The Effect of Topography on Long-Term Spontaneous Development of Soil and Woody Cover on Graded and Untreated Overburden




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
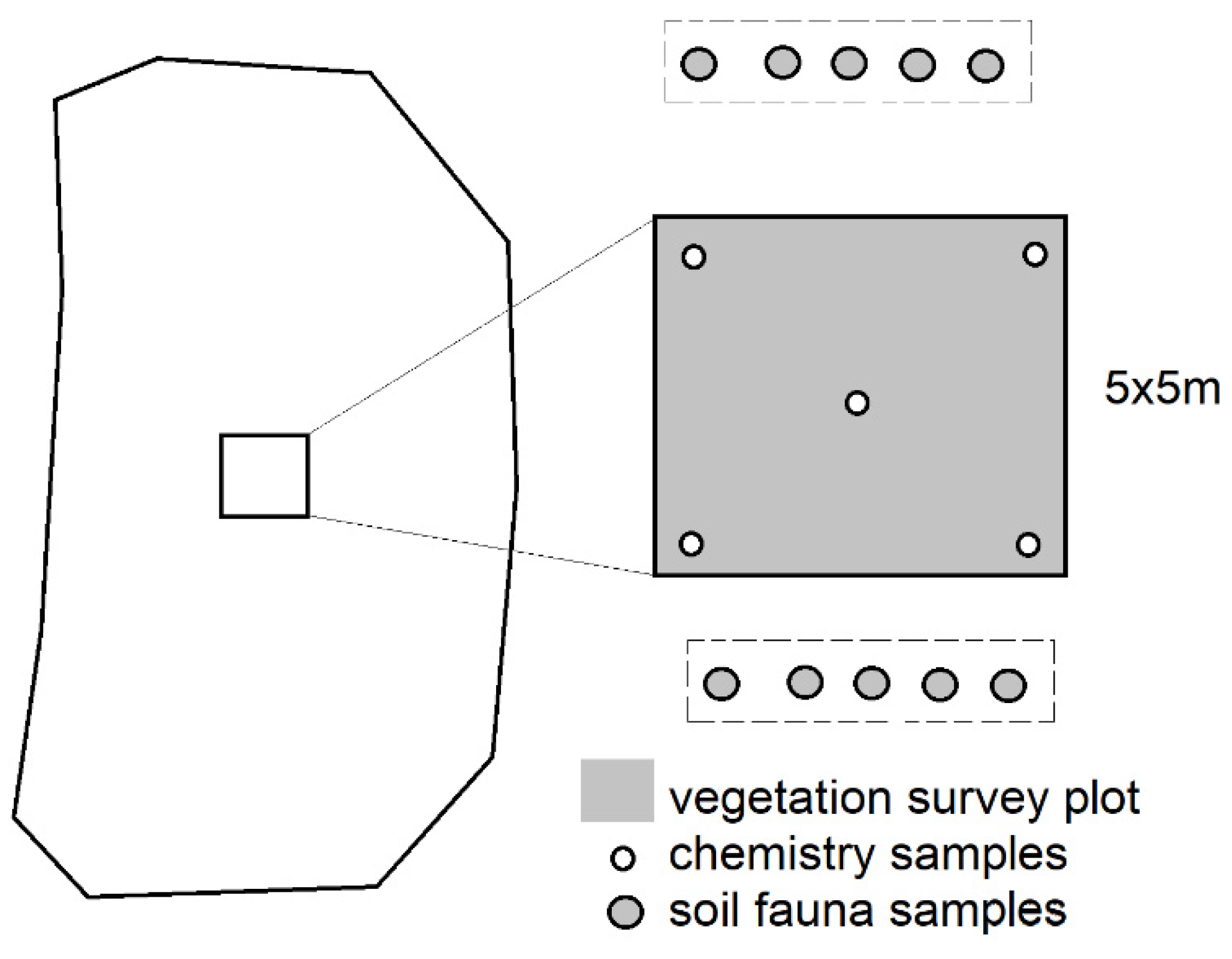
2.2. Sampling and Analyses
2.3. Statistical Evaluation
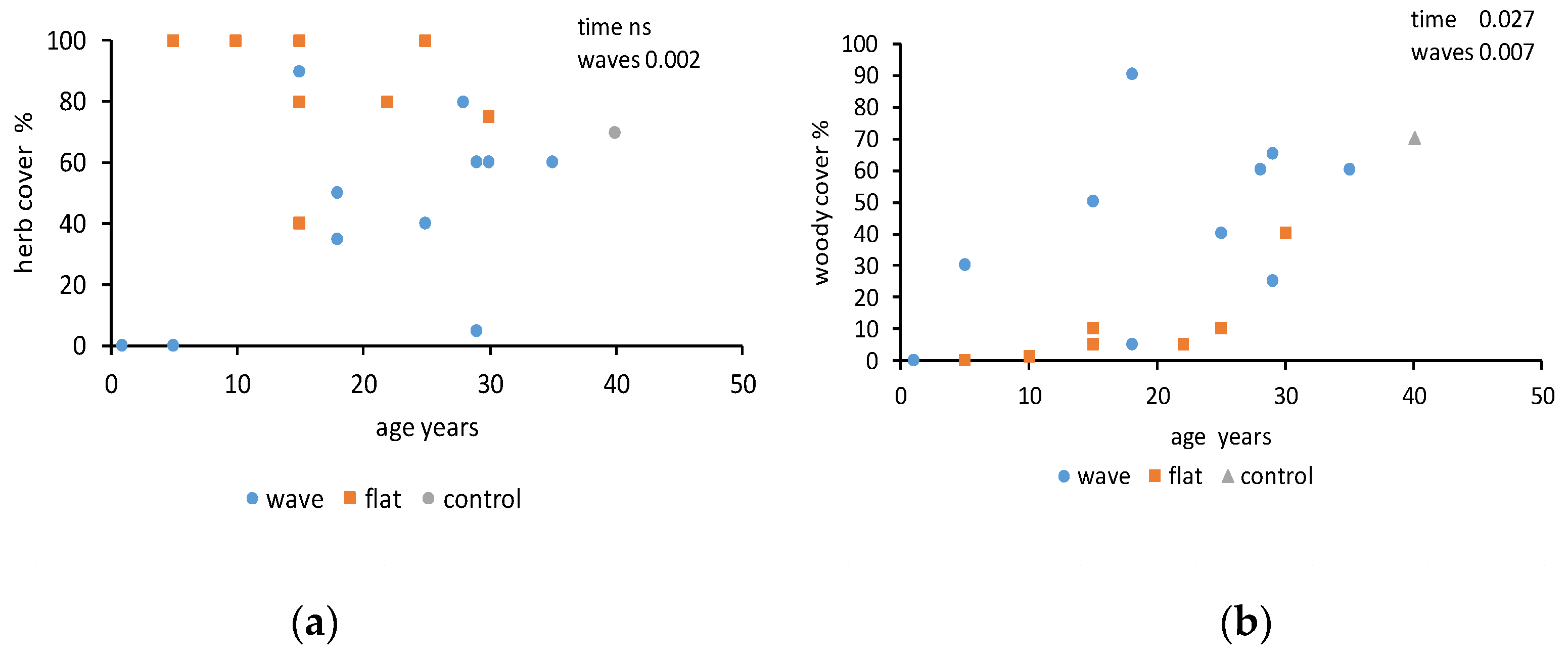
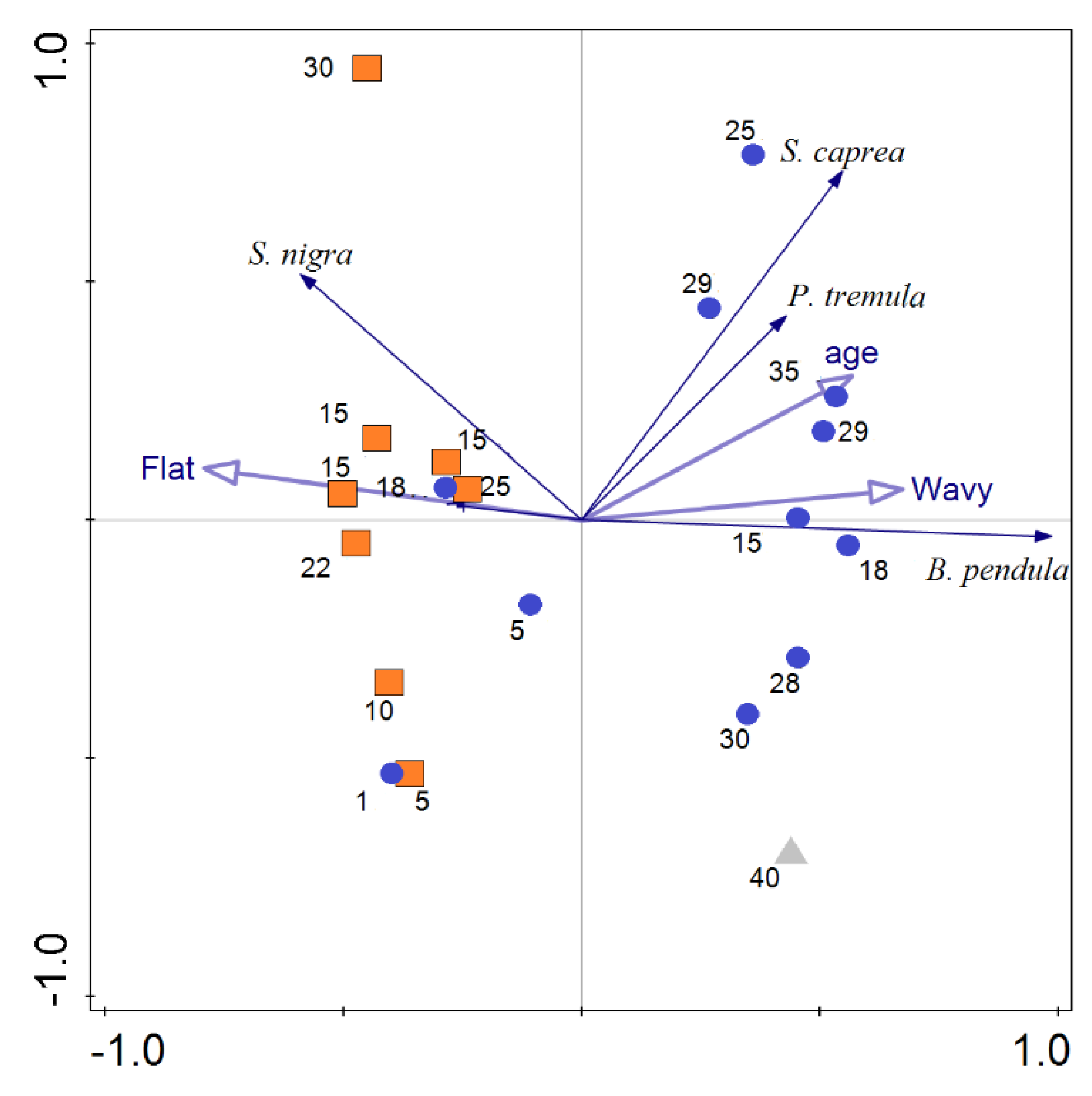
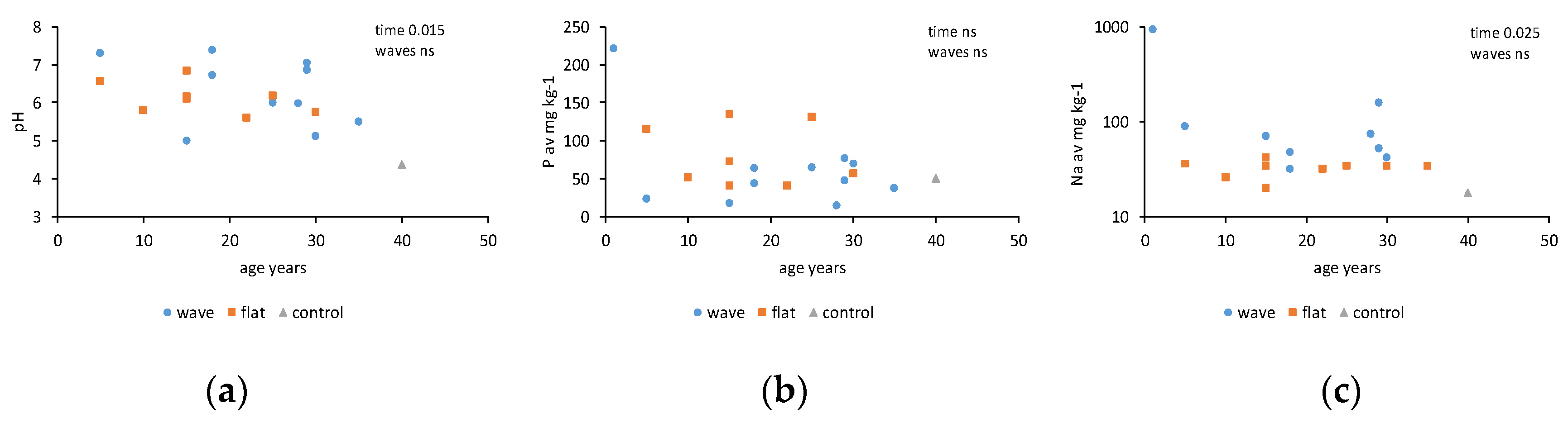
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mudrák, O.; Frouz, J.; Velichová, V. Understory vegetation in reclaimed and unreclaimed post-mining forest stands. Ecol. Eng. 2010, 36, 783–790. [Google Scholar] [CrossRef]
- Tropek, R.; Cerna, I.; Straka, J.; Čížek, O.; Konvička, M. Is coal combustion the last chance for vanishing insects of inland drift sand dunes in Europe? Biol. Conserv. 2013, 162, 60–64. [Google Scholar] [CrossRef]
- Tropek, R.; Hejda, M.; Kadlec, T.; Spitzer, L. Local and landscape factors affecting communities of plants and diurnal Lepidoptera in black coal spoil heaps: Implications for restoration management. Ecol. Eng. 2013, 57, 252–260. [Google Scholar] [CrossRef]
- Tropek, R.; Kadlec, T.; Hejda, M.; Kocarek, P.; Skuhrovec, J.; Malenovsky, I.; Vodka, S.; Spitzer, L.; Banar, P.; Konvička, M. Technical reclamations are wasting the conservation potential of post-mining sites. A case study of black coal spoil dumps. Ecol. Eng. 2012, 43, 13–18. [Google Scholar] [CrossRef]
- Frouz, J.; Mudrák, O.; Reitschmiedová, E.; Walmsley, A.; Vachová, P.; Šimáčková, H.; Albrechtová, J.; Moradi, J.; Kučera, J. Rough wave-like heaped overburden promotes establishment of woody vegetation while leveling promotes grasses during unassisted post mining site development. J. Environ. Manag. 2018, 205, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Prach, K.; Hobbs, R.J. Spontaneous succession versus technical reclamation in the restoration of disturbed sites. Restor. Ecol. 2008, 16, 363–366. [Google Scholar] [CrossRef]
- Tropek, R.; Kadlec, T.; Karesova, P.; Spitzer, L.; Kocarek, P.; Malenovsky, I.; Banar, P.; Tuf, I.H.; Hejda, M.; Konvička, M. Spontaneous succession in limestone quarries as an effective restoration tool for endangered arthropods and plants. J. Appl. Ecol. 2010, 47, 139–147. [Google Scholar] [CrossRef]
- Tropek, R.; Konvička, M. Should restoration damage rare biotopes? Biol. Conserv. 2011, 144, 1299. [Google Scholar] [CrossRef]
- Shu, W.S.; Ye, Z.H.; Zhang, Z.Q.; Lan, C.Y.; Wong, M.H. Natural Colonization of Plants on Five Lead/Zinc Mine Tailings in Southern China. Restor. Ecol. 2005, 13, 49–60. [Google Scholar] [CrossRef]
- Ashby, W.C. Reclamation with trees pre- and post-SMCRA in Southern Illinois. Int. J. Surf. Mining Reclam. Environ. 1998, 12, 117–121. [Google Scholar] [CrossRef]
- Zipper, C.E.; Burger, J.A.; Skousen, J.G.; Angel, P.N.; Barton, C.D.; Davis, V.; Franklin, J.A. Restoring forests and associated ecosystem services on Appalachian coal surface mines. Environ. Manag. 2011, 47, 751–765. [Google Scholar] [CrossRef] [PubMed]
- Cejpek, J.; Kuráž, V.; Frouz, J. Hydrological properties of soils in reclaimed and unreclaimed sites after brown-coal mining. Polish J. Environ. Stud. 2013, 22, 645–652. [Google Scholar]
- Kuráž, V.; Frouz, J.; Kuráž, M.; Mako, A.; Shustr, V.; Cejpek, J.; Romanov, O.V.; Abakumov, E.V. Changes in some physical properties of soils in the chronosequence of self-overgrown dumps of the Sokolov quarry-dump complex, Czechia. Eurasian Soil Sci. 2012, 45, 266–272. [Google Scholar] [CrossRef]
- Baasch, A.; Kirmer, A.; Tischew, S. Nine years of vegetation development in a postmining site: Effects of spontaneous and assisted site recovery. J. Appl. Ecol. 2012, 49, 251–260. [Google Scholar] [CrossRef]
- Boscutti, F.; Vianello, A.; Bozzato, F.; Casolo, V. Vegetation structure, species life span, and exotic status elucidate plant succession in a limestone quarry reclamation. Restor. Ecol. 2017, 25, 595–604. [Google Scholar] [CrossRef]
- Prach, K.; Řehounková, K.; Lencová, K.; Jírová, A.; Konvalinková, P.; Mudrák, O.; Študent, V.; Vaněček, Z.; Tichý, L.; Petřík, P.; et al. Vegetation succession in restoration of disturbed sites in Central Europe: The direction of succession and species richness across 19 seres. Appl. Veg. Sci. 2014, 17, 193–200. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Liu, X.; Zhou, M.; Ai, D.; Wang, G.; Wang, Y.; Chu, C.; Lundholm, J.T. The effect of environmental heterogeneity on species richness depends on community position along the environmental gradient. Sci. Rep. 2015, 5, 15723. [Google Scholar] [CrossRef]
- Frouz, J.; Pižl, V.; Tajovský, K.; Starý, J.; Holec, M.; Materna, J. Soil macro- and mesofauna succession in post-mining sites and other disturbed areas. In Soil Biota and Ecosystem Development in Post Mining Sites; Frouz, J., Ed.; CRC: Boca Rato, FL, USA, 2014. [Google Scholar]
- Williams, B.M.; Houseman, G.R. Experimental evidence that soil heterogeneity enhances plant diversity during community assembly. J. Plant Ecol. 2014, 7, 461–469. [Google Scholar] [CrossRef]
- Hendrychová, M.; Šálek, M.; Tajovský, K.; Řehoř, M. Soil properties and species richness of invertebrates on afforested sites after brown coal mining. Restor. Ecol. 2012, 20, 561–567. [Google Scholar] [CrossRef]
- Heneberg, P.; Bogusch, P.; Řehounek, J. Sandpits provide critical refuge for bees and wasps (Hymenoptera: Apocrita). J. Insect Conserv. 2013, 17, 473–490. [Google Scholar] [CrossRef]
- Doležalová, J.; Vojar, J.; Smolová, D.; Solský, M.; Kopecký, O. Technical reclamation and spontaneous succession produce different water habitats: A case study from Czech post-mining sites. Ecol. Eng. 2012, 43, 5–12. [Google Scholar] [CrossRef]
- Tichánek, F.; Tropek, R. Conservation value of post-mining headwaters: Drainage channels at a lignite spoil heap harbour threatened stream dragonflies. J. Insect Conserv. 2015, 19, 975–985. [Google Scholar] [CrossRef]
- Deák, B.; Valkó, O.; Török, P.; Kelemen, A.; Miglécz, T.; Szabó, S.; Szabó, G.; Tóthmérész, B. Micro-topographic heterogeneity increases plant diversity in old stages of restored grasslands. Basic Appl. Ecol. 2015, 16, 291–299. [Google Scholar] [CrossRef]
- Frouz, J.; Kalčík, J.; Velichová, V. Factors causing spatial heterogeneity in soil properties, plant cover, and soil fauna in a non-reclaimed post-mining site. Ecol. Eng. 2011, 37, 1910–1913. [Google Scholar] [CrossRef]
- Kappes, H.; Clausius, A.; Topp, W. Historical small-scale surface structures as a model for post-mining land reclamation. Restor. Ecol. 2012, 20, 322–330. [Google Scholar] [CrossRef]
- Moradi, J.; Potocký, P.; Kočárek, P.; Bartuška, M.; Tajovský, K.; Tichánek, F.; Frouz, J.; Tropek, R. Influence of surface flattening on biodiversity of terrestrial arthropods during early stages of brown coal spoil heap restoration. J. Environ. Manag. 2018, 220, 1–7. [Google Scholar] [CrossRef]
- Franklin, J.A.; Zipper, C.E.; Burger, J.A.; Skousen, J.G.; Jacobs, D.F. Influence of herbaceous ground cover on forest restoration of eastern US coal surface mines. New For. 2012, 43, 905–924. [Google Scholar] [CrossRef]
- Zeleznik, J.; Skousen, J. Survival of three tree species on old reclaimed surface mines in Ohio. J. Environ. Qual. 1996, 25, 1429–1435. [Google Scholar] [CrossRef] [Green Version]
- Frouz, J.; Nováková, A. Development of soil microbial properties in topsoil layer during spontaneous succession in heaps after brown coal mining in relation to humus microstructure development. Geoderma 2005, 129, 54–64. [Google Scholar] [CrossRef]
- Frouz, J.; Prach, K.; Pižl, V.; Háněl, L.; Starý, J.; Tajovský, K.; Materna, J.; Balík, V.; Kalčík, J.; Řehounková, K. Interactions between soil development, vegetation and soil fauna during spontaneous succession in post mining sites. Eur. J. Soil Biol. 2008, 44, 109–121. [Google Scholar] [CrossRef]
- Frouz, J.; Kalčík, J. Accumulation of soil organic carbon in relation to other soil characteristic during spontaneous succession in non reclaimed colliery spoil heaps after brown coal mining near Sokolov (the Czech Republic). Ekologia 2006, 25, 388–397. [Google Scholar]
- Benes, J.; Kepka, P.; Konvička, M. Limestone quarries as refuges for European xerophilous butterflies. Conserv. Biol. 2003, 17, 1058–1069. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vicentini, F.; Hendrychova, M.; Tajovský, K.; Pižl, V.; Frouz, J. The Effect of Topography on Long-Term Spontaneous Development of Soil and Woody Cover on Graded and Untreated Overburden. Forests 2020, 11, 602. https://doi.org/10.3390/f11050602
Vicentini F, Hendrychova M, Tajovský K, Pižl V, Frouz J. The Effect of Topography on Long-Term Spontaneous Development of Soil and Woody Cover on Graded and Untreated Overburden. Forests. 2020; 11(5):602. https://doi.org/10.3390/f11050602
Chicago/Turabian StyleVicentini, Fabio, Markéta Hendrychova, Karel Tajovský, Václav Pižl, and Jan Frouz. 2020. "The Effect of Topography on Long-Term Spontaneous Development of Soil and Woody Cover on Graded and Untreated Overburden" Forests 11, no. 5: 602. https://doi.org/10.3390/f11050602
APA StyleVicentini, F., Hendrychova, M., Tajovský, K., Pižl, V., & Frouz, J. (2020). The Effect of Topography on Long-Term Spontaneous Development of Soil and Woody Cover on Graded and Untreated Overburden. Forests, 11(5), 602. https://doi.org/10.3390/f11050602