Modeling Current and Future Potential Geographical Distribution of Carpinus tientaiensis, a Critically Endangered Species from China


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Species Distribution Data
2.2. Acquisition and Processing of Climate and Altitude Data
2.3. MaxEnt Model Operation and Evaluation
2.4. Classification of Habitat Suitability
2.5. Dynamic Changes and Centroid Migrations of the Suitable Distribution Area
3. Results
3.1. Model Accuracy
3.2. Suitable Distribution Areas of C. tientaiensis in the Current Climate
3.3. Dominant Environmental Factors Limiting the Survival and Distribution of C. tientaiensis
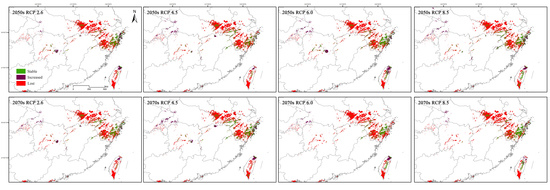
3.4. Suitable Distribution Areas of C. tientaiensis in Future Climate Change Scenarios
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Manthey, M.; Box, E.O. Realized climatic niches of deciduous trees: Comparing western Eurasia and eastern North America. J. Biogeogr. 2007, 34, 1028–1040. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change. Climate Change 2013: The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [CrossRef]
- Barrett, M.A.; Brown, J.L.; Junge, R.E.; Yoder, A.D. Climate change, predictive modeling and lemur health: Assessing impacts of changing climate on health and conservation in madagascar. Biol. Conserv. 2013, 157, 409–422. [Google Scholar] [CrossRef]
- Bertrand, R.; Lenoir, J.; Piedallu, C.; Riofrio-Dillon, G.; Ruffray, P.; Vidal, C.; Pierrat, J.; Gégout, J. Changes in plant community composition lag behind warming in lowland forests. Nature 2011, 479, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Dyderski, M.K.; Paź-Dyderska, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.C. Climate change. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef] [Green Version]
- Heinrichs, J.A.; Bender, D.J.; Schumaker, N.H. Habitat degradation and loss as key drivers of regional population extinction. Ecol. Model. 2016, 335, 64–73. [Google Scholar] [CrossRef]
- Bogich, T.L.; Barker, G.M.; Mahlfeld, K.; Climo, F.; Green, R.; Balmford, A. Fragmentation, grazing and the species-area relationship. Ecography 2012, 35, 224–231. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M. Ecological Niches and Geographic Distributions; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Qin, A.L.; Liu, B.; Guo, Q.S.; Bussmann, R.W.; Ma, F.Q.; Jian, Z.J.; Xu, G.X.; Pei, S.X. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Newbold, T. Applications and limitations of museum data for conservation and ecology, with particular attention to species distribution models. Prog. Phys. Geogr. 2010, 34, 3–22. [Google Scholar] [CrossRef]
- Marcer, A.; Sáez, L.; Molowny-Horas, R.; Pons, X.; Pino, J. Using species distribution modelling to disentangle realised versus potential distributions for rare species conservation. Biol. Conserv. 2013, 166, 221–230. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distribution. Ecol. Model. 2006, 19, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Li, G.Q.; Du, S.; Wen, Z.M. Mapping the climatic suitable habitat of oriental arborvitae (Platycladus orientalis) for introduction and cultivation at a global scale. Sci. Rep. 2016, 6, 30009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borthakur, S.K.; Baruah, P.S.; Deka, K.; Das, P.; Sarma, B.; Adhikari, D.; Tanti, B. Habitat distribution modelling for improving conservation status of Brucea mollis Wall. ex Kurz.–An endangered potential medicinal plant of Northeast India. J. Nat. Conserv. 2018, 43, 104–110. [Google Scholar] [CrossRef]
- Singh, P.B.; Mainali, K.; Jiang, Z.; Thapa, A.; Subedi, N.; Awan, M.N.; Ilyas, O.; Luitel, H.; Zhou, Z.; Hu, H. Projected distribution and climate refugia of endangered Kashmir musk deer Moschus cupreus in greater Himalaya, South Asia. Sci. Rep. 2020, 10, 1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.C. Plantae novae Chekiangenses. In Contributions from the Biological Laboratory of the Science Society of China; Botanical Series; The Biological laboratory of the Science Society of China: Nanjing, China, 1932; Volume 8, pp. 135–142. [Google Scholar]
- Li, P.C.; Skvortsov, A.K. Betulaceae. In Flora of China; Wu, C.Y., Raven, P.H., Eds.; Science Press: Beijing, China, 1999; Volume 4, pp. 289–300. [Google Scholar]
- Shaw, K.; Roy, S.; Wilson, B. Carpinus tientaiensis. The IUCN Red List of Threatened Species 2014, e.T194617A2353175. Available online: https://www.iucnredlist.org/species/194617/2353175 (accessed on 1 August 2014). [CrossRef]
- Chen, M.S.; Jin, Z.X.; Ke, S.X. Measurement and analysis of leaf shape variation of Carpinus tientaiensis in different light environment. Sci. Silvae Sin. 2018, 54, 54–63. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Taylor, K.E.; Ronald, S.; Meehl, G.A. An overview of CMIP5 and the Experiment Design. Bull. Am. Meteorol. Soc. 2011, 93, 485–498. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J. Transferability, sample selection bias and background data in presence-only modelling: A response to Peterson et al. (2007). Ecography 2008, 31, 272–278. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, D.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2010, 17, 43–57. [Google Scholar] [CrossRef]
- Raes, N.; Steege, H.T. A null-model for significance testing of presence-only species distribution models. Ecography 2007, 30, 727–736. [Google Scholar] [CrossRef]
- Fielding, A.H. What are the appropriate characteristics of an accuracy measure? In Predicting Species Occurrences: Issues of Accuracy and Scale; Scott, J.M., Heglund, P., Morrison, M.L., Raven, P.H., Eds.; Island Press: Washington, DC, USA, 2002; pp. 271–280. [Google Scholar]
- Reineking, B.; Der, B.S. Constrain to perform: Regularization of habitat models. Ecol. Model. 2006, 193, 675–690. [Google Scholar] [CrossRef]
- Khadka, K.K.; Kannan, R.; Ilyas, O.; Abbas, F.; James, D.A. Where are they? Where will they be? In pursuit of current and future whereabouts of endangered Himalayan musk deer. Mamm. Biol. 2017, 85, 30–36. [Google Scholar] [CrossRef]
- Zhang, K.L.; Yao, L.J.; Meng, J.S.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- Narouei-Khandan, H.A. Ensemble Models to Assess the Risk of Exotic Plant Pathogens in a Changing Climate. Master’s Thesis, Lincoln University, Oakland, CA, USA, 2014. [Google Scholar]
- Robertson, M.P.; Peter, C.I.; Villet, M.H.; Ripley, B.S. Comparing models for predicting species’ potential distributions: A case study using correlative and mechanistic predictive modelling techniques. Ecol. Model. 2003, 164, 153–167. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.N.; Seo, C.; Thorne, J.; Nelson, J.K.; Erwin, S.; O’Brien, J.M.; Schwartz, M.W. Using species distribution models to predict new occurrences for rare plants. Divers. Distrib. 2009, 15, 565–576. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Guisan, A.; Broennimann, O.; Engler, R.; Vust, M.; Yoccoz, N.G.; Lehmann, A.; Zimmermann, N.E. Using Niche-based models to improve the sampling of rare species. Conserv. Biol. 2006, 20, 501–511. [Google Scholar] [CrossRef]
- Garcia, K.; Lasco, R.; Ines, A.; Lyon, B.; Pulhin, F. Predicting geographic distribution and habitat suitability due to climate change of selected threatened forest tree species in the Philippines. Appl. Geogr. 2013, 44, 12–22. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudik, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Hernández, P.A.; Graham, C.H.; Master, M.M.; Albert, D. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M.; Matthews, S.N.; Peters, M.P. Estimating potential habitat for 134 eastern US tree species under six climate scenarios. Forest Ecol. Manag. 2008, 254, 390–406. [Google Scholar] [CrossRef]
- Shaw, K.; Roy, S.; Wilson, B. Carpinus putoensis. The IUCN Red List of Threatened Species 2014, e.T32303A2813038. Available online: https://www.iucnredlist.org/species/32303/2813038 (accessed on 1 August 2014). [CrossRef]
- Roy, S.; Shaw, K.; Wilson, B. Carpinus hebestroma. The IUCN Red List of Threatened Species 2016, e.T194577A2350438. Available online: https://www.iucnredlist.org/species/194577/2350438 (accessed on 1 August 2014). [CrossRef]
- Lu, Z.Q.; Liu, J.Q.; Beech, E. Carpinus langaoensis. The IUCN Red List of Threatened Species 2018. Available online: https://www.iucnredlist.org/species/117839549/117839554 (accessed on 8 August 2017). [CrossRef]
- Engler, R.; Guisan, A.; Rechsteiner, L. An improved approach for predicting the distribution of rare and endangered species from occurrence and pseudo-absence data. J. Appl. Ecol. 2004, 41, 263–274. [Google Scholar] [CrossRef]
- Cai, R.S.; Tan, H.J.; Kontoyiannis, H. Robust surface warming in offshore china seas and its relationship to the East Asian monsoon wind field and ocean forcing on interdecadal time scales. J. Clim. 2017, 30, 8987–9005. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Li, S.S.; Lu, J.Y.; Yan, J.P.; Liu, X.; Kong, F.; Wang, J. Spatiotemporal variability of temperature in northern and southern Qinling Mountains and its influence on climatic boundary. Acta Geogr. Sin. 2018, 73, 13–24. [Google Scholar] [CrossRef]
- Burbano-Girón, J. Modeling the Dispersion of Atelines (Primates, Atelinae) Through Scenarios of Climate Change and Habitat Fragmentation in Colombia. Conservation Implications for the Persistence of Species into the Future. Master’s Thesis, Universidad Nacional de Colombia, Bogotá, Colombia, 2013. [Google Scholar]
- Hu, X.S.; Wu, C.Z.; Hong, W.; Qiu, R.Z.; Li, J.; Hong, T. Forest cover change and its drivers in the upstream area of the Minjiang River, China. Ecol. Indic. 2014, 46, 121–128. [Google Scholar] [CrossRef]
- Yi, Y.J.; Zhou, Y.; Cai, Y.P.; Yang, W.; Li, Z.W.; Zhao, X. The influence of climate change on an endangered riparian plant species: The root of riparian Homonoia. Ecol. Indic. 2018, 92, 40–50. [Google Scholar] [CrossRef]
- Yi, Y.J.; Tang, C.H.; Yang, Z.F.; Chen, X. Influence of Manwan Reservoir on fish habitat in the middle reach of the Lancang River. Ecol. Eng. 2014, 69, 106–117. [Google Scholar] [CrossRef]
- Cunha, D.D.A.; Ferreira, L.V. Impacts of the Belo Monte hydroelectric dam construction on pioneer vegetation formations along the Xingu River, Pará State, Brazil. Braz. J. Bot. 2012, 35, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.Y.; Zhang, L.F.; Shen, P.; Ren, B.Q.; Liang, Y.; Chen, Z.D. Wind Pollination Characteristics of Styles in Betulaceae. Chin. Bull. Bot. 2014, 49, 524–538. [Google Scholar] [CrossRef]
- Vetaas, O.R. Realized and potential climate niches: A comparison of four Rhododendron tree species. J. Biogeogr. 2002, 29, 545–554. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Variable Description | Unit | Contribution Rate |
|---|---|---|---|
| bio17 | Precipitation of the driest quarter | mm | 17.7 |
| bio15 | CV of precipitation seasonality | % | 16.6 |
| bio12 | Annual precipitation | mm | 15.3 |
| bio14 | Precipitation of the driest month | mm | 13.0 |
| bio18 | Precipitation of the coldest quarter | mm | 10.7 |
| bio3 | Isothermality (bio2/bio7 × 100) | % | 9.7 |
| bio2 | Mean diurnal temperature range | °C | 9.5 |
| bio10 | Mean temperature of warmest quarter | °C | 3.5 |
| bio1 | Annual mean temperature | °C | 1.5 |
| bio19 | Precipitation of the coldest quarter | mm | 1.0 |
| bio8 | Mean temperature of wettest quarter | °C | 0.8 |
| alt | Altitude | m | 0.3 |
| bio16 | Precipitation of wettest quarter | mm | 0.2 |
| bio9 | Mean temperature of driest quarter | °C | 0.1 |
| bio6 | Min temperature of coldest month | °C | 0.0 |
| Climate Scenarios | Suitable Distribution Area (103 km2) | Core Distribution Area (103 km2) | Stable | Increased | Lost | ||||
|---|---|---|---|---|---|---|---|---|---|
| Area (103 km2) | Rate (%) | Area (103 km2) | Rate (%) | Area (103 km2) | Rate (%) | ||||
| Current | 90.79 | 13.68 | - | - | - | - | - | - | |
| 2050s | RCP 2.6 | 26.20 | 2.99 | 17.74 | 19.54 | 8.46 | 9.32 | 73.05 | 80.46 |
| RCP 4.5 | 24.12 | 2.03 | 16.15 | 17.79 | 7.97 | 8.78 | 74.64 | 82.21 | |
| RCP 6.0 | 32.03 | 2.20 | 20.21 | 22.26 | 11.82 | 13.02 | 70.58 | 77.74 | |
| RCP 8.5 | 33.05 | 3.66 | 20.64 | 22.73 | 12.41 | 13.67 | 70.05 | 77.27 | |
| Mean | 28.85 | 2.72 | 18.68 | 20.58 | 10.17 | 11.20 | 72.11 | 79.42 | |
| 2070s | RCP 2.6 | 25.01 | 2.39 | 17.56 | 19.34 | 7.45 | 8.21 | 73.23 | 80.66 |
| RCP 4.5 | 31.03 | 2.54 | 19.42 | 21.39 | 11.61 | 12.79 | 71.37 | 78.61 | |
| RCP 6.0 | 19.41 | 2.21 | 13.62 | 15.00 | 5.80 | 6.39 | 77.17 | 85.00 | |
| RCP 8.5 | 23.36 | 2.34 | 18.37 | 20.24 | 4.99 | 5.50 | 72.42 | 79.76 | |
| Mean | 24.70 | 2.37 | 17.24 | 18.99 | 7.46 | 8.22 | 73.55 | 81.01 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, R.; Chu, X.; He, Q.; Tang, Y.; Song, M.; Zhu, Z. Modeling Current and Future Potential Geographical Distribution of Carpinus tientaiensis, a Critically Endangered Species from China. Forests 2020, 11, 774. https://doi.org/10.3390/f11070774
Zhao R, Chu X, He Q, Tang Y, Song M, Zhu Z. Modeling Current and Future Potential Geographical Distribution of Carpinus tientaiensis, a Critically Endangered Species from China. Forests. 2020; 11(7):774. https://doi.org/10.3390/f11070774
Chicago/Turabian StyleZhao, Runan, Xiaojie Chu, Qianqian He, Yan Tang, Min Song, and Zunling Zhu. 2020. "Modeling Current and Future Potential Geographical Distribution of Carpinus tientaiensis, a Critically Endangered Species from China" Forests 11, no. 7: 774. https://doi.org/10.3390/f11070774
APA StyleZhao, R., Chu, X., He, Q., Tang, Y., Song, M., & Zhu, Z. (2020). Modeling Current and Future Potential Geographical Distribution of Carpinus tientaiensis, a Critically Endangered Species from China. Forests, 11(7), 774. https://doi.org/10.3390/f11070774