, E.R.; Ismail, I.; Baharum, S.N.; Normah, M.N. Somatic embryogenesis and metabolic differences between embryogenic and non-embryogenic structures in mangosteen. Plant Cell Tissue Organ 2016, 127, 443–459. [Google Scholar] [CrossRef]
- Dodeman, V.L.; Ducreux, G.; Kreis, M. Zygotic embryogenesis versus somatic embryogenesis. J. Exp. Bot. 1997, 48, 1493–1509. [Google Scholar]
- Jariteh, M.; Ebrahimzadeh, H.; Niknam, V.; Mirmasoumi, M.; Vahdati, K. Developmental changes of protein, proline and some antioxidant enzymes activities in somatic and zygotic embryos of Persian walnut (Juglans regia L.). Plant Cell Tissue Organ 2015, 122, 101–115. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Ali, H.; Yücesan, B.; Saeed, S.; Rehman, K.; Khan, M.A. Evaluation of biochemical markers during somatic embryogenesis in Silybum marianum L. 3 Biotech 2016, 6, 71. [Google Scholar] [CrossRef] [Green Version]
- Klimaszewska, K.; Noceda, C.; Pelletier, G.; Label, P.; Rodriguez, R.; Lelu-Walter, M.A. Biological characterization of young and aged embryogenic cultures of Pinus pinaster (Ait.). Vitr. Cell. Dev. Boil. Plant 2009, 45, 20. [Google Scholar] [CrossRef]
- Jo, L.; Dos Santos, A.L.; Bueno, C.A.; Barbosa, H.R.; Floh, E.I. Proteomic analysis and polyamines, ethylene and reactive oxygen species levels of Araucaria angustifolia (Brazilian pine) embryogenic cultures with different embryogenic potential. Tree Physiol. 2014, 34, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Filipović, B.K.; Simonović, A.D.; Trifunović, M.M.; Dmitrović, S.S.; Savić, J.M.; Jevremović, S.B.; Subotić, A.R. Plant regeneration in leaf culture of Centaurium erythraea Rafn. Part 1: The role of antioxidant enzymes. Plant Cell Tissue Organ 2015, 121, 703–719. [Google Scholar] [CrossRef]
- Fukumoto, T.; Hayashi, N.; Sasamoto, H. Atomic force microscopy and laser confocal scanning microscopy analysis of callose fibers developed from protoplasts of embryogenic cells of a conifer. Planta 2005, 223, 40–45. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.K.; Durzan, D.J. Somatic polyembryogenesis from callus of mature sugar pine embryos. Bio/technology 1986, 4, 643–645. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Park, Y.S.; Overton, C.; Maceacheron, I.; Bonga, J.M. Optimized somatic embryogenesis in Pinus strobus L. In Vitro Cell Dev. Boil. Plant 2001, 37, 392–399. [Google Scholar] [CrossRef]
- Von Arnold, S.; Sabala, I.; Bozhkov, P.; Dyachok, J.; Filonova, L. Developmental pathways of somatic embryogenesis. Plant Cell Tissue Organ 2002, 69, 233–249. [Google Scholar] [CrossRef]
- Morel, A.; Teyssier, C.; Trontin, J.F.; Eliášová, K.; Pešek, B.; Beaufour, M.; Morabito, D.; Boizot, N.; Le Metté, C.; Belal Bessai, L. Early molecular events involved in Pinus pinaster Ait. somatic embryo development under reduced water availability: Transcriptomic and proteomic analyses. Physiol. Plant. 2014, 152, 184–201. [Google Scholar] [CrossRef] [Green Version]
- Somani, B.L.; Khanade, J.; Sinha, R. A modified anthrone-sulfuric acid method for the determination of fructose in the presence of certain proteins. Anal. Biochem. 1987, 167, 327–330. [Google Scholar] [CrossRef]
- Vale, E.M.; Reis, R.S.; Passamani, L.Z.; Santa-Catarina, C.; Silveira, V. Morphological analyses and variation in carbohydrate content during the maturation of somatic embryos of Carica papaya. Physiol. Mol. Biol. Plant 2018, 24, 295–305. [Google Scholar] [CrossRef]
- Colvin, H.W.; Attebery, J.T.; Ivy, J.T. Comparison of the Anthrone Reagent and a Copper-Reduction Method for Determining Blood Sugar in Calves. J. Dairy Sci. 1961, 44, 2081–2088. [Google Scholar] [CrossRef]
- McCready, R.; Guggolz, J.; Silviera, V. Owens H: Determination of starch and amylose in vegetables. Anal. Chem. 1950, 22, 1156–1158. [Google Scholar] [CrossRef]
- Liu, M.X.; Xu, Z.G.; Yang, Y.; Feng, Y.J. Effects of different spectral lights on Oncidium PLBs induction, proliferation, and plant regeneration. Plant Cell Tissue Organ 2011, 106, 1–10. [Google Scholar]
- Kaewubon, P.; Hutadilok-Towatana, N.; Da Silva, J.A.T. Meesawat U: Ultrastructural and biochemical alterations during browning of pigeon orchid (Dendrobium crumenatum Swartz) callus. Plant Cell Tissue Organ 2015, 121, 53–69. [Google Scholar] [CrossRef]
- Helaly, M.N.; El-Hosieny, H.A.; El-Sarkassy, N.M.; Fuller, M.P. Growth, lipid peroxidation, organic solutes, and anti-oxidative enzyme content in drought-stressed date palm embryogenic callus suspension induced by polyethylene glycol. In Vitro Cell Dev. Boil. Plant 2017, 53, 133–141. [Google Scholar] [CrossRef]
- Rahnama, H.; Ebrahimzadeh, H. The effect of NaCl on antioxidant enzyme activities in potato seedlings. Biol. Plant. 2005, 49, 93–97. [Google Scholar] [CrossRef]
- Park, S.Y.; Klimaszewska, K.; Park, J.Y.; Mansfield, S.D. Lodgepole pine: The first evidence of seed-based somatic embryogenesis and the expression of embryogenesis marker genes in shoot bud cultures of adult trees. Tree Physiol. 2010, 30, 1469–1478. [Google Scholar] [CrossRef] [PubMed]
- Businge, E.; Bygdell, J.; Wingsle, G.; Moritz, T.; Egertsdotter, U. The effect of carbohydrates and osmoticum on storage reserve accumulation and germination of Norway spruce somatic embryos. Physiol. Plant. 2013, 149, 273–285. [Google Scholar] [CrossRef]
- Ree, J.F.; Polesi, L.G.; Back, F.; Bertolazi, A.A.; Silveira, V.; Guerra, M.P. Aging peach palm (Bactris gasipaes Kunth) cultures lose embryogenic potential and metabolic cellular function due to continuous culture in hypoxic environments. Plant Cell Tissue Organ 2020, 140, 49–67. [Google Scholar] [CrossRef]
- Passamani, L.Z.; Reis, R.S.; Vale, E.M.; Sousa, K.R.; Aragão, V.P.; Santa-Catarina, C.; Silveira, V. Long-term culture with 2, 4-dichlorophenoxyacetic acid affects embryogenic competence in sugarcane callus via changes in starch, polyamine and protein profiles. Plant Cell Tissue Organ 2020, 140, 415–429. [Google Scholar] [CrossRef]
- Wang, X.; Shi, L.; Lin, G.; Pan, X.; Chen, H.; Wu, X.; Takáč, T.; Šamaj, J.; Xu, C. A systematic comparison of embryogenic and non-embryogenic cells of banana (Musa spp. AAA): Ultrastructural, biochemical and cell wall component analyses. Sci. Hortic. 2013, 159, 178–185. [Google Scholar] [CrossRef]
- Fatima, S.; Mujib, A.; Samaj, J. Anti-oxidant enzyme responses during in vitro embryogenesis in Catharanthus roseus. J. Hortic. Sci. Biotechnol. 2011, 86, 569–574. [Google Scholar] [CrossRef]
- Pinto, D.L.P.; De Almeida, A.M.R.; Rêgo, M.M.; Da Silva, M.L.; De Oliveira, E.J.; Otoni, W.C. Somatic embryogenesis from mature zygotic embryos of commercial passionfruit (Passiflora edulis Sims) genotypes. Plant Cell Tissue Organ 2011, 107, 521–530. [Google Scholar] [CrossRef]
- Tang, W.; Newton, R.J. Increase of polyphenol oxidase and decrease of polyamines correlate with tissue browning in Virginia pine (Pinus virginiana Mill.). Plant Sci. 2004, 167, 621–628. [Google Scholar] [CrossRef]
- De Oliveira Prudente, D.; De Souza, L.B.; Paiva, R. Plant Somatic Embryogenesis: Modulatory Role of Oxidative Stress. Proc. Natl. Acad. Sci. India Sect. B: Biol. Sci. 2019, 89, 1–5. [Google Scholar]
- Orłowska, A.; Kępczyńska, E. Oxidative status in Medicago truncatula Gaertn. non-embryogenic and embryogenic tissues with particular reference to somatic embryogenesis. Plant Cell Tissue Organ 2020, 140, 35–48. [Google Scholar]
- Varghese, B.; Pammenter, N.; Berjak, P. Cryo-tolerance of zygotic embryos from recalcitrant seeds in relation to oxidative stress—a case study on two amaryllid species. J. Plant Physiol. 2012, 169, 999–1011. [Google Scholar]
- Medeiros De Araújo Silva, M.; Medeiros, L.E.; Jaislanny, M.; Cavalcante Granja Ma Nuela Willadino, L.; Camara, T. Antioxidant enzymes activity in embryogenic and non-embryogenic tissues in sugarcane. Acta Biológica Colomb. 2014, 19, 203–210. [Google Scholar] [CrossRef]
- Morel, A.; Trontin, J.F.; Corbineau, F.; Lomenech, A.M.; Beaufour, M.; Reymond, I.; Le Metté, C.; Ader, K.; Harvengt, L.; Cadene, M. Cotyledonary somatic embryos of Pinus pinaster Ait. most closely resemble fresh, maturing cotyledonary zygotic embryos: Biological, carbohydrate and proteomic analyses. Planta 2014, 240, 1075–1095. [Google Scholar] [CrossRef] [Green Version]
- Noah, A.M.; Niemenak, N.; Sunderhaus, S.; Haase, C.; Omokolo, D.N.; Winkelmann, T.; Braun, H.P. Comparative proteomic analysis of early somatic and zygotic embryogenesis in Theobroma cacao L. J. Proteom. 2013, 78, 123–133. [Google Scholar] [CrossRef]
- Dos Santos, A.L.W.; Elbl, P.; Navarro, B.V.; De Oliveira, L.F.; Salvato, F.; Balbuena, T.S.; Floh, E.I.S. Quantitative proteomic analysis of Araucaria angustifolia (Bertol.) Kuntze cell lines with contrasting embryogenic potential. J. Proteom. 2016, 130, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Li, H.; Fu, S.; Chen, B.; Sun, W.; Zhang, J.; Zhang, J. An iTRAQ-based proteomics approach to clarify the molecular physiology of somatic embryo development in prince Rupprecht’s larch (Larix principis-rupprechtii Mayr). PLoS ONE 2015, 10, e0119987. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.D.; Datta, S. Antioxidant enzyme activities during in vitro morphogenesis of gladiolus and the effect of application of antioxidants on plant regeneration. Biol. Plant. 2003, 47, 179–183. [Google Scholar] [CrossRef]
- Zhang, S.G.; Han, S.Y.; Yang, W.H.; Wei, H.L.; Zhang, M.; Qi, L.W. Changes in H2O2 content and antioxidant enzyme gene expression during the somatic embryogenesis of Larix leptolepis. Plant Cell Tissue Organ 2010, 100, 21. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, C.; Gao, F.; Wang, H.; Shen, H.; Yang, L. Physiological and Biochemical Traits in Korean Pine Somatic Embryogenesis. Forests 2020, 11, 577. https://doi.org/10.3390/f11050577
Peng C, Gao F, Wang H, Shen H, Yang L. Physiological and Biochemical Traits in Korean Pine Somatic Embryogenesis. Forests. 2020; 11(5):577. https://doi.org/10.3390/f11050577
Chicago/Turabian StylePeng, Chunxue, Fang Gao, Hao Wang, Hailong Shen, and Ling Yang. 2020. "Physiological and Biochemical Traits in Korean Pine Somatic Embryogenesis" Forests 11, no. 5: 577. https://doi.org/10.3390/f11050577
APA StylePeng, C., Gao, F., Wang, H., Shen, H., & Yang, L. (2020). Physiological and Biochemical Traits in Korean Pine Somatic Embryogenesis. Forests, 11(5), 577. https://doi.org/10.3390/f11050577