Global Geographic Distribution and Host Range of Fusarium circinatum, the Causal Agent of Pine Pitch Canker

 , ,
, ,  , ,
, ,  , ,
, ,  , ,
, ,  ,
,  , ,
, ,  , , ,
, , ,  , , , ,
, , , ,  , , , , ,
, , , , ,  , ,
, ,  add
Show full author list
add
Show full author list
Abstract
:1. Introduction
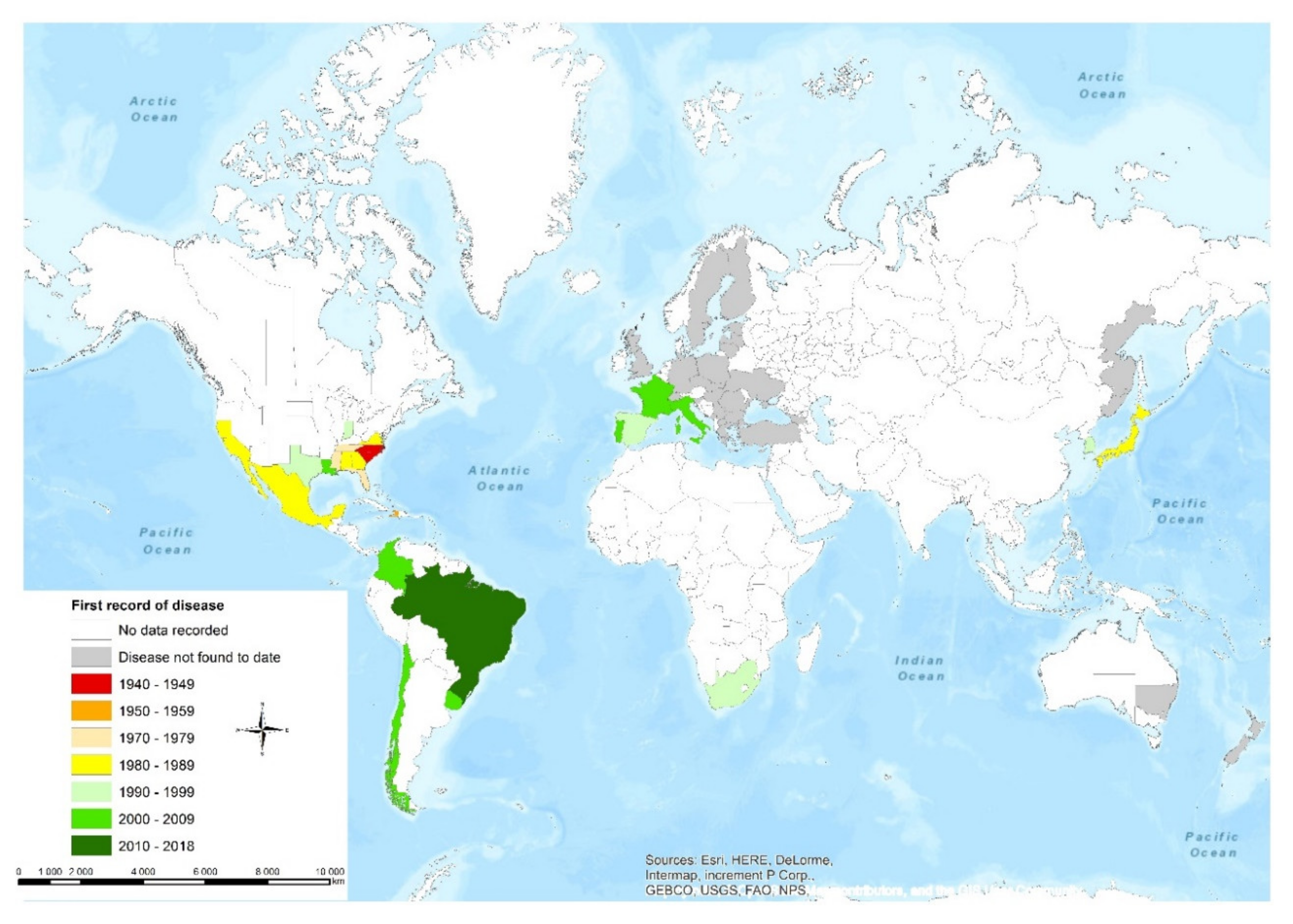
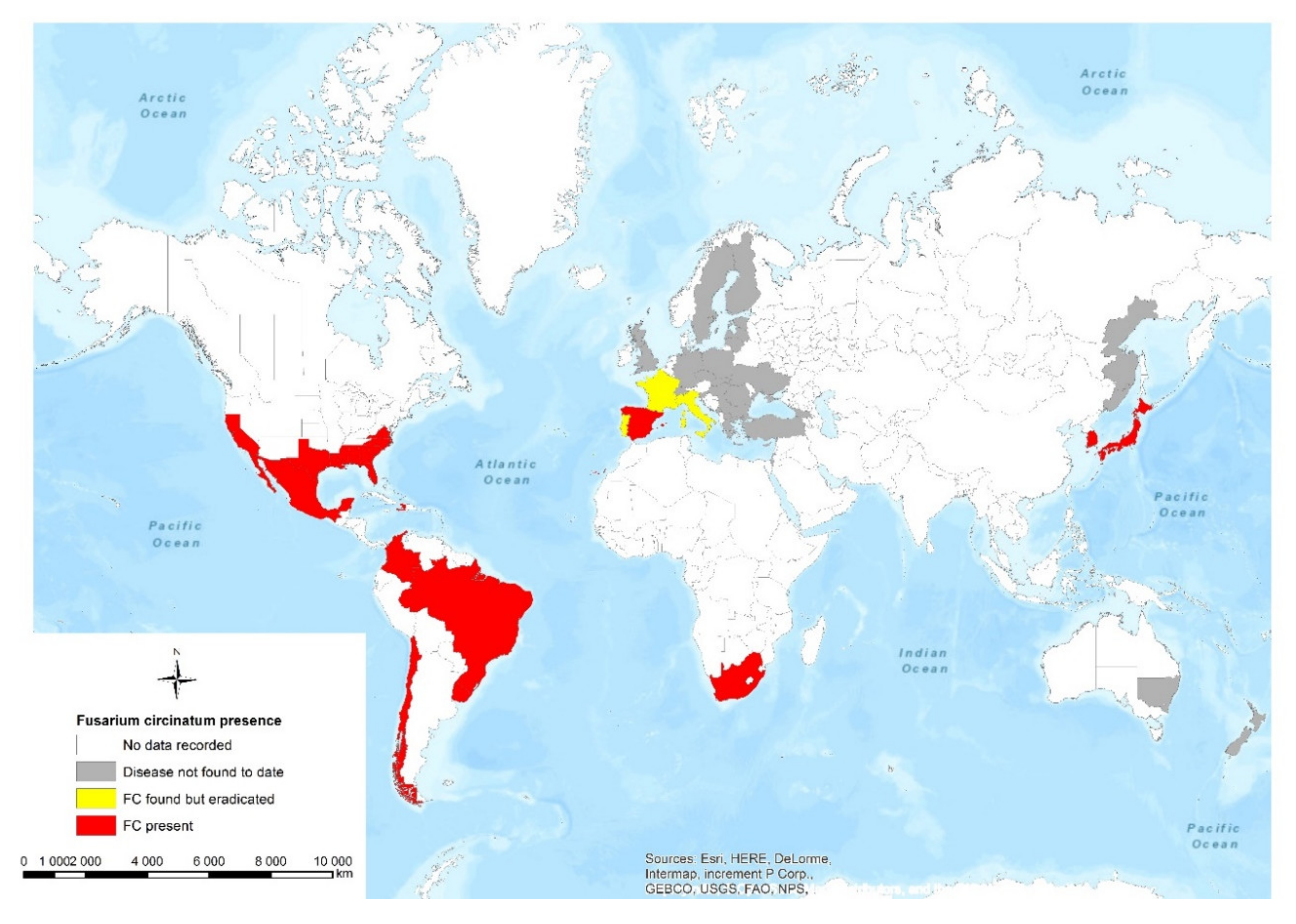
2. The Geographic Distribution of F. circinatum
2.1. Europe
2.2. North America and South America
2.3. Asia
2.4. Africa
2.5. Oceania
3. Taxonomy and Evolution of Fusarium circinatum
4. Population Dynamics of Fusarium circinatum
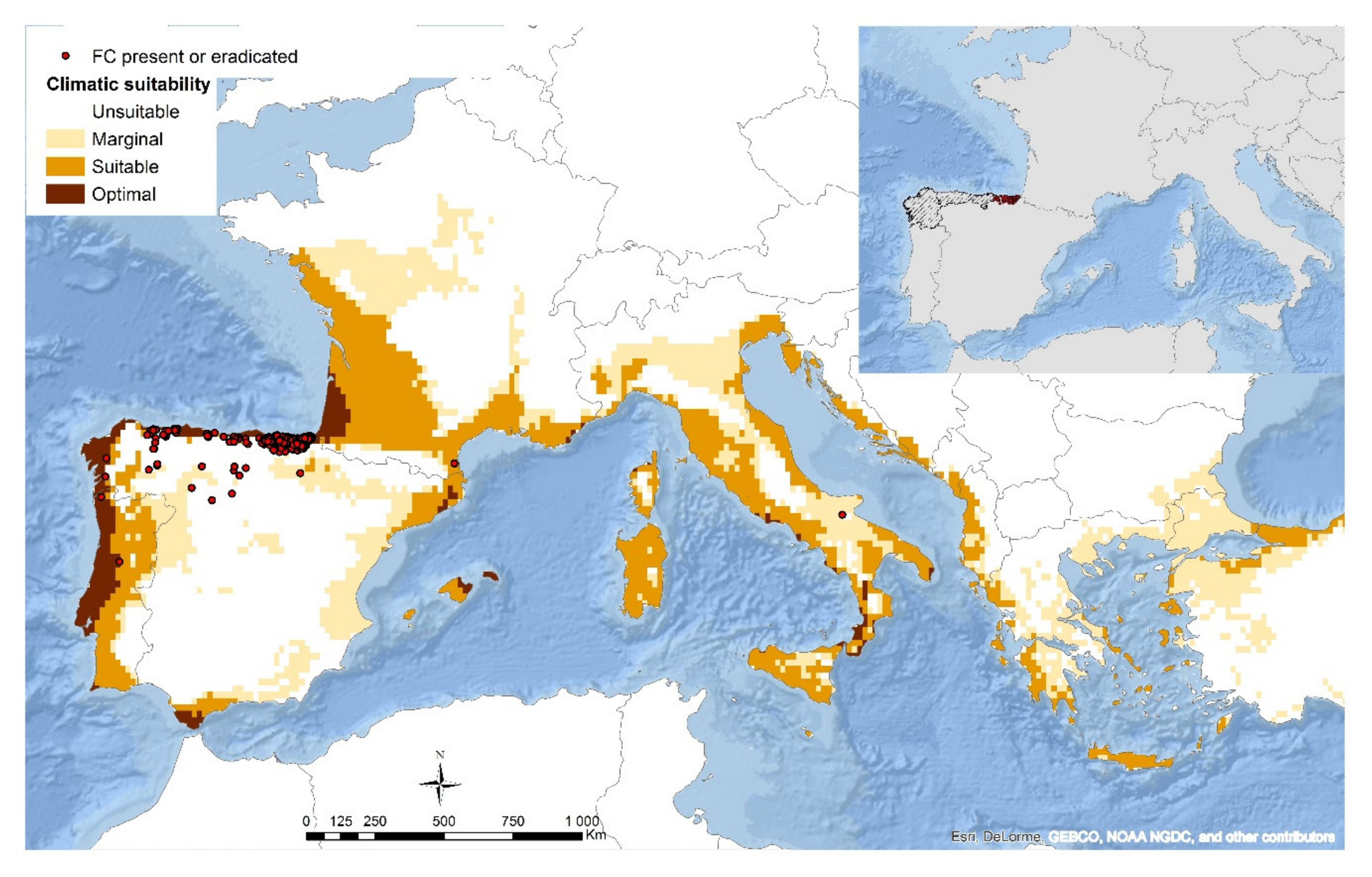
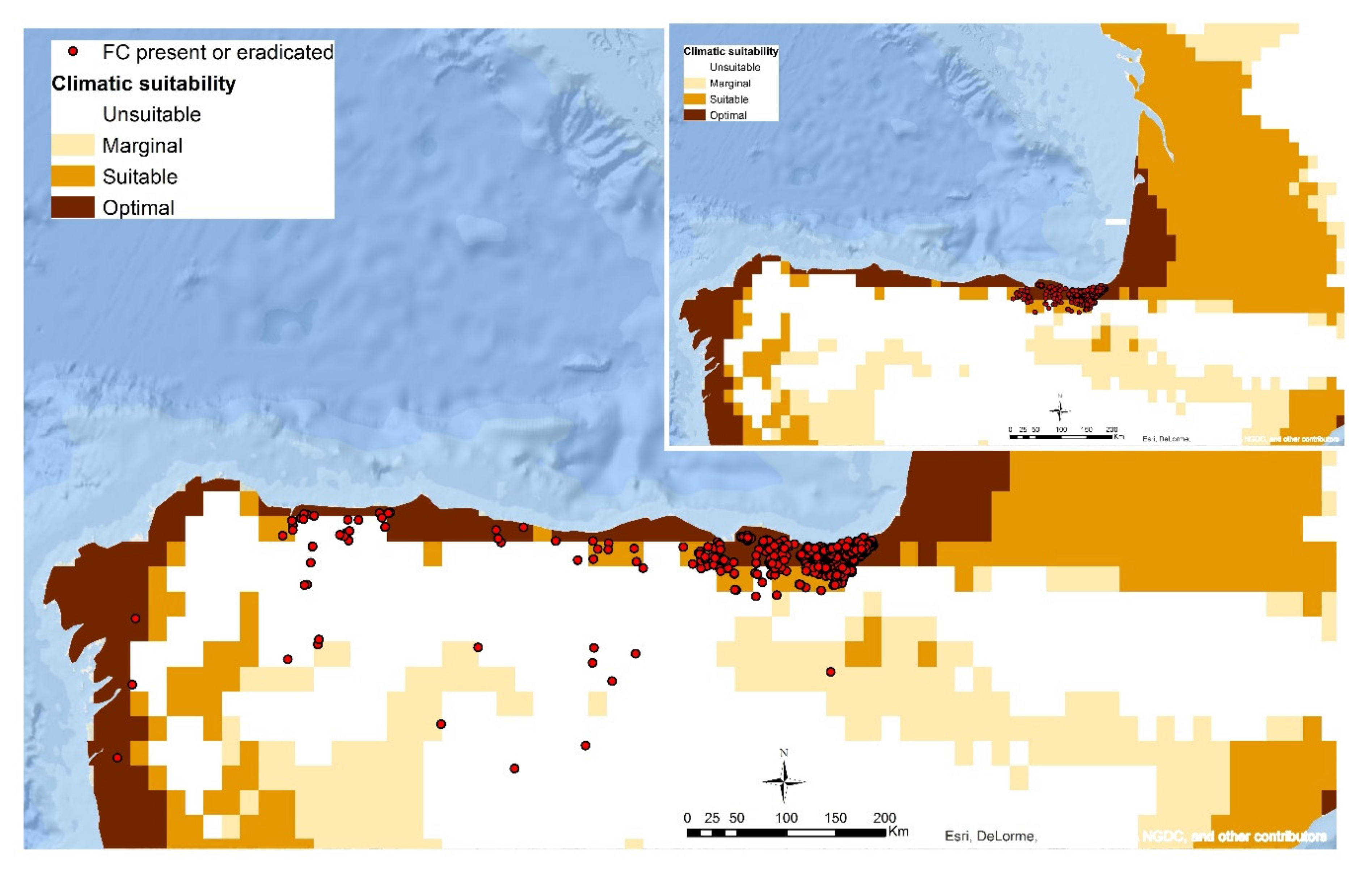
5. Climatic Influence on Fusarium circinatum Distribution and Modelling Potential Pathways of Introduction
6. Host Range
6.1. Host Susceptibility Ratings
6.2. Species with Variable Susceptibility Ratings
6.3. Analyses of Geo-Database Data and Host Range
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Geiser, D.M.; Aoki, T.; Bacon, C.W.; Baker, S.E.; Bhattacharyya, M.K.; Brandt, M.E.; Brown, D.W.; Burgess, L.W.; Chulze, S.; Coleman, J.J.; et al. One Fungus, One Name: Defining the Genus Fusarium in a Scientifically Robust Way That Preserves Longstanding Use. Phytopathology 2013, 103, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Vettraino, A.M.; Potting, R.; Raposo, R. EU Legislation on Forest Plant Health: An Overview with a Focus on Fusarium circinatum. Forests 2018, 9, 568. [Google Scholar] [CrossRef] [Green Version]
- García-Serna, I. Diplodia pinea (Desm.) Kickx y Fusarium circinatum Niremberg & O’Donell, Principales Hongos de Chancro de las Masas Forestales de Pinus radiata D. Don del País Vasco. Ph.D. Thesis, University of the Basque Country, Bilbao, Spain, 2011. [Google Scholar]
- Wingfield, M.J.; Hammerbacher, A.; Ganley, R.J.; Steenkamp, E.T.; Gordon, T.R.; Wingfield, B.D.; Coutinho, T. Pitch canker caused by Fusarium circinatum—A growing threat to pine plantations and forests worldwide. Australas. Plant Pathol. 2008, 37, 319–334. [Google Scholar] [CrossRef]
- Swett, C.L.; Gordon, T. First Report of Grass Species (Poaceae) as Naturally Occurring Hosts of the Pine Pathogen Gibberella circinata. Plant Dis. 2012, 96, 908. [Google Scholar] [CrossRef]
- Swett, C.L.; Porter, B.; Fourie, G.; Steenkamp, E.T.; Gordon, T.R.; Wingfield, M.J. Association of the pitch canker pathogen Fusarium circinatum with grass hosts in commercial pine production areas of South Africa. South. For. J. For. Sci. 2014, 76, 161–166. [Google Scholar] [CrossRef]
- Hernandez-Escribano, L.; Iturritxa, E.; Elvira-Recuenco, M.; Berbegal, M.; Campos, J.A.; Renobales, G.; Garcia, I.; Raposo, R. Herbaceous plants in the understory of a pitch canker-affected Pinus radiata plantation are endophytically infected with Fusarium circinatum. Fungal Ecol. 2018, 32, 65–71. [Google Scholar] [CrossRef]
- Martínez-Álvarez, P.; Pando, V.; Diez, J.J. Alternative species to replace Monterey pine plantations affected by pitch canker caused by Fusarium circinatum in northern Spain. Plant Pathol. 2014, 63, 1086–1094. [Google Scholar] [CrossRef]
- Martín-García, J.; Paraschiv, M.; Flores-Pacheco, J.A.; Chira, D.; Díez, J.J.; Fernández, M. Susceptibility of Several Northeastern Conifers to Fusarium circinatum and Strategies for Biocontrol. Forests 2017, 8, 318. [Google Scholar] [CrossRef] [Green Version]
- Martín-García, J.; Lukačevičová, A.; Flores-Pacheco, J.A.; Díez, J.J.; Dvořák, M. Evaluation of the Susceptibility of Several Czech Conifer Provenances to Fusarium circinatum. Forests 2018, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Storer, A.J.; Gordon, T.R.; Clark, S.L. Clark Association of the pitch canker fungus, Fusarium subglutinans f. sp. pini, with Monterey pine seeds and seedlings in California. Plant Pathol. 1998, 47, 649–656. [Google Scholar] [CrossRef]
- Viljoen, A. First Report of Fusarium subglutinans f. sp. pini on Pine Seedlings in South Africa. Plant Dis. 1994, 78, 309. [Google Scholar] [CrossRef]
- Wikler, K.; Storer, A.; Newman, W.; Gordon, T.; Wood, D. The dynamics of an introduced pathogen in a native Monterey pine (Pinus radiata) forest. For. Ecol. Manag. 2003, 179, 209–221. [Google Scholar] [CrossRef]
- Gordon, T.R. Biology and Management of Gibberella circinata, the cause of pitch canker in pines. In Control of Fusarium Diseases; Alves-Santos, F.M., Diez, J.J., Eds.; Research Signpost: Kerala, India, 2011; pp. 195–207. [Google Scholar]
- Bezos, D.; Martínez-Alvarez, P.; Fernández, M.; Diez, J.J. Epidemiology and management of pine pitch canker disease in Europe—A review. Balt. For 2017, 23, 279–293. [Google Scholar]
- Kelley, W.D. Incidence of Pitch Canker Among Clones of Loblolly Pine in Seed Orchards. Plant Dis. 1982, 66, 1171. [Google Scholar] [CrossRef]
- Dwinell, L.D. Pitch Canker: A Disease Complex of Southern Pines. Plant Dis. 1985, 69, 270. [Google Scholar] [CrossRef]
- Swett, C.L.; Reynolds, G.J.; Gordon, T.R. Infection without wounding and symptomless shoot colonization of Pinus radiata by Fusarium circinatum, the cause of pitch canker. For. Pathol. 2018, 48, 1–7. [Google Scholar] [CrossRef]
- Kuhlman, E.G.; Dianis, S.D.; Smith, T.K. Epidemiology of pitch canker disease in a loblolly pine seed orchard in North Carolina. Phytopathology 1982, 72, 1212–1216. [Google Scholar] [CrossRef]
- Runion, G.B.; Bruck, R.I. Associations of the pine tip moth with pitch canker of loblolly pine. Phytopathology 1985, 75, 1339. [Google Scholar]
- Aguayo, J.; Fourrier-Jeandel, C.; Capdevielle, X.; Vétillard, F.; Piou, D.; Iturritxa, E.; Robin, C. Assessment of molecular detection of Fusarium circinatum in insects and passive spore traps in Pinus radiata plantations. For. Pathol. 2020, 50, e12574. [Google Scholar] [CrossRef]
- Martín-Rodrigues, N.; Sanchez-Zabala, J.; Salcedo, I.; Majada, J.; González-Murua, C.; Duñabeitia, M.K. New insights into radiata pine seedling root infection byFusarium circinatum. Plant Pathol. 2015, 64, 1336–1348. [Google Scholar] [CrossRef]
- Zamora-Ballesteros, C.; Díez, J.J.; Martín-García, J.; Witzell, J.; Solla, A.; Ahumada, R.; Capretti, P.; Cleary, M.; Drenkhan, R.; Dvořák, M.; et al. Pine Pitch Canker (PPC): Pathways of Pathogen Spread and Preventive Measures. Forests 2019, 10, 1158. [Google Scholar] [CrossRef] [Green Version]
- Hepting, G.H.; Roth, E.R. Pitch canker, a new disease of southern pines. J. For. 1946, 44, 742–744. [Google Scholar]
- Anonymous. DP 22: Fusarium circinatum. ISPM 27 Diagnostic Protocols for Regulated Pests; Iternational Plant Protection Convention (IPPC): Rome, Italy, 2017; p. 22. [Google Scholar]
- CABI. Gibberella circinata (Pitch Canker). Available online: https://www.cabi.org/isc/datasheet/25153 (accessed on 27 May 2020).
- EPPO. Fusarium circinatum. Available online: https://gd.eppo.int/taxon/GIBBCI/distribution (accessed on 27 May 2020).
- Ganley, R.J.; Watt, M.; Manning, L.; Iturritxa, E. A global climatic risk assessment of pitch canker disease. Can. J. For. Res. 2009, 39, 2246–2256. [Google Scholar] [CrossRef]
- Baker, R.; Candresse, T.S.; Erzsébet Dormannsné, E.; Gilioli, G.; Grégoire, J.; Jeger, M.J. Risk assessment of Gibberella circinata for the EU territory and identification and evaluation of risk management options. EFSA 2010, 8, 1620. [Google Scholar]
- Möykkynen, T.; Capretti, P.; Pukkala, T. Modelling the potential spread of Fusarium circinatum, the causal agent of pitch canker in Europe. Ann. For. Sci. 2014, 72, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Santini, A.; Ghelardini, L.; de Pace, C.; Desprez-Loustau, M.L.; Capretti, P.; Chandelier, A.; Cech, T.; Chira, D.; Diamandis, S.; Gaitniekis, T.; et al. Biogeographical patterns and determinants of invasion by forest pathogens in Europe. New Phytol. 2012, 197, 238–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wingfield, M.; Brockerhoff, E.G.; Wingfield, B.D.; Slippers, B. Planted forest health: The need for a global strategy. Science 2015, 349, 832–836. [Google Scholar] [CrossRef]
- Collar Urquijo, J. Informe de la reunión del grupo de trabajo de laboratorios de diagnóstico y prospecciones fitosanitarias. Almería 1995, 7. [Google Scholar]
- Landeras, E.; Garcia, P.; Fernández, Y.; Braña, M.; Fernández-Alonso, O.; Méndez-Lodos, S.; Pérez-Sierra, A.; Leon, M.; Abad-Campos, P.; Berbegal, M.; et al. Outbreak of Pitch Canker Caused by Fusarium circinatum on Pinus spp. in Northern Spain. Plant Dis. 2005, 89, 1015. [Google Scholar] [CrossRef]
- Carlucci, A.; Colatruglio, L.; Frisullo, S. First Report of Pitch Canker Caused by Fusarium circinatum on Pinus halepensis and P. pinea in Apulia (Southern Italy). Plant Dis. 2007, 91, 1683. [Google Scholar] [CrossRef]
- EPPO. First Report of Gibberella Circinata in France; EPPO Reporting Service; EPPO Global Database: Paris, France, 2006; p. 104. [Google Scholar]
- Bragança, H.; Diogo, E.; Moniz, F.; Amaro, P. First Report of Pitch Canker on Pines Caused by Fusarium circinatum in Portugal. Plant Dis. 2009, 93, 1079. [Google Scholar] [CrossRef] [PubMed]
- Lauricica, J.M.; Muguruza, J.R. Presencia de Fusarium Subglutinan ssp. Pinien Viveros de Pino Radiata en Bizkaia. In Proceedings of the Actas XIV Reunión Anual del Grupo de Trabajo Fitosanitario de Forestales Parques YJardines Zaragoza, Zaragoza, Spain, 18–20 November 1997; pp. 301–303. [Google Scholar]
- Dwinell, L.D.; Adams, D.; Guerra-Santos, J.J.; Aguirre, J.R.M. Pitch Canker Disease of Pinus radiata. In Proceedings of the ICPP98—7th International Congress of Plant Pathology, London, UK, 9–16 August 1998; Available online: http://www.bspp.org.uk/icpp98/3.7/30.html (accessed on 27 April 2020).
- Dwinell, L.D. Global distribution of the pitch canker fungus. In Current and Potential Impacts of Pitch Canker in Radiata pine, IMPACT Monterey Workshop, Monterey, CA, USA, 30 November–3 December 1998; Devey, M.E., Matheson, A.C., Gordon, T.R., Eds.; CSIRO Forestry and Forest Products: Canberra, Australia, 1999; pp. 54–57. [Google Scholar]
- EPPO. Update of the Situation of Fusarium circinatum in Spain; EPPO Reporting Service; EPPO Global Database: Paris, France, 2019; p. 196. [Google Scholar]
- EPPO. Update of the Situation of Fusarium circinatum in Portugal; EPPO Reporting Service; EPPO Global Database: Paris, France, 2019; p. 170. [Google Scholar]
- EPPO. Gibberella Circinata Eradicated in France; EPPO Reporting Service; EPPO Global Database: Paris, France, 2008; p. 103. [Google Scholar]
- EPPO. Situation of Gibberella circinata in France; EPPO Reporting Service; EPPO Global Database: Paris, France, 2009; p. 093. [Google Scholar]
- EPPO. Gibberella circinata Detected Again in France; EPPO Reporting Service; EPPO Global Database: Paris, France, 2010; p. 034. [Google Scholar]
- Ioos, R.; Aloi, F.; Piškur, B.; Guinet, C.; Mullett, M.; Berbegal, M.; Bragança, H.; Cacciola, S.O.; Oskay, F.; Cornejo, C.; et al. Transferability of PCR-based diagnostic protocols: An international collaborative case study assessing protocols targeting the quarantine pine pathogen Fusarium circinatum. Sci. Rep. 2019, 9, 8195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elvira-Recuenco, M.; Cacciola, S.; Sanz-Ros, A.V.; Garbelotto, M.; Aguayo, J.; Solla, A.; Mullett, M.; Drenkhan, T.; Oskay, F.; Kaya, A.G.A.; et al. Potential Interactions between Invasive Fusarium circinatum and Other Pine Pathogens in Europe. Forests 2019, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Field Guide. Pine Pitch Canker. Available online: https://drive.google.com/file/d/1rA7EPEUzaBXikevC_skV9cpuu0VuCAYZ/view (accessed on 27 April 2020).
- Loit, K.; Adamson, K.; Bahram, M.; Puusepp, R.; Anslan, S.; Kiiker, R.; Drenkhan, R.; Tedersoo, L. Relative Performance of MinION (Oxford Nanopore Technologies) versus Sequel (Pacific Biosciences) Third-Generation Sequencing Instruments in Identification of Agricultural and Forest Fungal Pathogens. Appl. Environ. Microbiol. 2019, 85, 01368-19. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Drenkhan, R.; Anslan, S.; Morales-Rodriguez, C.; Cleary, M. High-throughput identification and diagnostics of pathogens and pests: Overview and practical recommendations. Mol. Ecol. Resour. 2018, 19, 47–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra-Santos, J.J. El Cancro Resinoso Causado por Subglurinans (Wollenw y Reink) Nelson, Toussoun y Marasas, Una Nueva Enfermedad de Pinus spp., en Mexico. Master’s Thesis, Universidad Autonoma Chapingo, Chapingo, Mexico, 1995. [Google Scholar]
- Wikler, K.; Gordon, T.R. An initial assessment of genetic relationships among populations of Fusarium circinatum in different parts of the world. Can. J. Bot. 2000, 78, 709–717. [Google Scholar]
- Hepting, G.H.; Roth, E.R. Host relations and spread of the pine pitch canker disease. Phytopathology 1953, 43, 475. [Google Scholar]
- Wingfield, M.; Jacobs, A.; Coutinho, T.; Ahumada, R.; Wingfield, B.D. First report of the pitch canker fungus, Fusarium circinatum, on pines in Chile. Plant Pathol. 2002, 51, 397. [Google Scholar] [CrossRef] [Green Version]
- Alonso, R.; Bettucci, L. First report of the pitch canker fungus Fusarium circinatum affecting Pinus taeda seedlings in Uruguay. Australas. Plant Dis. Notes 2009, 4, 91–92. [Google Scholar]
- Steenkamp, E.T.; Rodas, C.A.; Kvas, M.; Wingfield, M. Fusarium circinatum and pitch canker of Pinus in Colombia. Australas. Plant Pathol. 2012, 41, 483–491. [Google Scholar] [CrossRef] [Green Version]
- Rodas, C.A. Important Pest and Diseases of Plantation Grown Pinus and Eucalyptus in Colombia and Their Control. PhD. Thesis, University of Pretoria, Pretoria, South Africa, 2013. [Google Scholar]
- Pfenning, L.; Costa, S.D.S.; de Melo, M.P.; Costa, H.; Ventura, J.A.; Auer, C.G.; dos Santos, Á.F. First report and characterization of Fusarium circinatum, the causal agent of pitch canker in Brazil. Trop. Plant Pathol. 2014, 39, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.K.; Lee, S.H.; Sung, I.Y.; Lee, Y.W. First report of pitch canker disease on Pinus rigida in Korea. Plant Pathol. J. 2000, 16, 52–54. [Google Scholar]
- Kobayashi, T.; Muramoto, M. Pitch canker of Pinus luchuensis, a new disease of Japanese forests. For. Pests 1989, 40, 169–173. [Google Scholar]
- Morris, A.; Fourie, G.; Greyling, I.; Steenkamp, E.T.; Jones, N.B. Re-use of seedling containers and Fusarium circinatum association with asymptomatic Pinus patula planting stock. South. For. J. For. Sci. 2014, 76, 177–187. [Google Scholar] [CrossRef]
- Fourie, G.; Wingfield, M.J.; Wingfield, B.D.; Jones, N.B.; Morris, A.; Steenkamp, E.T. Culture-independent detection, and quantification of Fusarium circinatum in a pine-producing seedling nursery. South. For. J. For. Sci. 2014, 76, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Jones, N.B.; Ford, C.; Light, M.E.; Nadel, R.; Greyling, I.; Fourie, G.; Wingfield, M.J.; Morris, A. Effect on nursery and field performance of Pinus patula seedlings after inoculation with Fusarium circinatum. South. For. J. For. Sci. 2014, 76, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Fitza, K.; Payn, K.; Steenkamp, E.T.; Myburg, A.; Naidoo, S. Chitosan application improves resistance to Fusarium circinatum in Pinus patula. S. Afr. J. Bot. 2013, 85, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, R.; Steenkamp, E.T.; Coutinho, T.; Wingfield, M. The pitch canker fungus, Fusarium circinatum: Implications for South African forestry. South. For. J. For. Sci. 2011, 73, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Fru, F.F.; Steenkamp, E.T.; Wingfield, M.J.; Santana, Q.C.; Roux, J. Unique clones of the pitch canker fungus, Fusarium circinatum, associated with a disease outbreak in a new region of South Africa. Eur. J. Plant Pathol. 2017, 148, 97–107. [Google Scholar] [CrossRef]
- Coutinho, T.; Steenkamp, E.T.; Mongwaketsi, K.; Wilmot, M.; Wingfield, M.J. First outbreak of pitch canker in a South African pine plantation. Australas. Plant Pathol. 2007, 36, 256. [Google Scholar] [CrossRef]
- Steenkamp, E.T.; Makhari, O.M.; Coutinho, T.; Wingfield, B.D.; Wingfield, M. Evidence for a new introduction of the pitch canker fungus Fusarium circinatum in South Africa. Plant Pathol. 2013, 63, 530–538. [Google Scholar] [CrossRef]
- Santana, Q.C.; Coetzee, M.P.A.; Wingfield, B.D.; Wingfield, M.; Steenkamp, E.T. Nursery-linked plantation outbreaks and evidence for multiple introductions of the pitch canker pathogen Fusarium circinatum into South Africa. Plant Pathol. 2015, 65, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Fru, F.F.; Steenkamp, E.T.; Wingfield, M.; Roux, J. High genetic diversity of Fusarium circinatum associated with the first outbreak of pitch canker on Pinus patula in South Africa. South. For. J. For. Sci. 2018, 81, 69–78. [Google Scholar] [CrossRef]
- Mitchell, R.; Wingfield, M.; Steenkamp, E.T.; Coutinho, T. Tolerance of Pinus patula full-sib families to Fusarium circinatum in a greenhouse study. South. For. J. For. Sci. 2012, 74, 247–252. [Google Scholar] [CrossRef]
- Nel, A.; Hodge, G.R.; Mongwaketsi, K.E.; Kanzler, A. Genetic parameters for Fusarium circinatum tolerance within open pollinated families of Pinus patula tested at screening facilities in South Africa and the USA. South. For. 2014, 76, 145–150. [Google Scholar] [CrossRef]
- Mitchell, R.G.; Wingfield, M.; Steenkamp, E.T.; Roux, J.; Verryn, S.; Coutinho, T. Comparison of the tolerance of Pinus patula seedlings and established trees to infection by Fusarium circinatum. South. For. J. For. Sci. 2014, 76, 151–159. [Google Scholar] [CrossRef]
- Mitchell, R.G.; Wingfield, M.; Hodge, G.R.; Steenkamp, E.T.; Coutinho, T. Selection of Pinus spp. in South Africa for tolerance to infection by the pitch canker fungus. New For. 2011, 43, 473–489. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, R.G.; Wingfield, M.J.; Hodge, G.R.; Steenkamp, E.T.; Coutinho, T.A. The tolerance of Pinus patula × Pinus tecunumanii, and other pine hybrids, to Fusarium circinatum in greenhouse trials. New For. 2013, 44, 443–456. [Google Scholar] [CrossRef]
- Roux, J.; Eisenberg, B.; Kanzler, A.; Nel, A.; Coetzee, V.; Kietzka, E.; Wingfield, M.J. Testing of selected South African Pinus hybrids and families for tolerance to the pitch canker pathogen, Fusarium circinatum. New For. 2006, 33, 109–123. [Google Scholar] [CrossRef]
- Ormsby, M. Pitch canker in quarantine—A biosecurity success story. Biosecurity 2004, 51, 10. [Google Scholar]
- Smith, S. Plant Health Clinic News. 2013. Available online: https://www.uaex.edu/yard-garden/plant-health-clinic/docs/2013_Plant_Health_Clinic_Newsletters/plant%20health%20clinic%20newsletter%2013.pdf (accessed on 27 May 2020).
- McCain, A.H.; Koehler, C.S.; Tjosvold, S.A. Pitch canker threatens California pines. Calif. Agric. 1987, 41, 22–23. [Google Scholar]
- Correll, J.C. Pitch Canker Disease in California: Pathogenicity, Distribution, and Canker Development on Monterey Pine (Pinus radiata). Plant Dis. 1991, 75, 676. [Google Scholar] [CrossRef]
- Huang, J.W. Fungi Associated with Damping-off of Slash Pine Seedlings in Georgia. Plant Dis. 1990, 74, 27. [Google Scholar] [CrossRef]
- Blakeslee, G.; Jokela, E.; Hollis, C.; Wilson, D.; Lante, W.; Allen, J. Pitch Canker in Young Loblolly Pines: Influence of Precommercial Thinning and Fertilization on Disease Incidence and Severity. South. J. Appl. For. 1999, 23, 139–143. [Google Scholar] [CrossRef] [Green Version]
- McCay-Buis, T.S.; Abney, T.S.; Cummings, R.B.; Huber, D.M. Pitch canker disease of white pine seedlings in Indiana. Phytopathology 1994, 84, 1122. [Google Scholar]
- Starkey, D.; Meeker, J.; Mangini, A. Pitch Canker of Southern Pines and Recent Cases in Florida, Louisiana, Mississippi, and Texas. In USDA Forest Service Proceedings RMRS-P; USDA Forest Service: Fort Collins, CO, USA, 2007; Volume 50, pp. 97–103. [Google Scholar]
- Dwinell, L.D. Susceptibility of southern pines to infection by Fusarium moniliforme var. subglutinans. Plant Dis. Repor. 1978, 62, 108–111. [Google Scholar]
- Kuhlman, E.G.; Dwinell, L.D.; Nelson, P.E.; Booth, C. Characterization of the Fusarium causing pitch canker of southern pines. Mycologia 1978, 70, 1131–1143. [Google Scholar] [CrossRef]
- Correll, J.C.; Gordon, T.R.; McCain, A.H. Genetic diversity in California and Florida populations of the pitch pine canker fungus Fusarium subglutinans var. pini. Phytopathology 1992, 82, 415–420. [Google Scholar] [CrossRef]
- Viljoen, A. Characterization of Fusarium Isolates from Gladiolus Corms Pathogenic to Pines. Plant Dis. 1995, 79, 1240. [Google Scholar] [CrossRef]
- Britz, H.; Coutinho, T.; Wingfield, M.J.; Marasas, W.F.O.; Gordon, T.R.; Leslie, J.F. Fusarium subglutinans f. sp. pini Represents a Distinct Mating Population in theGibberella fujikuroi Species Complex. Appl. Environ. Microbiol. 1999, 65, 1198–1201. [Google Scholar] [CrossRef] [Green Version]
- Nirenberg, H.I.; O’Donnell, K. New Fusarium species and combinations within the Gibberella fujikuroi species complex. Mycologia 1998, 90, 434–458. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E.; Nirenberg, H.I. Molecular systematic and phylogeography of the Gibberella fujikuroi species complex. Mycologia 1998, 90, 465–493. [Google Scholar] [CrossRef]
- Britz, H.; Coutinho, T.A.; Wingfield, M.J.; Marasas, W.F.O. Validation of the description of Giberella circinata and morphological differentiation of the anamorph Fusarium circinatum. Sydowia 2002, 54, 9–22. [Google Scholar]
- Nelson, P.E.; Toussoun, T.A.; Marasas, W.F.O. Fusarium species: An Illustrated Manual of Identification; Penn State University Press: University Park, PA, USA, 1983. [Google Scholar]
- Hawksworth, D.L. A new dawn for the naming of fungi: Impacts of decisions made in Melbourne in July 2011 on the future publication and regulation of fungal names. IMA Fungus 2011, 2, 155–162. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Rooney, A.; Proctor, R.H.; Brown, D.W.; McCormick, S.P.; Ward, T.J.; Frandsen, R.J.N.; Lysøe, E.; Rehner, S.A.; Aoki, T.; et al. Phylogenetic analyses of RPB1 and RPB2 support a middle Cretaceous origin for a clade comprising all agriculturally and medically important fusaria. Fungal Genet. Boil. 2013, 52, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Kvas, M.; Marasas, W.F.O.; Wingfield, B.D.; Wingfield, M.J.; Steenkamp, E.T. Diversity, and evolution of Fusarium species in the Gibberella fujikuroi complex. Fungal Divers. 2009, 34, 1–21. [Google Scholar]
- Guerra-Santos, J.J. Pitch canker on Monterey pine in Mexico. In Current and Potential Impacts of Pitch Canker in Radiata pine, Proceedings of the IMPACT Monterey Workshop, Monterey, CA, USA, 30 November–3 December 1998; Devey, M.E., Matheson, A.C., Gordon, T.R., Eds.; CSIRO Forestry and Forest Products: Canberra, Australia, 1999; pp. 58–61. [Google Scholar]
- Iltis, H.H. Homeotic Sexual Translocations and the Origin of Maize (Zea Mays, Poaceae): A New look at an old problem. Econ. Bot. 2000, 54, 7–42. [Google Scholar] [CrossRef]
- Swett, C.L.; Gordon, T.R. Endophytic association of the pine pathogen Fusarium circinatum with corn (Zea mays). Fungal Ecol. 2015, 13, 120–129. [Google Scholar] [CrossRef]
- Herron, D.; Wingfield, M.; Wingfield, B.D.; Rodas, C.; Marincowitz, S.; Steenkamp, E.T. Novel taxa in the Fusarium fujikuroi species complex from Pinus spp. Stud. Mycol. 2015, 80, 131–150. [Google Scholar] [CrossRef] [Green Version]
- Perry, J.P.; Graham, A.; Richardson, D. The history of pines in Mexico and Central America. In Ecology and Biogeography of Pinus; Richardson, D.M., Ed.; Cambridge University Press: Cambridge, UK, 2000; pp. 137–149. [Google Scholar]
- Millar, C.I. Impact of the Eocene on the Evolution of Pinus L. Ann. Mo. Bot. Gard. 1993, 80, 471. [Google Scholar] [CrossRef]
- Leslie, J.F. Gibberella fujikuroi: Available populations and variable traits. Can. J. Bot. 1995, 73, 282–291. [Google Scholar] [CrossRef]
- Martin, S.H.; Wingfield, B.D.; Wingfield, M.J.; Steenkamp, E.T. Structure, and evolution of the Fusarium mating type locus: New insights from the Gibberella fujikuroi complex. Fungal Genet. Boil. 2011, 48, 731–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covert, S.F.; Briley, A.; Wallace, M.; McKinney, V.T. Partial MAT-2 Gene Structure, and the Influence of Temperature on Mating Success in Gibberella circinata. Fungal Genet. Boil. 1999, 28, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Wilken, P.M.; Steenkamp, E.T.; Wingfield, M.J.; de Beer, Z.W.; Wingfield, B.D. Which MAT gene? Pezizomycotina (Ascomycota) mating-type gene nomenclature reconsidered. Fungal Boil. Rev. 2017, 31, 199–211. [Google Scholar] [CrossRef] [Green Version]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing Ltd.: Oxford, UK, 2006. [Google Scholar]
- Leslie, J.F.; Klein, K.K. Female Fertility and Mating Type Effects on Effective Population Size and Evolution in Filamentous Fungi. Genetics 1996, 144, 557–567. [Google Scholar]
- Berbegal, M.; Pérez-Sierra, A.; Armengol, J.; Grünwald, N.J. Evidence for Multiple Introductions and Clonality in Spanish Populations of Fusarium circinatum. Phytopathology 2013, 103, 851–861. [Google Scholar] [CrossRef] [Green Version]
- Iturritxa, E.; Ganley, R.J.; Wright, J.; Heppe, E.; Steenkamp, E.T.; Gordon, T.R.; Wingfield, M.J. A genetically homogenous population of Fusarium circinatum causes pitch canker of Pinus radiata in the Basque Country, Spain. Fungal Boil. 2011, 115, 288–295. [Google Scholar] [CrossRef] [Green Version]
- Britz, H.; Wingfield, M.J.; Coutinho, T.; Marasas, W.F.O.; Leslie, J.F. Female Fertility and Mating Type Distribution in a South African Population of Fusarium subglutinans f. sp. pini. Appl. Environ. Microbiol. 1998, 64, 2094–2095. [Google Scholar] [CrossRef] [Green Version]
- Britz, H.; Coutinho, T.; Wingfield, B.D.; Marasas, W.F.O.; Wingfield, M. Diversity, and differentiation in two populations of Gibberella circinata in South Africa. Plant Pathol. 2005, 54, 46–52. [Google Scholar] [CrossRef]
- Leslie, J.F. Fungal Vegetative Compatibility. Annu. Rev. Phytopathol. 1993, 31, 127–150. [Google Scholar] [CrossRef]
- Viljoen, A.; Wingfield, M.J.; Gordon, T.R.; Marasas, W.F.O. Genotypic diversity in a South African population of the pitch canker fungus Fusarium subglutinans f.sp. pini. Plant Pathol. 1997, 46, 590–593. [Google Scholar] [CrossRef]
- Santana, Q.C.; Coetzee, M.P.A.; Steenkamp, E.T.; Mlonyeni, O.X.; Hammond, G.N.A.; Wingfield, M.J.; Wingfield, B.D. Microsatellite discovery by deep sequencing of enriched genomic libraries. Biotechniques 2009, 46, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Gordon, T.; Storer, A.; Okamoto, D. Population structure of the pitch canker pathogen, Fusarium subglutinans f. sp. pini, in California. Mycol. Res. 1996, 100, 850–854. [Google Scholar] [CrossRef]
- Muramoto, M. Pitch Canker of Pinus luchuensis in Japan. Plant Dis. 1990, 74, 530. [Google Scholar] [CrossRef]
- Gordon, T.R. Pitch Canker Disease of Pines. Phytopathology 2006, 96, 657–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullett, M.; Perez-Sierra, A.; Armengol, J.; Berbegal, M. Phenotypical and Molecular Characterisation of Fusarium circinatum: Correlation with Virulence and Fungicide Sensitivity. Forests 2017, 8, 458. [Google Scholar] [CrossRef] [Green Version]
- Britz, H.; Couhnho, T.; Gordon, T.; Wingfield, M. Characterisation of the pitch canker fungus, Fusarium circinatum, from Mexico. S. Afr. J. Bot. 2001, 67, 609–614. [Google Scholar] [CrossRef] [Green Version]
- Adamson, K.; Mullett, M.; Solheim, H.; Barnes, I.; Müller, M.M.; Hantula, J.; Vuorinen, M.; Kacergius, A.; Markovskaja, S.; Musolin, D.; et al. Looking for relationships between the populations of Dothistroma septosporum in northern Europe and Asia. Fungal Genet. Boil. 2018, 110, 15–25. [Google Scholar] [CrossRef]
- Laas, M.; Adamson, K.; Drenkhan, R. A look into the genetic diversity of Lecanosticta acicola in northern Europe. Fungal Biol. 2019, 123, 773–782. [Google Scholar] [CrossRef]
- Wallin, J.R.; Waggoner, P.E. The influence of climate on the development and spread of Phytophthora infestans in artificially inoculated potato plots. Plant Dis. Rep. 1950, 190, 19–33. [Google Scholar]
- Hanso, M.; Drenkhan, R. Diplodia pinea is a new pathogen on Austrian pine (Pinus nigra) in Estonia. Plant Pathol. 2009, 58, 797. [Google Scholar] [CrossRef]
- Hanso, M.; Drenkhan, R. Lophodermium needle cast, insect defoliation and growth responses of young Scots pines in Estonia. For. Pathol. 2011, 42, 124–135. [Google Scholar] [CrossRef]
- Hanso, M.; Drenkhan, R. Simple visualization of climate change for improving the public perception in forest pathology/Kliimamuutuste visualiseerimise lihtne viis seoste paremaks tajumiseks metsapatoloogias. For. Stud. 2013, 58, 37–45. [Google Scholar] [CrossRef] [Green Version]
- McDonald, M.J. Temperature effects on Fusarium subglutinans f.sp. pini infection on juvenile Pinus radiata (Monterey pine) and influence on growth of Fusarium subglutinans f.sp. pini isolates from California and Florida. Master’s Thesis, San Jose State University, San Jose, CA, USA, 1994. [Google Scholar]
- Gordon, T.; Kirkpatrick, S.C.; Petersen, J.C.; Friel, C.J. Potential diversity in vegetative compatibility groupings in the California population of Gibberella circinata. Mycol. Res. 2006, 110, 936–940. [Google Scholar] [CrossRef] [PubMed]
- Blank, L.; Martín-García, J.; Bezos, D.; Vettraino, A.M.; Krasnov, H.; Lomba, J.M.; Fernández, M.; Diez, J.J. Factors Affecting the Distribution of Pine Pitch Canker in Northern Spain. Forests 2019, 10, 305. [Google Scholar] [CrossRef] [Green Version]
- Watt, M.; Ganley, R.J.; Kriticos, D.J.; Manning, L.K. Dothistroma needle blight and pitch canker: The current and future potential distribution of two important diseases of Pinus species. Can. J. For. Res. 2011, 41, 412–424. [Google Scholar] [CrossRef]
- Serra-Varela, M.J.; Alía, R.; Pórtoles, J.; Gonzalo, J.; Soliño, M.; Grivet, D.; Raposo, R. Incorporating exposure to pitch canker disease to support management decisions of Pinus pinaster Ait. in the face of climate change. PLoS ONE 2017, 12, e0171549. [Google Scholar] [CrossRef]
- Drenkhan, R.; Tomešová-Haataja, V.; Fraser, S.; Bradshaw, R.E.; Vahalík, P.; Mullett, M.; Martín-García, J.; Bulman, L.S.; Wingfield, M.J.; Kirisits, T.; et al. Global geographic distribution and host range of Dothistroma species: A comprehensive review. For. Pathol. 2016, 46, 408–442. [Google Scholar] [CrossRef]
- New, M.; Lister, D.; Hulme, M.; Makin, I. A high-resolution data set of surface climate over global land areas. Clim. Res. 2002, 21, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high-resolution interpolated climate surfaces for global land areas. Int. J. Clim. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Woo, K.-S.; Koo, Y.-B.; Yeo, J.-K. Variation in susceptibility of six pine species and hybrids to pitch canker caused by Fusarium circinatum. For. Pathol. 2008, 38, 419–428. [Google Scholar] [CrossRef]
- Muramoto, M.; Tashiro, T.; Minamihashi, H. Distribution of Fusarium moniliforme var. subglutinans in Kagoshima Prefecture and its pathogenicity to pines. J. Jpn. For. Soc. 1993, 75, 1–9. [Google Scholar]
- Swett, C.L.; Gordon, T.R. Colonization of corn (Zea mays) by the pitch canker pathogen, Fusarium circinatum: Insights into the evolutionary history of a pine pathogen. Phytopathology 2009, 99, S126–S127. [Google Scholar]
- Guerra-Santos, J.J.; Cibrian-Tovar, D. El Cancro Resinoso por Fusarium subglutinans (Wollenw y Reink) Nelson, Tousson y Marasas, Una nueva Enfermedad de Pinus spp. en Mexico; Revista Chapingo (Mexico). In Serie Ciencias Forestales y del Ambiente; Universidad Autonoma Chapingo: Chapingo, Mexico, 1998; pp. 279–284. [Google Scholar]
- Hodge, G.; Dvorak, W. Differential responses of Central American and Mexican pine species and Pinus radiata to infection by the pitch canker fungus. New For. 2000, 19, 241–258. [Google Scholar] [CrossRef]
- Porter, B.; Wingfield, M.; Coutinho, T. Susceptibility of South African native conifers to the pitch canker pathogen, Fusarium circinatum. S. Afr. J. Bot. 2009, 75, 380–382. [Google Scholar] [CrossRef] [Green Version]
- Vivas, M.; Zas, R.; Solla, A. Screening of Maritime pine (Pinus pinaster) for resistance to Fusarium circinatum, the causal agent of Pitch Canker disease. Forests 2011, 85, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Gordon, T.R.; Okamoto, D.; Storer, A.J.; Wood, D.L. Susceptibility of Five Landscape Pines to Pitch Canker Disease, Caused by Fusarium subglutinans f. sp. pini. HortScience 1998, 33, 868–871. [Google Scholar] [CrossRef] [Green Version]
- Iturritxa, E.; Ganley, R.J.; Raposo, R.; Mesanza, N.; Kirkpatrick, S.C.; Gordon, T.; García-Serna, I. Resistance levels of Spanish conifers against Fusarium circinatum and Diplodia pinea. For. Pathol. 2013, 43, 488–495. [Google Scholar] [CrossRef]
- Dvorak, W.; Hodge, G.; Kietzka, J. Genetic variation in survival, growth, and stem form of Pinus leiophylla in Brazil and South Africa and provenance resistance to pitch canker. South. Hemisphere For. J. 2007, 69, 125–135. [Google Scholar] [CrossRef]
- Hodge, G.R.; Dvorak, W.S. Variation in pitch canker resistance among provenances of Pinus patula and Pinus tecunumanii from Mexico and Central America. New For. 2006, 33, 193–206. [Google Scholar] [CrossRef]
- Dvorak, W.S.; Potter, K.M.; Hipkins, V.D.; Hodge, G. Genetic Diversity and Gene Exchange in Pinus oocarpa, a Mesoamerican Pine with Resistance to the Pitch Canker Fungus (Fusarium circinatum). Int. J. Plant Sci. 2009, 170, 609–626. [Google Scholar] [CrossRef] [Green Version]
- Elvira-Recuenco, M.; Iturritxa, E.; Majada, J.; Alia, R.; Raposo, R. Adaptive Potential of Maritime Pine (Pinus pinaster) Populations to the Emerging Pitch Canker Pathogen, Fusarium circinatum. PLoS ONE 2014, 9, e114971. [Google Scholar] [CrossRef] [PubMed]
- Iturritxa, E.; Mesanza, N.; Elvira-Recuenco, M.; Serrano, Y.; Quintana, E.; Raposo, R. Evaluation of genetic resistance in Pinus to pitch canker in Spain. Australas. Plant Pathol. 2012, 41, 601–607. [Google Scholar] [CrossRef]
- Carey, W.A.; Oak, S.W.; Enebak, S.A. Pitch canker ratings of longleaf pine clones correlate with Fusarium circinatum infestation of seeds and seedling mortality in containers. For. Pathol. 2005, 35, 205–212. [Google Scholar] [CrossRef]
- Enebak, S.A.; Stanosz, G.R. Responses of conifer species of the Great Lakes region of North America to inoculation with the pitch canker pathogen Fusarium circinatum. For. Pathol. 2003, 33, 333–338. [Google Scholar] [CrossRef]
- Broaddus, J.B. Branch Dieback and Cone and Seed Infection Caused by Fusarium moniliforme var. subglutinans in a Loblolly Pine Seed Orchard in South Carolina. Phytopathology 1985, 75, 1104. [Google Scholar] [CrossRef]
- Enebak, S.A.; Carey, W.A. Pitch Canker Caused by Fusarium circinatum Identified on Spruce Pine in Alabama. Plant Dis. 2003, 87, 449. [Google Scholar] [CrossRef]
- Schmale, D.G.; Gordon, T. Variation in susceptibility to pitch canker disease, caused by Fusarium circinatum, in native stands of Pinus muricata. Plant Pathol. 2003, 52, 720–725. [Google Scholar] [CrossRef]
- Gordon, T.; Storer, A.J.; Wood, D.L. The Pitch Canker Epidemic in California. Plant Dis. 2001, 85, 1128–1139. [Google Scholar] [CrossRef]
- Zanoni, T.A.; Farjon, A. World Checklist and Bibliography of Conifers. Brittonia 1999, 51, 76. [Google Scholar] [CrossRef]
- Price, A.; Liston, A.; Strauss, S.H. Evolution, phylogeny, and systematics. In Ecology and Biogeography of Pinus; Richardson, D.M., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 49–68. [Google Scholar]
- García, D.B.; Lomba, J.M.; Martínez-Álvarez, P.; Fernández, M.; Diez, J.J. Effects of Pruning in Monterrey Pine Plantations Affected by Fusarium circinatum. For. Syst. 2012, 21, 481. [Google Scholar] [CrossRef] [Green Version]
- Storer, A.J.; Gordon, T.; Dallara, P.L.; Wood, D. Pitch canker kills pines, spreads to new species and regions. Calif. Agric. 1994, 48, 9–13. [Google Scholar] [CrossRef]
- Fisher, R.F.; Garbett, W.S.; Underhill, E.M. Effects of Fertilization on Healthy and Pitch Canker-Infected Pines. South. J. Appl. For. 1981, 5, 77–79. [Google Scholar] [CrossRef]
- Dwinell, L.D.; Ryan, P.L.; Kuhlman, E.G. Pitch canker of loblolly pine in seed orchards. In Proceedings of the 14th Southern Forest Tree Improvement Conference, Gainesville, FL, USA, 14–16 June 1977; pp. 130–137. [Google Scholar]
- Woo, K.S.; Yoon, J.H.; Han, S.U.; Kim, C.S. Assessment of pathogenic variation against pitch canker pathogen, Fusarium circinatum in Pinus thunbergii and responses of natural selection Pinus × rigitaeda to branch inoculation in a seed orchard. Plant Pathol. J. 2010, 26, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Vivas, M.; Nunes, C.; Coimbra, M.A.; Solla, A. Maternal effects and carbohydrate changes of Pinus pinaster after inoculation with Fusarium circinatum. Trees 2013, 28, 373–379. [Google Scholar] [CrossRef]
- Ford, C.; Jones, N.B.; Chirwa, P.W. Pinus patula and pine hybrid hedge productivity in South Africa: A comparison between two vegetative propagation systems exposed to natural infection by Fusarium circinatum. South. For. J. For. Sci. 2014, 76, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Kanzler, A.; Nel, A.; Ford, C. Development and commercialisation of the Pinus patula x P. tecunumanii hybrid in response to the threat of Fusarium circinatum. New For. 2014, 45, 417–437. [Google Scholar] [CrossRef]
- Dick, M.A.; Simpson, J.A. Fusarium circinatum as a Cause of Damping-off: Exploring the Host Range. Unpublished Report. 2003. Available online: http://www.nzfoa.org.nz/images/stories/pdfs/content/fhrc_reports/2002-04.pdf (accessed on 11 February 2020).
- Fernández-Fernández, M.; Naves, P.; Musolin, D.L.; Selikhovkin, A.V.; Cleary, M.; Chira, D.; Paraschiv, M.; Gordon, T.; Solla, A.; Papazova-Anakieva, I.; et al. Pine Pitch Canker and Insects: Regional Risks, Environmental Regulation, and Practical Management Options. Forests 2019, 10, 649. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Continent/ Country/ State | Year of First Record of FC in Nursery and/or Wider Environment | Host Species | Type of Planting/Sampling Site | Identification Method | References or Data Holder |
|---|---|---|---|---|---|
| Europe | |||||
| France | 20051, 2 | Pseudotsuga menziesii, Pinus sp. | Urban greenery | species-specific PCR, sequence analysis | [37] |
| France | 2008 2 | P. menziesii | Nursery | species-specific PCR, sequence analysis | [44] |
| Italy | 20051, 2 | P. halepensis, P. pinea | Urban greenery | species-specific PCR | [36] |
| Portugal | 20071, 2 | Pinus radiata, P. pinaster | Nursery | species-specific PCR, sequence analysis | [38] |
| Portugal | 2016 2 | P. radiata | Forest plantation | species-specific PCR, sequence analysis | [42] |
| Spain | 19951 | P. radiata, P. halepensis | Nursery, in Galicia | visual observation | [34] |
| Spain | 1997 | P. radiata | Nurseries, in Basque Country | mycelial morphology | [39] |
| Spain | 2003 | P. radiata, P. pinaster | Nursery, in Asturias | species-specific PCR | [35] |
| Spain | 2004 | P. radiata | Plantation, in Cantabria | species-specific PCR | [35] |
| Spain | 2005 | P. sylvestris, P. nigra, P. pinaster, P. pinea | Nurseries in Castilla y León | morphological traits, species-specific PCR | Regional Forest Health Service |
| Spain | 2005 | P. sylvestris, P. nigra, P. pinea, P. radiata | Forest plantations, in Castilla y León | morphological traits, species-specific PCR | Regional Forest Health Service |
| Asia | |||||
| Japan | 19811 | P. luchuensis | Forest, Amamioshima Island (Ryukyu Archipelago) and the Okinawa island | mycelial morphology | [60] |
| South Korea | 19951 | P. rigida | Urban greenery, forest | mycelial morphology | [59] |
| Africa | |||||
| South Africa | 19911 | P. patula | Nursery, Ngodwana Mpumalanga Province | mycelial morphology | [12] |
| South Africa | 2005 | P. radiata | Plantation, Tokai, Western Cape Province | species-specific PCR, sequence analysis | [67] |
| South Africa | 2007 | P. greggii | Plantation, Ugie, Eastern Cape Province | sequence analysis | [68,69] |
| South Africa | 2014 | P. patula | Plantation, Louis Trichardt, Limpopo Province | sequence analysis | [70] |
| North America | |||||
| Haiti | 19531 | P. occidentalis | Natural forest | mycelial morphology | [53] |
| Mexico | 19891 | P. douglasiana, P. halepensis, P. leiophylla, P. greggii, P. patula | Forest plantation and natural stand | unknown, probably visual observation | [40] |
| United States of America | |||||
| Alabama | 1980 | Pinus taeda | Forest plantation | mycelial morphology | [16] |
| Arkansas | 2013 | P. elliottii, P. palustris, P. taeda | unknown | unknown | [78] |
| California | 1986 | P. radiata | Forest plantation | visual observation | [79,80] |
| Florida | 1974 | P. elliottii var. elliottii | Forest plantation and seed orchards | unknown | [17,40] |
| Georgia | 1987 | unknown | Nursery | mycelial morphology | [81] |
| Georgia | 1988 | P. taeda | Forest plantation | visual observation | [82] |
| Indiana | 1994 | P. elliottii var. elliottii | Nursery | mycelial morphology | [83] |
| Louisiana | 2004 | P. taeda | Forest plantation | visual observation | [84] |
| Mississippi | 1974 | P. taeda | Seed orchards | visual observation | [17] |
| North Carolina | 19451 | P. virginiana, P. echinata, P. rigida | Natural forest | visual observation, mycelial morphology | [24] |
| South Carolina | 19451 | P. virginiana | unknown | visual observation, mycelial morphology | [24] |
| Tennessee | 1978 | P. echinata | unknown | visual observation | [85] |
| Texas | 1991 | unknown | Forest plantation | mycelial morphology | [80] |
| Virginia | 1985 | P. echinata | unknown | visual observation | [17,85] |
| South America | |||||
| Brazil | 20141 | P. taeda | Nursery | sequence analysis | [58] |
| Chile | 20011 | P. radiata | Nursery | sequence analysis | [54] |
| Colombia | 20051 | P. maximinoi, P. Patula, P. Tecunumanii | Nursery | species-specific PCR, sequence analysis | [56,57] |
| Colombia | 2006 | P. patula | Forest plantation | sequence analysis | [56,57] |
| Uruguay | 20091 | P. taeda | Nursery | species-specific PCR | [55] |
| Minimum | Average | Maximum | |
|---|---|---|---|
| Altitude | −3 m a.s.l. | 262 m a.s.l. | 3619 m a.s.l. |
| Annual mean temperature | 6.2 °C | 13.7°C | 25.4 °C |
| Mean temperature of the warmest month | 12.4 °C | 24.7 °C | 36.5 °C |
| Mean temperature of the coldest month | −14.2 °C | 4.8 °C | 16.7 °C |
| Annual precipitation sum | 324 mm | 1259 mm | 3062 mm |
| Precipitation sum of the wettest month | 51 mm | 154 mm | 583 mm |
| Precipitation sum of the driest month | 0 mm | 57 mm | 151 mm |
| Susceptibility/ Host Species1 | Common English Names | Subgenus; Section; Subsection2 | Type of Infection | Growth or Test Conditions | Seedlings and/or Plant Age3 | References |
|---|---|---|---|---|---|---|
| Susceptibility high | ||||||
| Pinus brutia Ten. | Turkish pine, Calabrian pine, East Mediteranean pine, Brutia pine | Pinus; Pinus; Halepenses | Artificial | Growth chamber | 2 years | J. Martín-García, unpublished |
| Pinus cembroides Zucc. | Pinyon pine, Mexican nut pine | Strobus; Parrya; Cembroides | Artificial | Greenhouse | Unknown | [138] |
| Pinus douglasiana Martínez | Gordon’s pine, Douglas pine | Pinus; Pinus; Ponderosae; ‘Pseudostrobus Group’ | Artificial | Greenhouse | Unknown | [138] |
| Pinus greggii Engelm. ex Parl. | Gregg’s pine | Pinus; Pinus; Oocarpae; ‘Oocarpa Group’ | Artificial | Field | 2 years | [76] |
| Pinus halepensis Mill. | Aleppo pine | Pinus; Pinus; Halepenses | Artificial | Growth chamber | 2 years | J. Martín-García, unpublished |
| Pinus hartwegii Lindl. | Endlicher pine | Pinus; Pinus; Ponderosae; ‘Montezumae Group’ | Artificial | Greenhouse | Unknown | [138] |
| Pinus herrerae Martínez | Herrera’s pine | Pinus; Pinus; Oocarpae; ‘Teocote Group’ | Artificial | Greenhouse | 12 weeks | [139] |
| Pinus montezumae Lamb. | Montezuma pine | Pinus; Pinus; Ponderosae; ‘Montezumae Group’ | Artificial | Greenhouse | Unknown | [138] |
| Pinus mugo Turra | Mountain pine, dwarf mountain pine | Pinus; Pinus, Pinus | Artificial | Growth chamber | Recently emerged | [9] |
| Pinus mugo Turra subsp. uncinata (Ramond ex DC.) Domin. | Swiss mountain pine | Pinus; Pinus, Pinus | Artificial | Growth chamber | Recently emerged | [8] |
| Pinus nigra J.F.Arnold | Austrian pine, black pine | Pinus; Pinus; Pinus | Artificial | Growth chamber | Recently emerged, 2 years | [8], J. Martín-García, unpublished |
| Pinus patula Schiede ex Schltdl. & Cham. | Patula pine, Jelecote pine, Mexican weeping pine, spreading-leaved pine | Pinus; Pinus; Oocarpae; ‘Oocarpa Group’ | Artificial | Greenhouse | 7–10 m | [71,75,140] |
| Pinus pinaster Aiton | Maritime pine | Pinus; Pinus; Pinus | Artificial | Growth chamber | Recently emerged, 2 years | [8,141], J. Martín-García, unpublished |
| Pinus pseudostrobus Lindl. | Smooth-bark Mexican pine | Pinus; Pinus; Ponderosae; ‘Pseudostrobus Group’ | Artificial | Greenhouse | Unknown | [138] |
| Pinus radiata D. Don | Monterey pine, radiata pine, insignis pine | Pinus; Pinus; Attenuatae | Artificial | Growth chamber, greenhouse, field | Recently emerged, 3 months, 1 year, 2 years, 2–3 years, 3–4 years, unknown | [8,76,80,88,139,142,143] |
| Pinus strobus L. | Eastern white pine, northern white pine, white pine, Weymouth pine (British), and soft pine | Strobus; Strobus;Strobi | Artificial | Growth chamber | Recently emerged | [8] |
| Pinus sylvestris L. | Scots/Scotch pine | Pinus; Pinus; Pinus | Artificial | Growth chamber, greenhouse | Recently emerged, 1.5 years, 2 years | [8,10] J. Martín-García, unpublished |
| Pinus taeda L. | Loblolly pine | Pinus; Pinus; Australes | Artificial | Greenhouse | 1 year | [88] |
| Susceptibility moderate-high | ||||||
| Pinus leiophylla Schiede ex Schltdl. & Cham. | Chihuahua pine, smooth-leaf pine, yellow pine | Pinus; Pinus; Leiophyllae | Artificial | Field | 5–8 years | [144] |
| Pinus patula Schiede ex Schltdl. & Cham. | Patula pine, Jelecote pine, Mexican weeping pine, spreading-leaved pine | Pinus; Pinus; Oocarpae; ‘Oocarpa Group’ | Artificial | Greenhouse, Field | 3 months, 21 weeks, 2 years | [76,145,146] |
| Pinus pinaster Aiton | Maritime pine, cluster pine | Pinus; Pinus; Pinus | Artificial | Greenhouse | 6 months, 3 years | [141,147] |
| Pinus taeda L. | Loblolly pine | Pinus; Pinus; Australes | Artificial | Greenhouse, Field | 21 weeks, 4 years | [88,147] |
| Susceptibility moderate | ||||||
| Pinus banksiana Lamb. | Jack pine, scrub pine | Pinus; Pinus; Contortae | Artificial | Greenhouse | 3.5 years | [148] |
| Pinus devoniana Lindl. | Michoacán pine | Pinus; Pinus; Ponderosae ‘Montezumae Group’ | Artificial | Greenhouse, field | 12 weeks | [97,139] |
| Pinus echinata Mill. | Shortleaf pine | Pinus; Pinus; Australes | Artificial | Greenhouse | 3–4 years | [135] |
| Pinus elliottii Engelm. | Slash pine | Pinus; Pinus; Australes | Artificial | Greenhouse, field | 7–9 months, 8 years | [74,75,144] |
| Pinus greggii Engelm. ex Parl. | Gregg’s pine | Pinus; Pinus; Oocarpae; ‘Oocarpa Group’ | Artificial | Greenhouse | 7 months | [75] |
| Pinus halepensis Mill. | Aleppo pine | Pinus; Pinus; Halepenses | Artificial | Greenhouse | 2 years, 3–4 years, unknown | [138,142,143] |
| Pinus kesiya Royle ex Gordon | Khasia pine, Khasi pine, Benguet pine | Pinus; Pinus; Pinus | Artificial | Field | 2 years | [76] |
| Pinus leiophylla Schiede ex Schltdl. & Cham. | Chihuahua pine, smooth-leaf pine, yellow pine | Pinus; Pinus; Leiophyllae | Artificial | Greenhouse | Unknown | [138] |
| Pinus mugo Turra subsp. uncinata (Ramond ex DC.) Domin. | Swiss mountain pine | Pinus; Pinus, Pinus | Artificial | Greenhouse | 2 years | [148] |
| Pinus nigra J.F.Arnold | Austrian pine, black pine | Pinus; Pinus; Pinus | Artificial | Field | Unknown | [8] |
| Pinus palustris Mill. | Longleaf pine | Pinus; Pinus; Australes | Natural | Nursery | Recently emerged | [149] |
| Pinus pinaster Aiton | Maritime pine, cluster pine | Pinus; Pinus; Pinus | Artificial | Greenhouse, field | 2 years, Unknown | [8,143] |
| Pinus pringlei Shaw | Pringle´s pine | Pinus; Pinus; Oocarpae; ‘Oocarpa Group’ | Artificial | Greenhouse | Unknown | [138] |
| Pinus strobus L. | Eastern white pine, northern white pine, white pine, Weymouth pine (British), and soft pine | Strobus; Strobus;Strobi | Artificial | Greenhouse | 3.5 years | [150] |
| Pinus sylvestris L. | Scots/Scotch pine | Pinus; Pinus; Pinus | Artificial | Greenhouse | 2 years, 3.5 years | [143,150] |
| Pinus tecunumanii F.Schwerdtf. ex Eguiluz & J.P.Perry | Schwerdtfeger’s Pine, Tecun Uman Pine | Pinus; Pinus; Oocarpae; ‘Oocarpa Group’ | Artificial | Greenhouse | 3 months, 12 weeks, 6–8 months (high elevation origin) | [139,145,146] |
| Pinus virginiana Mill. | Virginia pine, Jersey pine, scrub pine | Pinus; Pinus; Contortae | Artificial | Greenhouse | 3–4 years | [135] |
| Susceptibility low-moderate | ||||||
| Pinus caribaea Morelet | Caribbean pine | Pinus; Pinus; Australes | Artificial | Greenhouse | 7 months | [75] |
| Pinus elliottii Engelm. | Slash pine | Pinus; Pinus; Australes | Artificial | Field | 2 years | [76] |
| Pinus mugo Turra subsp. uncinata (Ramond ex DC.) Domin. | Swiss mountain pine | Pinus; Pinus, Pinus | Artificial | Field | Unknown | [8] |
| Pinus pinaster Aiton | Maritime pine, cluster pine | Pinus; Pinus; Pinus | Artificial | Greenhouse | 2 years | [148] |
| Pinus sylvestris L. | Scots/Scotch pine | Pinus; Pinus; Pinus | Artificial | Growth chamber, greenhouse | Recently emerged, 2 years | [9,148] |
| Pinus taeda L. | Loblolly pine | Pinus; Pinus; Australes | Artificial | Greenhouse | 7–9 months, 1 year | [74,151] |
| Pinus thunbergii Parl. | black pine, Japanese black pine, Japanese pine | Pinus; Pinus; Pinus | Artificial | Greenhouse | 3–4 years | [135] |
| Susceptibility low | ||||||
| Pinus ayacahuite Ehrenb. ex Schltdl. | Mexican white pine, ayacahuite pine | Strobus; Strobus; Strobi | Artificial | Greenhouse | Unknown | [138] |
| Pinus canariensis C.Sm. | Canary Island pine | Pinus; Pinus; Canarienses | Artificial | Greenhouse | 2–3 years, 3–4 years | [3,142] |
| Pinus caribaea Morelet | Caribbean pine | Pinus; Pinus; Australes | Artificial | Greenhouse | 3 months | [139] |
| Pinus clausa (Chapm. ex Engelm.) Vasey ex Sarg. | Sand pine, Florida spruce pine, Alabama pine | Pinus; Pinus; Contortae | Artificial, natural | Greenhouse, field | 18 months, 6–8 years | [82] |
| Pinus glabra Walter | Spruce pine | Pinus; Pinus; Australes | Artificial, natural | Greenhouse, field | 5 months, unknown | [152] |
| Pinus jaliscana Perez de la Rosa | Jalisco pine | Pinus; Pinus; Oocarpae; ‘Oocarpa Group’ | Artificial | Greenhouse | 3 months | [139] |
| Pinus luchuensis Mayr | Luchu/Ryukyu pine | Pinus; Pinus; Pinus | Artificial | Greenhouse | 2 years | [136] |
| Pinus maximinoi H.E. Moore | Thinleaf pine | Pinus; Pinus; Ponderosae; ‘Pseudostrobus Group’ | Artificial | Greenhouse | 3 months, 7 months, 8–9 months | [56,74,145] |
| Pinus monophylla Torr. & Frém. | Singleleaf pinyon pine | Strobus; Parrya; Cembroides | Natural | Field | Unknown | [79] |
| Pinus nigra J.F.Arnold | Austrian pine, black pine | Pinus; Pinus; Pinus | Artificial | Greenhouse | 2 years, 3.5 years | [143,150,153] |
| Pinus occidentalis Sw. | Western Indian pine | Pinus; Pinus; Australes | Natural | Field | Unknown | [53] |
| Pinus oocarpa Schiede ex Schltdl. | Ocote pine, Egg-cone pine, hazelnut pine | Pinus; Pinus; Oocarpae; ‘Oocarpa Group’ | Artificial | Greenhouse | 3 months, 7 months | [75,139,145] |
| Pinus pringlei Shaw | Pringle’s pine | Pinus; Pinus; Oocarpae; ‘Oocarpa Group’ | Artificial | Greenhouse | 7 months | [75] |
| Pinus pseudostrobus Lindl. syn. Pinus oaxacana Mirov | Smooth-bark Mexican pine | Pinus; Pinus; Ponderosae; ‘Pseudostrobus Group’ | Natural, artificial | Field, greenhouse | Unknown, 7 months | [74,97] |
| Pinus pungens Lamb. | Table mountain pine | Pinus; Pinus; Australes | Artificial | Unknown | Unknown | [53] |
| Pinus resinosa Aiton | Red pine, Norway pine | Pinus; Pinus; Pinus | Artificial | Greenhouse | 3.5 years | [150] |
| Pinus rigida Mill. | Pitch pine | Pinus; Pinus; Australes | Artificial | Greenhouse | 3–4 years | [135] |
| Pinus serotina Michx. | Pond pine, marsh pine, pocosin pine | Pinus; Pinus; Australes | Artificial | Greenhouse | 1 year | [53,151] |
| Pinus sylvestris L. | Scots/Scotch pine | Pinus; Pinus; Pinus | Artificial | Field | 2 years, Unknown | [8] |
| Pinus taeda L. | Loblolly pine | Pinus; Pinus; Australes | Artificial | Field | 8 years | [140] |
| Pinus tecunumanii F.Schwerdtf. ex Eguiluz & J.P.Perry | Schwerdtfeger’s Pine, Tecun Uman Pine | Pinus; Pinus; Oocarpae; ‘Oocarpa Group’ | Artificial | Greenhouse, field | 3 months, 12 weeks, 6–8 months, 8 years (low-elevation origin) | [56,74,139,144,145,146] |
| Pinus thunbergii Parl. | Black pine, Japanese black pine, Japanese pine | Pinus; Pinus; Pinus | Artificial | Greenhouse | 3–4 years | [142] |
| Resistant | ||||||
| Pinus koraiensis Siebold and Zucc. | Korean pine | Strobus; Strobus; Cembrae | Artificial | Greenhouse | 3 years | [59] |
| Highly variable susceptibility | ||||||
| Pinus densiflora Siebold and Zucc. | Japanese red pine, Korean red pine | Pinus; Pinus; Pinus | Artificial | Greenhouse | 3–4 years | [59,135,136] |
| Pinus muricata D. Don | Bishop pine | Pinus; Pinus; Attenuatae; | Artificial | Greenhouse | Unknown | [79] |
| Pinus pinea L. | Italian stone pine, umbrella pine | Pinus; Pinus; Pineae | Artificial | Greenhouse, growth chambers | 6 months, 2 years, 3-4 years | [35,37,143,154], J. Martín-García, unpublished |
| Susceptible, unknown susceptibility rate | ||||||
| Pinus attenuata Lemmon | Knobcone pine, Narrowcone pine | Pinus; Pinus; Attenuatae | Artificial | Greenhouse | Unknown | [79,154] |
| Pinus brutia Ten. var. eldarica (Medw.) Silba | Eldarica pine | Pinus; Pinus; Halepenses | Artificial | Greenhouse | Unknown | [79] |
| Pinus canariensis C.Sm. | Canary Island pine | Pinus; Pinus; Canarienses | Artificial | Greenhouse | Unknown | [79] |
| Pinus contorta Douglas ex Loudon | Shore pine, lodgepole pine | Pinus; Pinus; Contortae | Artificial | Greenhouse | Unknown | [141] |
| Pinus coulteri D. Don | Coulter pine, big-cone pine | Pinus; Pinus; Ponderosae; ‘Sabinianae Group’ | Artificial | Greenhouse | Unknown | [79,154] |
| Pinus halepensis Mill. | Aleppo pine | Pinus; Pinus; Halepenses | Artificial | Nursery | 2 years | [35] |
| Pinus jeffreyi A.Murray bis | Jeffrey pine | Pinus; Pinus; Ponderosae | Artificial | Greenhouse | Unknown | [79] |
| Pinus lambertiana Douglas | Sugar pine | Strobus; Strobus; Strobi | Artificial | Greenhouse | Unknown | [79] |
| Pinus nigra J.F. Arnold subsp. laricio (Poir.) Maire | Corsican pine | Pinus; Pinus; Pinus | Natural | Nursery | <6 months | A.V. Sanz-Ros, unpublished |
| Pinus pinaster Aiton | Maritime pine, cluster pine | Pinus; Pinus; Pinus | Artificial | Nursery | 2 years | [36] |
| Pinus ponderosa Douglas ex Loudon | Ponderosa pine, bull pine, blackjack pine | Pinus; Pinus; Ponderosae | Artificial | Greenhouse | Unknown | [79] |
| Pinus sabiniana Douglas ex D. Don | Gray/foothill/digger pine | Pinus; Pinus; Ponderosae; ‘Sabinianae Group’ | Artificial | Greenhouse | Unknown | [79] |
| Susceptibility/ Host Species1 | Sampling Site | Status of Host | Tree Age, Years | Reference |
|---|---|---|---|---|
| Susceptibility high | ||||
| Pinus radiata D. Don | Plantation, unknown | Exotic, native | Unknown, 20y | [12,34,128,154,157] |
| Susceptibility low-moderate | ||||
| Pinus discolor D.K. Bailey & Hawksw. | Unknown | Native | Unknown | [138] |
| Pinus douglasiana Martinéz | Unknown | Native | Unknown | [138] |
| Pinus durangensis Martinéz | Unknown | Native | Unknown | [138] |
| Pinus halepensis Mill. | Unknown | Exotic | Unknown | [138] |
| Pinus leiophylla Schiede ex Schltdl. & Cham. | Unknown | Native | Unknown | [138] |
| Susceptibility low | ||||
| Pinus ayacahuite Ehrenb. ex Schltdl. | Unknown | Native | Unknown | [138] |
| Pinus canariensis C. Sm. | Urban trees | Exotic | Unknown | [154] |
| Pinus luchuensis Mayr | Greenhouse | Native | 11–19 years | [136] |
| Pinus pinaster Aiton | Plantation | Native | Unknown | |
| Highly variable susceptibility | ||||
| Pinus densiflora Siebold and Zucc. | Unknown | Native | Unknown | [136] |
| Pinus muricata D. Don | Unknown, Plantation | Native | Unknown, 12-13 years | [80,153] |
| Pinus pinea L. | Plantation, urban trees | Exotic, native | Unknown | [35,154] |
| Susceptible, unknown susceptibility rate | ||||
| Pinus arizonica Engelm. | Plantation, natural forest | Native | Unknown | [43,97,138] |
| Pinus armandii Franch. | Natural forest | Native | Unknown | [136] |
| Pinus attenuata Lemm | Unknown | Native | Unknown | [154,158] |
| Pinus canariensis C. Sm. | Unknown | Exotic | Unknown | [15,80] |
| Pinus cembroides Zucc. | Unknown | Native | Unknown | [138] |
| Pinus contorta Douglas ex Loudon | Unknown | Native | Unknown | [154] |
| Pinus contorta Douglas ex Loudon var. contorta | Natural forest | Native | Unknown | [158] |
| Pinus coulteri D. Don | Unknown | Native | Unknown | [154] |
| Pinus elliottii Engelm. var. densa (Little and Dorman) E. Murray | Plantation | Native | Unknown | [17] |
| Pinus elliottii Engelm. var. elliottii | Plantation, natural forest | Native, exotic | Unknown | [13,159] |
| Pinus greggii Engelm. ex Parl. | Unknown | Exotic | Unknown | [74,76] |
| Pinus halepensis Mill. | Unknown, urban trees | Exotic, native | Unknown | [35,80] |
| Pinus hartwegii Lindl. | Plantation and natural forest | Native | Unknown | [40,138] |
| Pinus kesiya Royle. ex Gordon | Plantation | Exotic | Unknown | [56] |
| Pinus leiophylla Schiede ex Schltdl. & Cham. var. leiophylla | Natural forest | Native | Unknown | [120] |
| Pinus montezumae Lamb. | Unknown | Native | Unknown | [138] |
| Pinus patula Schiede ex Schltdl. & Cham. | Plantation | Exotic | Unknown | [56] |
| Pinus ponderosa Douglas ex Lawson | Plantation | Native | Unknown | [154] |
| Pinus pringlei Shaw | Unknown | Native | Unknown | [138] |
| Pinus pseudostrobus Lindl. | Unknown | Native | Unknown | [138] |
| Pinus pseudostrobus Lindl. var. apulcensis (Lindl.) Shaw | Plantation and natural forest | Native | Unknown | [40,97,138] |
| Pinus radiata × attenuata | Natural forest | Native | Unknown | [158] |
| Pinus sabiniana Douglas ex D. Don | Unknown | Native | Unknown | [154,158] |
| Pinus taeda L. | Unknown, greenhouse | Native | Unknown, 21 years | [151,160] |
| Pinus thunbergii Parl. | Roadside, golf course | Native | Unknown | [161] |
| Pinus torreyana Parry ex Carrière | Unknown | Native | Unknown | [154,158] |
| Susceptibility/ Host Species1 | Sampling Site | Type of Infection | Plants Stage Tested | Reference |
|---|---|---|---|---|
| Susceptibility moderate-high | ||||
| Pinus greggii × Pinus tecunumanii | Field | Artificial | Established plantation trees | [76] |
| Pinus patula × Pinus greggii | Field | Artificial | Established plantation trees | [76] |
| Pinus patula × Pinus greggii var. australis (from southern Mexico) | Greenhouse | Artificial | Nursery plants established from rooted cuttings | [75] |
| Pinus patula × Pinus radiata | Field | Artificial | Established plantation trees | [76] |
| Susceptibility moderate | ||||
| Pinus patula × Pinus greggii var. greggii | Nursery | Natural | Nursery plants established from rooted cuttings | [163] |
| Pinus patula × Pinus pringlei | Greenhouse | Artificial | Nursery plants established from rooted cuttings | [75] |
| Pinus patula × Pinus tecunumanii (High elevation source) | Greenhouse | Artificial | Nursery plants established from rooted cuttings | [75] |
| Pinus patula × Pinus tecunumanii | Field | Artificial | Established plantation trees | [76] |
| Susceptibility low-moderate | ||||
| Pinus patula × Pinus elliottii | Greenhouse | Artificial | Nursery plants established from rooted cuttings | [75] |
| Pinus patula × Pinus tecunumanii (High elevation source) | Nursery | Natural | Nursery plants established from rooted cuttings | [163] |
| Pinus rigida × Pinus taeda | Greenhouse | Artificial | Nursery plants established from rooted cuttings | [135] |
| Susceptibility low | ||||
| Pinus caribaea × Pinus oocarpa | Greenhouse | Artificial | Nursery plants established from rooted cuttings | [75] |
| Pinus elliottii × Pinus caribaea | Field, greenhouse | Artificial | Established plantation trees, nursery plants established from rooted cuttings | [75,76] |
| Pinus elliottii × Pinus taeda | Greenhouse | Artificial | Nursery plants established from rooted cuttings | [75] |
| Pinus elliottii × Pinus tecunumanii | Greenhouse | Artificial | Nursery plants established from rooted cuttings | [75] |
| Pinus patula × Pinus caribaea var. hondurensis | Nursery | Natural | Nursery plants established from rooted cuttings | [163] |
| Pinus patula × Pinus greggii var. australis | Nursery | Natural | Nursery plants established from rooted cuttings | [163] |
| Pinus patula × Pinus oocarpa | Field, greenhouse | Artificial | Established plantation trees, nursery plants established from rooted cuttings | [75,76] |
| Pinus patula × Pinus tecunumanii | Field | Artificial | Established plantation trees | [76] |
| Pinus patula × Pinus tecunumanii (Low elevation source) | Greenhouse, nursery | Artificial, natural | Nursery plants established from rooted cuttings | [75,163] |
| Pinus taeda × Pinus tecunumanii | Field | Artificial | Established plantation trees | [76] |
| Pinus tecunumanii (High elevation source) × Pinus caribaea | Greenhouse | Artificial | Nursery plants established from rooted cuttings | [75] |
| Pinus tecunumanii (High elevation source) × Pinus oocarpa | Greenhouse | Artificial | Nursery plants established from rooted cuttings | [75] |
| Pinus tecunumanii (Low elevation source) × Pinus caribaea | Greenhouse | Artificial | Nursery plants established from rooted cuttings | [75,164] |
| Susceptibility/Host Species 1 | Common English Names | Type of Infection | Growth or Test Conditions | Host Age, Years | References |
|---|---|---|---|---|---|
| Susceptibility low | |||||
| Larix kaempferi (Lamb.) Carr. | Japanese larch | Artificial | Greenhouse | unknown | [3] |
| Libocedrus decurrens (Torr.) Florin | Incense cedar | Artificial | Greenhouse | unknown | [8] |
| Pseudotsuga menziesii (Mirb.) Franco | Douglas-fir | Artificial, natural | Greenhouse, field | unknown | [3] |
| Resistant | |||||
| Cedrus atlantica (Endl.) G.Manetti ex Carrière | Atlas cedar | Artificial | Greenhouse | Unknown | [8] |
| Chamaecyparis lawsoniana (A. Murray) Parl. | Port Orford cedar, Lawson cypress | Artificial | Greenhouse | Unknown | [8] |
| Cupressocyparis × leylandii (A.B.Jacks. & Dallim.) Dallim. | Leyland cypress | Artificial | Greenhouse | Unknown | [8] |
| Cupressus macrocarpa Hartw. ex Gordon | Monterey cypress | Artificial | Greenhouse | Recently emerged | [165] |
| Eucalyptus regnans F. Muell. | Mountain ash, swamp gum | Artificial | Greenhouse | Recently emerged | [165] |
| Eucalyptus globulus Labill. | Australian fever tree, southern blue gum | Artificial | Greenhouse | Recently emerged | [165] |
| Picea abies (L.) H. Karst. | Norway spruce | Artificial | Greenhouse | 2 years | [10] |
| Picea jezoensis Carrière | Dark-bark spruce | Artificial | Greenhouse | Unknown | [3] |
| Picea sitchensis (Bong.) Carrière | Sitka spruce | Artificial | Greenhouse | Unknown | [3] |
| Podocarpus latifolia (Thunb.) R.Br. ex Mirb. | Broad-leaved yellowwood, real yellowwood | Artificial | Greenhouse | Unknown | [140] |
| Podocarpus elongatus Aiton L´Hérit. ex Pers. | Breede River yellowwood | Artificial | Greenhouse | Unknown | [140] |
| Podocarpus henkelii Stapf ex Dallim. & A.B.Jacks. | Henkel’s yellowwood | Artificial | Greenhouse | Unknown | [140] |
| Sequoia sempervirens (D. Don) Endl. | Coast redwood | Artificial | Greenhouse | Unknown | [3] |
| Sequoiadendron giganteum (Lindl.) J.Buchh. | Giant sequoia | Artificial | Greenhouse | Unknown | [3,8] |
| Thuja plicata Donn ex D. Don | Western Redcedar | Artificial | Greenhouse | Unknown | [8] |
| Widdringtonia schwartzii (Marloth) Mast. | Willowmore Cedar, Baviaans Cedar, Willowmore Cypress | Artificial | Greenhouse | Unknown | [140] |
| Widdringtonia cederbergensis (Marsh) | Clanwilliam cedar, Cape cedar | Artificial | Greenhouse | Unknown | [140] |
| Widdringtonia nodiflora (L.) Powrie | Mountain cedar, mountain cypress | Artificial | Greenhouse | Unknown | [140] |
| Resistant herbs | |||||
| Gladiolus L. | Gladius, a sword, gladioli | Artificial | Greenhouse | Corms | [88] |
| Lolium perenne L. | Perennial ryegrass | Artificial | Greenhouse | Recently emerged | [165] |
| Trifolium repens L. | White clover | Artificial | Greenhouse | Recently emerged | [165] |
| Highly variable susceptibility | |||||
| Abies alba Mill. | European silver fir | Artificial | Greenhouse | Recently emerged, unknown | [8] |
| Larix decidua Mill. | European larch | Artificial | Greenhouse | Recently emerged, 2 years | [8,10] |
| Picea glauca (Moench) Voss | White spruce | Artificial | Greenhouse | Recently emerged, unknown | [3,8,9] |
| Susceptible, unknown susceptibility rate | |||||
| Agrostis capillaris L. | Common bent, colonial bent, or browntop | Natural | Plantation of P. radiata | Unknown | [7] |
| Briza maxima L. | Big quaking grass, great quaking grass | Natural | Nursey | Unknown | [6] |
| Centaurea debeauxii Godr. & Gren. | Meadow knapweed | Natural | Plantation of P. radiata | Unknown | [7] |
| Cymbidium sp. Sw. | Boat orchids | Natural | Unknown | Unknown | [26] |
| Ehrharta erecta Lamb. var. erecta (Hochst.) Pilg. | Panic veldtgrass | Natural | Nursey | Unknown | [6] |
| Festuca arundinacea Schreb. | Tall fescue | Natural | Field | Unknown | [5] |
| Holcus lanatus L. | Common velvet grass | Natural | Field | Unknown | [5] |
| Hypochaeris radicata L. | Cat´s ear | Natural | Plantation of P. radiata | Unknown | [7] |
| Musa acuminata Colla | Wild banana | Natural | Unknown | Unknown | [26] |
| Pentameris pallida (Thunb.) Galley & H.P.Linder | Pussy tail grass | Natural | Nursey | Unknown | [6] |
| Pseudarrhenatherum longifolium (Thore) Rouy | Oat grass | Natural | Plantation of P. radiata | Unknown | [7] |
| Rubus ulmifolius Schott | Elmleaf blackberry | Natural | Plantation of P. radiata | Unknown | [7] |
| Sonchus oleraceus L. | Common sowthistle | Natural | Plantation of P. radiata | Unknown | [7] |
| Zea mays L. | Maize | Natural | Unknown | Unknown | [137] |
| Teucrium scorodonia L. | Woodland germander, wood sage | Natural | Plantation of P. radiata | Unknown | [7] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drenkhan, R.; Ganley, B.; Martín-García, J.; Vahalík, P.; Adamson, K.; Adamčíková, K.; Ahumada, R.; Blank, L.; Bragança, H.; Capretti, P.; et al. Global Geographic Distribution and Host Range of Fusarium circinatum, the Causal Agent of Pine Pitch Canker. Forests 2020, 11, 724. https://doi.org/10.3390/f11070724
Drenkhan R, Ganley B, Martín-García J, Vahalík P, Adamson K, Adamčíková K, Ahumada R, Blank L, Bragança H, Capretti P, et al. Global Geographic Distribution and Host Range of Fusarium circinatum, the Causal Agent of Pine Pitch Canker. Forests. 2020; 11(7):724. https://doi.org/10.3390/f11070724
Chicago/Turabian StyleDrenkhan, Rein, Beccy Ganley, Jorge Martín-García, Petr Vahalík, Kalev Adamson, Katarína Adamčíková, Rodrigo Ahumada, Lior Blank, Helena Bragança, Paolo Capretti, and et al. 2020. "Global Geographic Distribution and Host Range of Fusarium circinatum, the Causal Agent of Pine Pitch Canker" Forests 11, no. 7: 724. https://doi.org/10.3390/f11070724
APA StyleDrenkhan, R., Ganley, B., Martín-García, J., Vahalík, P., Adamson, K., Adamčíková, K., Ahumada, R., Blank, L., Bragança, H., Capretti, P., Cleary, M., Cornejo, C., Davydenko, K., Diez, J. J., Lehtijärvi, H. T. D., Dvořák, M., Enderle, R., Fourie, G., Georgieva, M., ... Mullett, M. S. (2020). Global Geographic Distribution and Host Range of Fusarium circinatum, the Causal Agent of Pine Pitch Canker. Forests, 11(7), 724. https://doi.org/10.3390/f11070724