Effects of Skidding Operations after Tree Harvesting and Soil Scarification by Felled Trees on Initial Seedling Emergence of Spanish Black Pine (Pinus nigra Arn. ssp. salzmannii)

 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
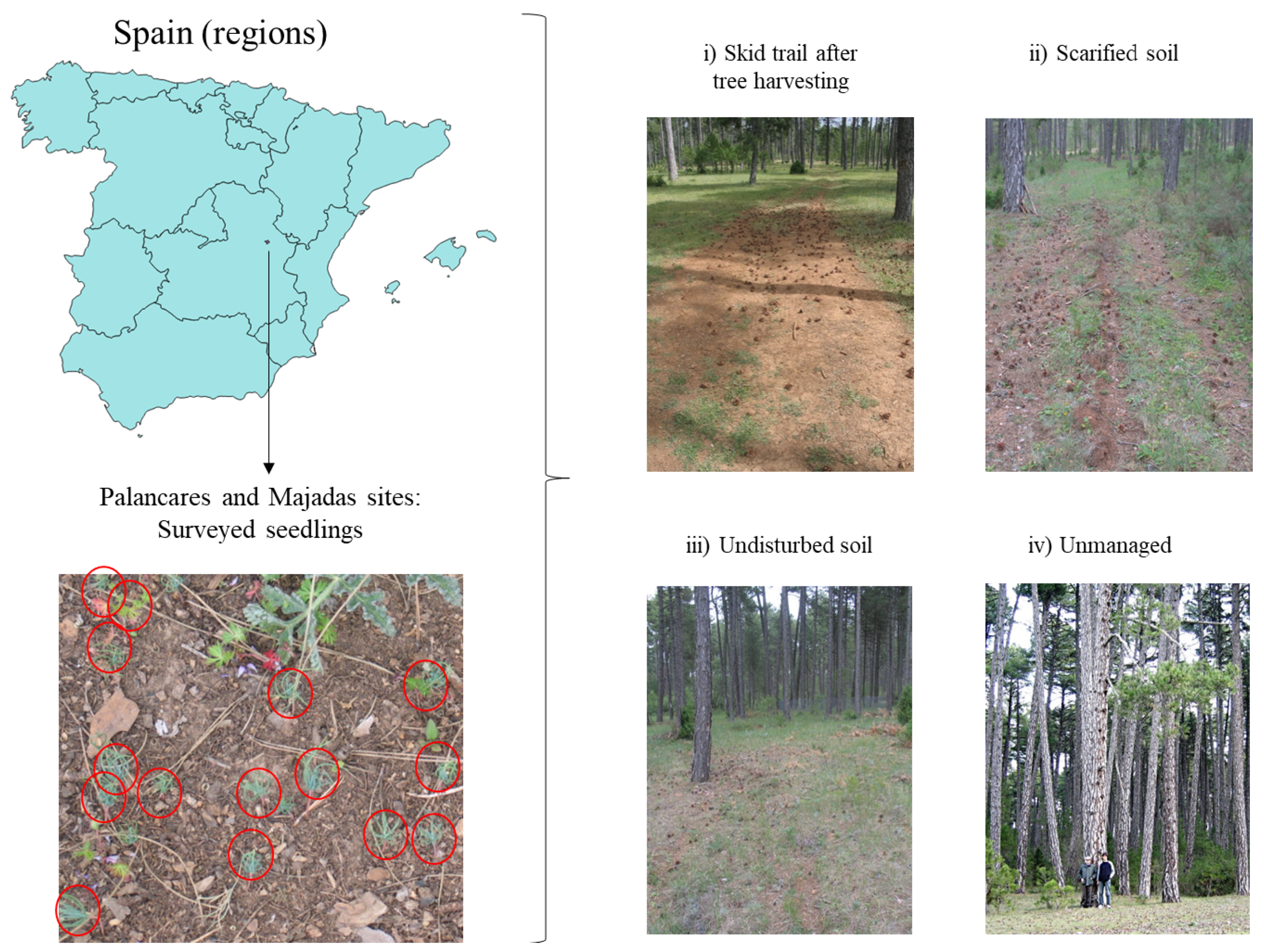
2.1. Study Area
2.2. Experimental Design
2.3. Characterization of the Forests and Soils
2.4. Seed Dispersal and Seedling Emergence Sampling
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oliver, C.D.; Larson, B.C.; Oliver, C.D. Forest Stand Dynamics; John Wiley & Sons: New York, NY, USA, 1996; p. 520. [Google Scholar]
- Ice, G.G.; Neary, D.G.; Adams, P.W. Effects of wildfire on soils and watershed processes. J. For. 2004, 102, 16–20. [Google Scholar]
- Leverkus, A.B.; Lindenmayer, D.B.; Thorn, S.; Gustafsson, L. Salvage Logging in the World’s Forests: Interactions between Natural Disturbance and Logging Need Recognition. Glob. Ecol. Biogeogr. 2018, 27, 1140–1154. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Ortega, R.; Miralles, I.; Plaza-Álvarez, P.A.; González-Romero, J.; Peña-Molina, E.; Moya, D.; Zema, D.A.; Wagenbrenner, J.W.; Heras, J.D.L. Effects of Wildfire and Logging on Soil Functionality in the Short-Term in Pinus Halepensis M. For. Eur. J. For. Res. 2020, 6, 2414. [Google Scholar] [CrossRef]
- Donato, D.C.; Fontaine, J.B.; Campbell, J.L.; Robinson, W.D.; Kauffman, J.B.; Law, B.E. Post-Wildfire Logging Hinders Regeneration and Increases. Fire Risk Sci. 2006, 311, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boucher, Y.; Grondin, P.; Auger, I. Land Use History (1840–2005) and Physiography as Determinants of Southern Boreal Forests. Landsc. Ecol. 2013, 29, 437–450. [Google Scholar] [CrossRef] [Green Version]
- Knapp, E.E.; Ritchie, M.W. Response of Understory Vegetation to Salvage Logging Following a High-Severity Wildfire. Ecosphere 2016, 7, e01550. [Google Scholar] [CrossRef] [Green Version]
- Wagenbrenner, J.W.; Macdonald, L.H.; Coats, R.N.; Robichaud, P.R.; Brown, R.E. Effects of Post-Fire Salvage Logging and a Skid Trail Treatment on Ground Cover, Soils, and Sediment Production in the Interior Western United States. Forest Ecol. Manag. 2015, 335, 176–193. [Google Scholar] [CrossRef]
- Prats, S.A.; Malvar, M.C.; Coelho, C.O.A.; Wagenbrenner, J.W. Hydrologic and erosion responses to compaction and added surface cover in post-fire logged areas: Isolating splash, interrill and rill erosion. J. Hydrol. 2019, 575, 408–419. [Google Scholar] [CrossRef]
- Kozlowski, T. Physiological Ecology of Natural Regeneration of Harvested and Disturbed Forest Stands: Implications for Forest Management. For. Ecol. Manag. 2002, 158, 195–221. [Google Scholar] [CrossRef]
- Calama, R.; Manso, R.; Lucas Borja, M.E.; Espelta, J.M.; Piqué, M.; Bravo Oviedo, F.; Peso Taranco, C.E.D.; Pardos, M. Natural regeneration in Iberian pines: A review of dynamic processes and proposals for management. For. Syst. 2017, 26, 1–20. [Google Scholar] [CrossRef]
- Modrý, M.; Hubený, D.; Rejšek, K. Differential Response of Naturally Regenerated European Shade Tolerant Tree Species to Soil Type and Light Availability. For. Ecol. Manag. 2004, 188, 185–195. [Google Scholar] [CrossRef]
- Neeman, G.; Goubitz, S.; Nathan, R. Reproductive Traits of Pinus halepensis in the Light of Fire–a Critical Review. Plant Ecol. 2004, 171, 69–79. [Google Scholar] [CrossRef]
- Vergarechea, M.; del Río, M.; Gordo, J.; Martín, R.; Cubero, D.; Calama, R. Spatio-temporal variation of natural regeneration in Pinus pinea and Pinus pinaster Mediterranean forests in Spain. Eur. J. For. Res. 2019, 138, 313–326. [Google Scholar] [CrossRef]
- Lucas-Borja, M.; Fonseca, T.; Linares, J.; Morote, F.G.; Serrano, F.L. Does the Recruitment Pattern of Spanish Black Pine (Pinus nigra Arn. ssp. salzmannii) Change the Regeneration Niche over the Early Life Cycle of Individuals? For. Ecol. Manag. 2012, 284, 93–99. [Google Scholar] [CrossRef]
- Prévosto, B.; Amandier, L.; Quesney, T.; Boisgelin, G.D.; Ripert, C. Regenerating Mature Aleppo Pine Stands in Fire-Free Conditions: Site Preparation Treatments Matter. For. Ecol. Manag. 2012, 282, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Herrera, C.M. Historical Effects and Sorting Processes as Explanations for Contemporary Ecological Patterns: Character Syndromes in Mediterranean Woody Plants. Am. Nat. 1992, 140, 421–446. [Google Scholar] [CrossRef] [Green Version]
- Prévosto, B.; Gavinet, J.; Ripert, C.; Fernandez, C. Identification of Windows of Emergence and Seedling Establishment in a Pine Mediterranean Forest under Controlled Disturbances. Basic Appl. Ecol. 2015, 16, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Tavankar, F. Plant species recovery and natural tree regeneration on skid trails in the Hyrcanian forests of Iran. J. Biodivers. Environ. Sci. 2013, 3, 10–16. [Google Scholar]
- Gomez, A.; Powers, R.F.; Singer, M.J.; Horwath, W.R. Soil Compaction Effects on Growth of Young Ponderosa Pine Following Litter Removal in Californias Sierra Nevada. Soil Sci. Soc. Am. J. 2002, 66, 1334–1343. [Google Scholar] [CrossRef]
- Bassett, I.E.; Simcock, R.C.; Mitchell, N.D. Consequences of Soil Compaction for Seedling Establishment: Implications for Natural Regeneration and Restoration. Austral Ecol. 2005, 30, 827–833. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Fonseca, T.F.; Lousada, J.L.; Silva-Santos, P.; Garcia, E.M.; Abellán, M.A. Natural Regeneration of Spanish Black Pine [Pinus nigra Arn. ssp. salzmannii (Dunal) Franco] at Contrasting Altitudes in a Mediterranean Mountain Area. Ecol. Res. 2012, 27, 913–921. [Google Scholar] [CrossRef]
- Aleksandrowicz-Trzcińska, M.; Drozdowski, S.; Studnicki, M.; Żybura, H. Effects of Site Preparation Methods on the Establishment and Natural-Regeneration Traits of Scots Pines (Pinus sylvestris L.) in Northeastern Poland. Forests 2018, 9, 717. [Google Scholar] [CrossRef] [Green Version]
- Lucas-Borja, M.E.; González-Romero, J.; Plaza-Álvarez, P.; Sagra, J.; Gómez, M.; Moya, D.; Cerdà, A.; Heras, J.D.L. The Impact of Straw Mulching and Salvage Logging on Post-Fire Runoff and Soil Erosion Generation under Mediterranean Climate Conditions. Sci. Total Environ. 2019, 654, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Del Cerro Barja, A.; Lucas-Borja, M.E.; García, E.M.; Serrano, F.R.L.; Abellán, M.A.; Morote, F.A.G.; López, R.N. Influence of Stand Density and Soil Treatment on the Spanish Black Pine (Pinus nigra Arn. ssp. salzmannii) Regeneration in Spain. For. Syst. 2009, 18, 167. [Google Scholar]
- Fleming, R.L.; Black, T.A.; Adams, R.S.; Stathers, R.J. Silvicultural treatments, microclimatic conditions and seedling response in Southern interior clearcuts. Can. J. Forest Res. 1998, 78, 115–126. [Google Scholar] [CrossRef]
- Harper, J.L. Population Biology of Plants; Blackburn Press: Caldwell, NJ, USA; London, UK, 2010; p. 892. [Google Scholar]
- Soil Survey Staff, Soil Taxonomy. A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; USDA Natural Resources Conservation Service: Washington, DC, USA, 1999; p. 869.
- Lucas-Borja, M.E.; Candel-Pérez, D.; Morote, F.A.G.; Onkelinx, T.; Tíscar, P.A.; Balandier, P. Pinus nigra Arn. ssp. salzmannii Seedling Recruitment Is Affected by Stand Basal Area, Shrub Cover and Climate Interactions. Ann. For. Sci. 2016, 73, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Fortin, M.; DeBlois, J. Modeling Tree Recruitment with Zero-Inflated Models: The Example of Hardwood Stands in Southern Québec, Canada. For. Sci. 2007, 53, 529–539. [Google Scholar]
- Manso, R.; Ligot, G.; Fortin, M. A recruitment model for beech–oak pure and mixed stands in Belgium, Forestry. Int. J. For. Res. 2020, 93, 124–132. [Google Scholar]
- Zeileis, A.; Kleiber, C.; Jackman, S. Regression Models for Count Data in R. J. Stat. Softw. 2008, 27, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Schupp, E.W. Annual Variation in Seedfall, Postdispersal Predation, and Recruitment of a Neotropical Tree. Ecology 1990, 71, 504–515. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Vacchiano, G. Interactions between Climate, Growth and Seed Production in Spanish Black Pine (Pinus nigra Arn. ssp. salzmannii) Forests in Cuenca Mountains (Spain). New For. 2018, 49, 399–414. [Google Scholar] [CrossRef] [Green Version]
- Lucas-Borja, M.E.; Fonseca, T.; Parresol, B.R.; Silva-Santos, P.; García-Morote, F.A.; Tíscar-Oliver, P.A. Modelling Spanish Black Pine Seedling Emergence: Establishing Management Strategies for Endangered Forest Areas. For. Ecol. Manag. 2011, 262, 195–202. [Google Scholar] [CrossRef]
- Shmida, A.; Ne’eman, G.; Goubitz, S.; Lev-Yadun, S. Sexual Allocation and Gender Segregation in Pinus halepensis, P. brutia and P. pinea. In Ecology, Biogeography and Management of Pinus halepensis and P. brutia Forest Ecosystems in the Mediterranean Basin; Ne’eman, G., Trabaud, L., Eds.; Backhuys Publishers: Leiden, Holland, 2000; pp. 91–104. [Google Scholar]
- Richardson, D.M.; Allsopp, N.; Dantonio, C.M.; Milton, S.J.; Rejmánek, M. Plant Invasions-the Role of Mutualisms. Biol. Rev. 2007, 75, 65–93. [Google Scholar] [CrossRef]
- Nilsson, M.C.; Zackrisson, O.; Sterner, O.; Wallstedt, A. Characterisation of the Differential Interference Effects of Two Boreal Dwarf Shrub Species. Oecologia 2000, 123, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Quilichini, A.; Debussche, M. Seed Dispersal and Germination Patternsin a Rare Mediterranean Island Endemic (Anchusa crispa Viv., Boraginaceae). Acta Oecol. 2000, 21, 303–313. [Google Scholar] [CrossRef]
- Ritari, A.; Ladhe, E. Effect of site preparation on physical properties of the soil in a thick-humus spruce stand. Comm. Inst. For. Fenn. 1978, 92, 1–36. [Google Scholar]
- Örlander, G.; Rosvall-Åhnebrink, G. Evaluating Seedling Quality by Determining Their Water Status. Scand. J. For Res. 1987, 2, 167–177. [Google Scholar]
- Bassman, J.H. Influence of two site preparation treatments on ecophysiology of planted Picea engelmannii× glauca seedlings. Can. J. For. Res. 1989, 19, 1359–1370. [Google Scholar] [CrossRef]
- Flint, L.; Childs, S. Effect of Shading, Mulching and Vegetation Control on Douglas-Fir Seedling Growth and Soil Water Supply. For. Ecol. Manag. 1987, 18, 189–203. [Google Scholar] [CrossRef]
- Martín-Alcón, S.; Coll, L. Unraveling the Relative Importance of Factors Driving Post-Fire Regeneration Trajectories in Non-Serotinous Pinus nigra Forests. For. Ecol. Manag. 2016, 361, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Agestam, E.; Ekö, P.M.; Nilsson, U.; Welander, N. The Effects of Shelterwood Density and Site Preparation on Natural Regeneration of Fagus Sylvatica in Southern Sweden. For. Ecol. Manag. 2003, 176, 61–73. [Google Scholar] [CrossRef]
- Pötzelsberger, E.; Hasenauer, H. Soil Change after 50years of Converting Norway Spruce Dominated Age Class Forests into Single Tree Selection Forests. For. Ecol. Manag. 2015, 338, 176–182. [Google Scholar] [CrossRef]
- Heydari, M.; Prévosto, B.; Abdi, T.; Mirzaei, J.; Mirab-Balou, M.; Rostami, N.; Khosravi, M.; Pothier, D. Establishment of Oak Seedlings in Historically Disturbed Sites: Regeneration Success as a Function of Stand Structure and Soil Characteristics. Ecol. Eng. 2017, 107, 172–182. [Google Scholar] [CrossRef]
- Heydari, M.; Prévosto, B.; Naji, H.R.; Mehrabi, A.A.; Pothier, D. Influence of Soil Properties and Burial Depth on Persian Oak (Quercus brantii Lindl.) Establishment in Different Microhabitats Resulting from Traditional Forest Practices. Eur. J. For. Res. 2017, 136, 287–305. [Google Scholar] [CrossRef] [Green Version]
- Keim, R.F.; Skaugset, A.E.; Weiler, M. Storage of water on vegetation under simulated rainfall of varying intensity. Adv. Water Resour. 2006, 29, 974–986. [Google Scholar] [CrossRef]
- Fyllas, N.M.; Dimitrakopoulos, P.G.; Troumbis, A.Y. Regeneration Dynamics of a Mixed Mediterranean Pine Forest in the Absence of Fire. For. Ecol. Manag. 2008, 256, 1552–1559. [Google Scholar] [CrossRef]
- Escudero, A.; Carnes, L.F.; Pérez-García, F. Seed Germination of Gypsophytes and Gypsovags in Semi-Arid Central Spain. J. Arid Environ. 1997, 36, 487–497. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Stand and Soil Attribute | Experimental Area | ||
|---|---|---|---|
| Palancares–Harvested | Majadas–Harvested | Sumidero– Unmanaged | |
| Density (trees ha−1) | 640 ± 41 | 656 ± 50 | 1199 ± 129 |
| Basal area (m2 ha−1) | 25 ± 5 | 25 ± 6 | 49 ± 6 |
| Total tree height (m) | 13 ± 6 | 11 ± 3 | 33 ± 4 |
| Shrub cover (%) | 53 ± 5 | 51 ± 7 | 41 ± 7 |
| pH | 6.37 ± 0.52 | 6.83 ± 0.52 | 9.86 ± 0.31 |
| Organic matter (%) | 7.48 ± 0.40 | 7.18 ± 0.40 | 4.11 ± 0.33 |
| NO3− (mg kg−1) | 27.72 ± 1.27 | 28.12 ± 1.27 | 32.29 ± 1.39 |
| Water-holding capacity (%) | 6.68 ± 0.16 | 6.14 ± 0.16 | 8.37 ± 0.38 |
| C/N | 10.44 ± 0.19 | 10.14 ± 0.19 | 8.58 ± 0.46 |
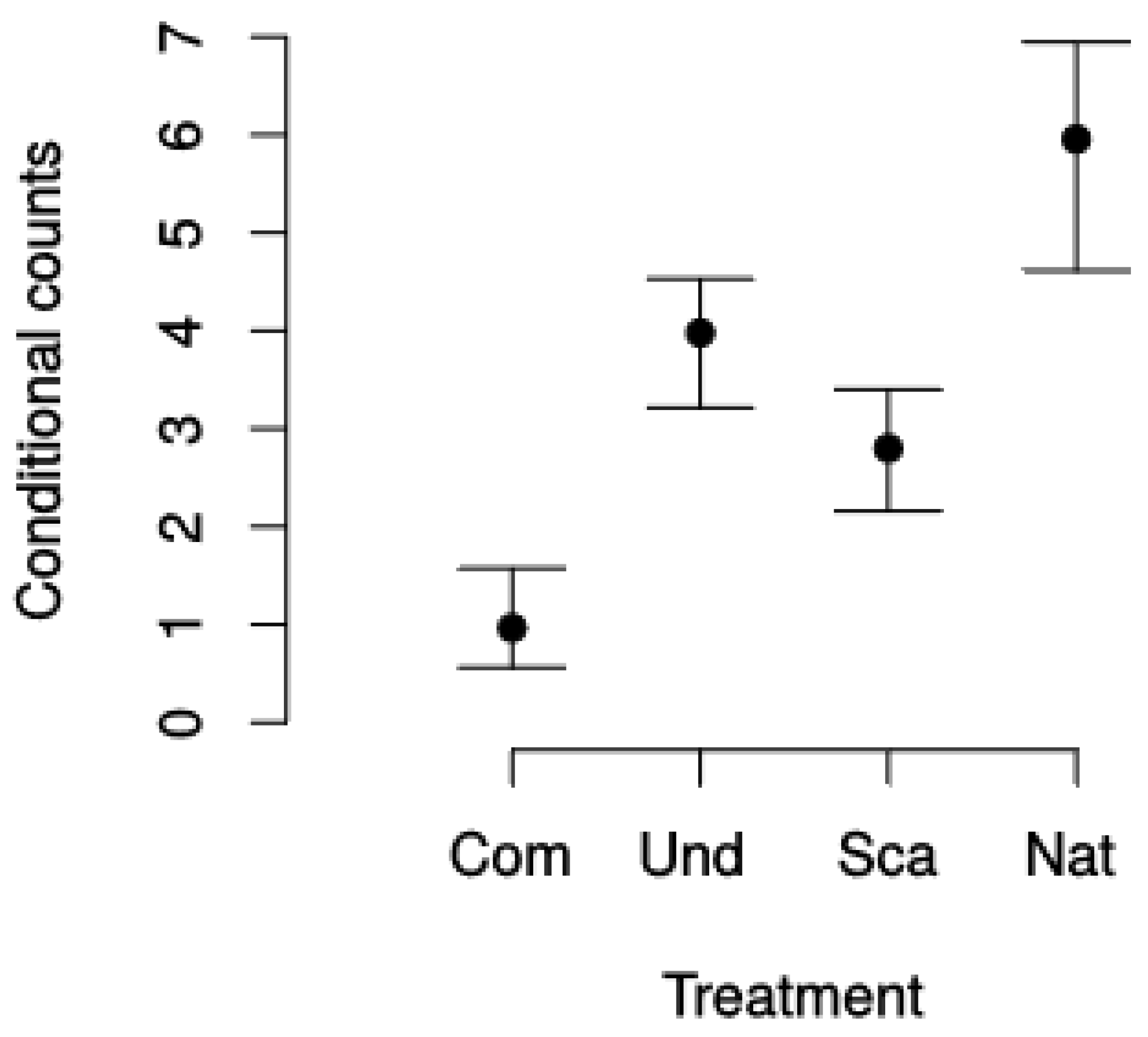
| Treatment | Site | Mean (Seedlings m2) | Std. Error |
|---|---|---|---|
| Compaction | Majadas | 0.2 | 0.5 |
| Compaction | Palancares | 1.7 | 2.3 |
| Scarification | Majadas | 2.2 | 1.9 |
| Scarification | Palancares | 3.4 | 2.3 |
| Undisturbed | Majadas | 3.4 | 2.8 |
| Undisturbed | Palancares | 4.6 | 4.5 |
| Natural | Sumidero | 6.0 | 10.2 |
| Treatment | Parameter | Estimate (Seedlings m2) | Std. Error | p-Value |
|---|---|---|---|---|
| Compaction (C, intercept) | β0,T | 0.3239 | 0.2395 | 0.1760 |
| Undisturbed (U) | β1,T | −3.8625 | 0.8508 | <0.0001 |
| Scarification (S) | β2,T | −2.4634 | 0.4848 | <0.0001 |
| Natural (N) | β3,T | −3.4377 | 0.7930 | <0.0001 |
| Treatment | Parameter | Estimate (Seedlings m2) | Std. Error | p-Value |
|---|---|---|---|---|
| Compaction (C, intercept) | γ0,T | 0.8342 | 0.1243 | <0.0001 |
| Undisturbed (U) | γ1,T | 0.5751 | 0.1358 | <0.0001 |
| Scarification (S) | γ2,T | 0.3067 | 0.1417 | 0.0304 |
| Natural (N) | γ3,T | 0.9936 | 0.1386 | <0.0001 |
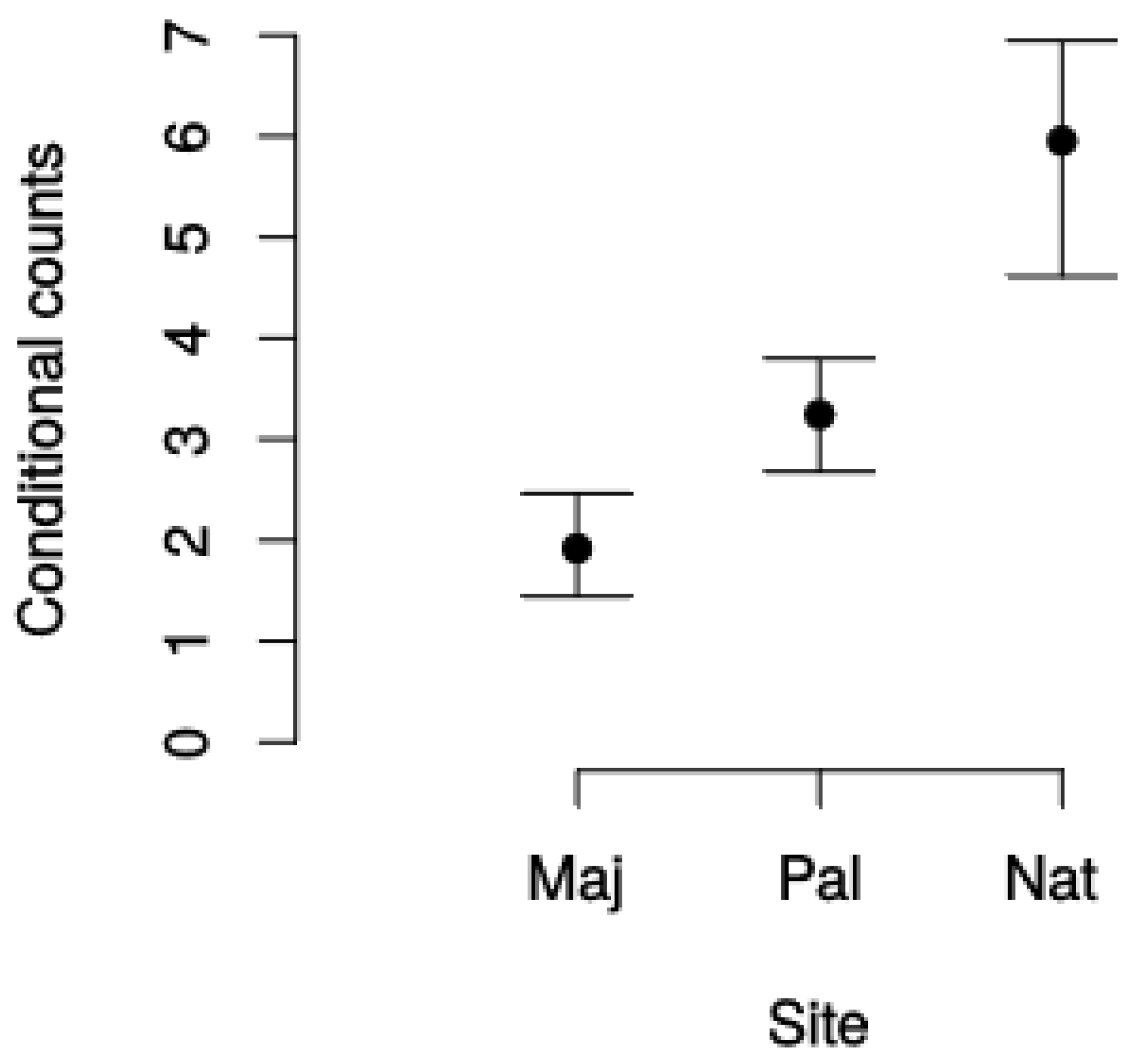
| Forest | Parameter | Estimate (Seedlings m2) | Std. Error | p-Value |
|---|---|---|---|---|
| Majadas–harvested (M, intercept) | β0,F | −0.6536 | 0.1988 | 0.0010 |
| Palancares–harvested (P) | β1,F | −1.1322 | 0.3323 | 0.0007 |
| Sumidero–unmanaged (S) | β2,F | −2.4634 | 0.7817 | 0.0016 |
| Forest | Parameter | Estimate (Seedlings m2) | Std. Error | p-Value |
|---|---|---|---|---|
| Majadas–harvested (M, intercept) | γ0,F | 1.07037 | 0.06808 | <0.0010 |
| Palancares–harvested (P) | γ1,F | 0.26157 | 0.08448 | 0.0020 |
| Sumidero–unmanaged (S) | γ2,F | 0.75743 | 0.09172 | <0.0001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucas-Borja, M.E.; Heydari, M.; Miralles, I.; Zema, D.A.; Manso, R. Effects of Skidding Operations after Tree Harvesting and Soil Scarification by Felled Trees on Initial Seedling Emergence of Spanish Black Pine (Pinus nigra Arn. ssp. salzmannii). Forests 2020, 11, 767. https://doi.org/10.3390/f11070767
Lucas-Borja ME, Heydari M, Miralles I, Zema DA, Manso R. Effects of Skidding Operations after Tree Harvesting and Soil Scarification by Felled Trees on Initial Seedling Emergence of Spanish Black Pine (Pinus nigra Arn. ssp. salzmannii). Forests. 2020; 11(7):767. https://doi.org/10.3390/f11070767
Chicago/Turabian StyleLucas-Borja, Manuel Esteban, Mehdi Heydari, Isabel Miralles, Demetrio Antonio Zema, and Ruben Manso. 2020. "Effects of Skidding Operations after Tree Harvesting and Soil Scarification by Felled Trees on Initial Seedling Emergence of Spanish Black Pine (Pinus nigra Arn. ssp. salzmannii)" Forests 11, no. 7: 767. https://doi.org/10.3390/f11070767
APA StyleLucas-Borja, M. E., Heydari, M., Miralles, I., Zema, D. A., & Manso, R. (2020). Effects of Skidding Operations after Tree Harvesting and Soil Scarification by Felled Trees on Initial Seedling Emergence of Spanish Black Pine (Pinus nigra Arn. ssp. salzmannii). Forests, 11(7), 767. https://doi.org/10.3390/f11070767