Potential Pollution Sources from Agricultural Activities on Tropical Forested Floodplain Wetlands Revealed by Soil eDNA


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Soil Isotopes and Plant End-Members
2.3. eDNA Analyses
2.4. Data Analyses
3. Results
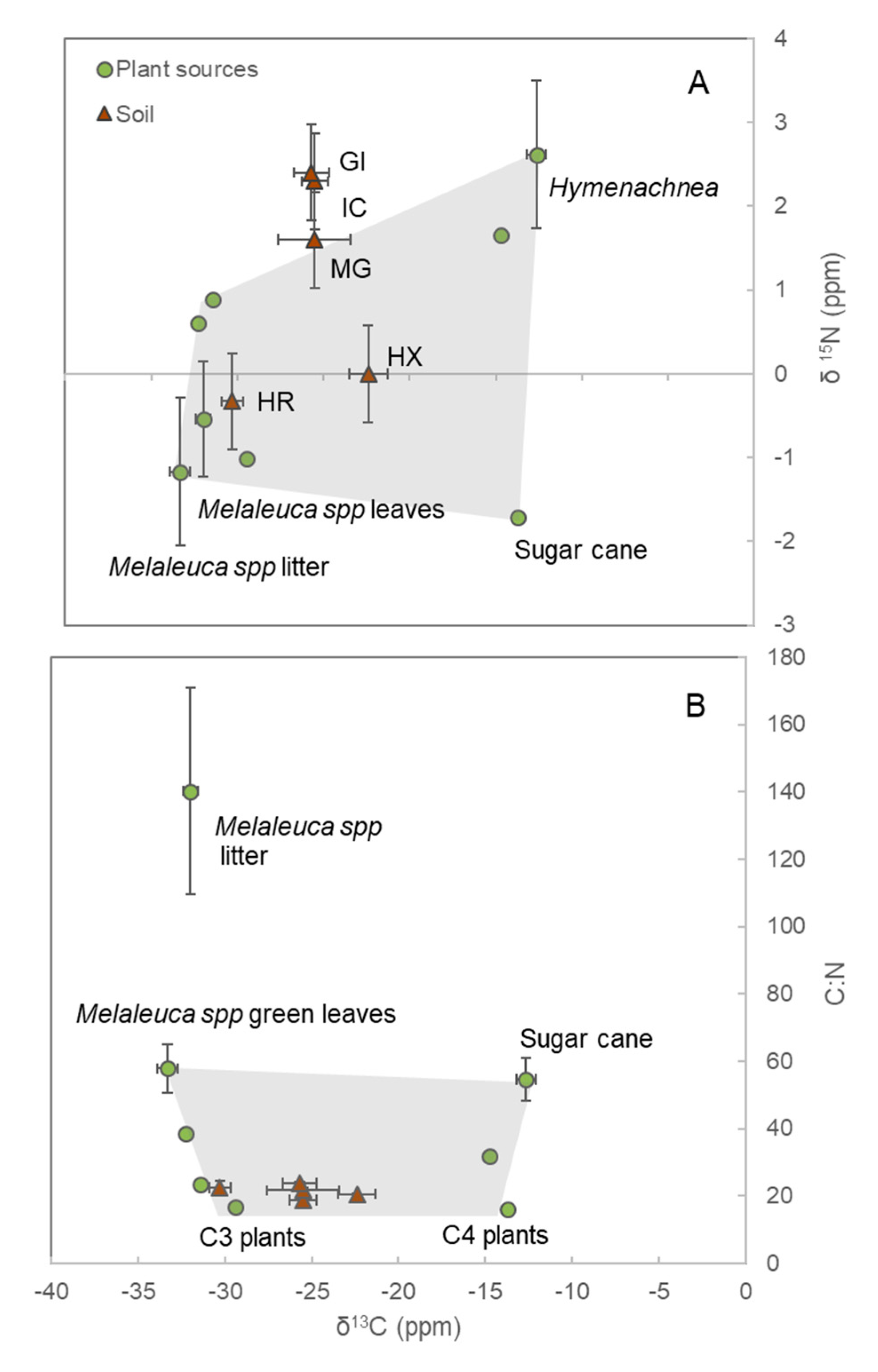
3.1. Soil Isotopes and End-Members
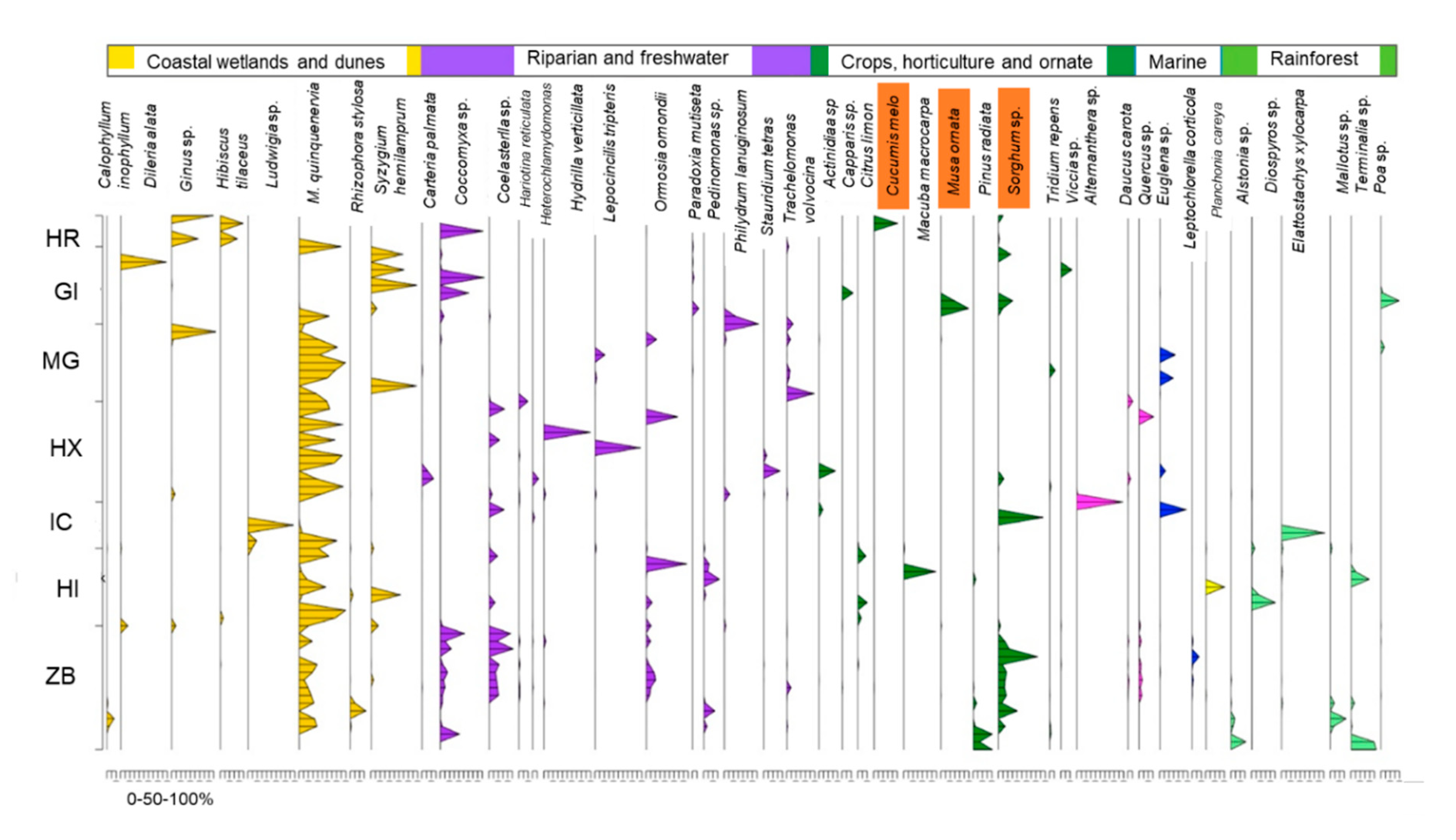
3.2. eDNA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Junk, W.F.; Bayley, P.B.; Sparks, R.E. The flood pulse concept in river-floodplain systems. Can. J. Fish. Aquat. Sci. 1989, 106, 110–127. [Google Scholar]
- Adame, M.; Pettit, N.; Valdez, D.; Ward, D.; Burford, M.; Bunn, S. The contribution of epiphyton to the primary production of tropical floodplain wetlands. Biotropica 2017, 49, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Haugaasen, T.; Peres, C.A.; Journal, S.; Mar, N.; Haugaasen, T.; Peres, C.A. Mammal assemblage structure in Amazonian flooded and unflooded forests. J. Trop. Ecol. 2016, 21, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.; Pearson, R.; Brodie, J.; Butler, B. Review and conceptual models of agricultural impacts and water quality in waterways of the Great Barrier Reef catchment area. Mar. Freshw. Res. 2016, 68, 1–19. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J. Wetlands, 5th ed.; Wiley: Hoboken, NJ, USA, 2015. [Google Scholar]
- Adame, M.F.; Reef, R.; Wong, V.N.L.; Balcombe, S.R.; Turschwell, M.P.; Kavehei, E.; Rodríguez, D.C.; Kelleway, J.J.; Masque, P.; Ronan, M. Carbon and nitrogen sequestration of Melaleuca floodplain wetlands in tropical Australia. Ecosystems 2019, 23, 454–466. [Google Scholar] [CrossRef]
- Lougheed, V.L.; Mcintosh, M.D.; Parker, C.A.; Stevenson, R.J. Wetland degradation leads to homogenization of the biota at local and landscape scales. Freshw. Biol. 2008, 53, 2402–2413. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gossilink, J.G. The value of wetlands: Importance of scale and landscape setting. Ecol. Econ. 2000, 35, 25–33. [Google Scholar] [CrossRef]
- Vymazal, J.; Březinová, T. The use of constructed wetlands for removal of pesticides from agricultural runoff and drainage: A review. Environ. Int. 2015, 75, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Bruesewitz, D.A.; Hamilton, D.P.; Schipper, L.A. Denitrification potential in lake sediment increases across a gradient of catchment agriculture. Ecosystems 2011, 14, 341–352. [Google Scholar] [CrossRef]
- Houlahan, J.E.; Findlay, C.S. Estimating the “critical” distance at which adjacent land-use degrades wetland water and sediment quality. Landsc. Ecol. 2004, 19, 677–690. [Google Scholar] [CrossRef]
- Johnson, A.K.L.; Ebert, S.P. Quantifying inputs of pesticides to the Great Barrier Reef Marine Park—A case study in the Herbert River catchment of North-East Queensland. Mar. Pollut. Bull. 2000, 41, 302–309. [Google Scholar] [CrossRef]
- Bainbridge, Z.T.; Brodie, J.E.; Faithful, J.W.; Sydes, D.A.; Lewis, S.E. Identifying the land-based sources of suspended sediments, nutrients and pesticides discharged to the Great Barrier Reef from the Tully—Murray Basin, Queensland, Australia. Mar. Freshw. Res. 2009, 60, 1081–1090. [Google Scholar] [CrossRef]
- Kroon, F.J.; Hook, S.E.; Jones, D.; Metcalfe, S.; Henderson, B.; Smith, R.; Warne, M.S.J.; Turner, R.D.; McKeown, A.; Westcott, D.A. Altered transcription levels of endocrine associated genes in two fisheries species collected from the Great Barrier Reef catchment and lagoon. Mar. Environ. Res. 2015, 104, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adame, M.F.; Arthington, A.H.; Waltham, N.; Hasan, S.; Selles, A.; Ronan, M. Managing threats and restoring wetlands within catchments of the Great Barrier Reef, Australia. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 829–839. [Google Scholar] [CrossRef]
- Vandergragt, M.; Warne, M.S.; Borschmann, G.; Johns, C.V. Pervasive pesticide contamination of wetlands in the Great Barrier Reef catchment area. J. Integr. Environ. Assess. Manag. 2020. [Google Scholar] [CrossRef]
- Brodie, J.E.; Mitchell, A.W. Nutrients in Australian tropical rivers: Changes with agricultural development and implications for receiving environments. Mar. Freshw. Res. 2005, 56, 279–302. [Google Scholar] [CrossRef]
- Costanzo, S.D.; O’Donohue, M.J.; Dennison, W.C.; Loneragan, N.R.; Thomas, M. A new approach for detecting and mapping sewage impacts. Mar. Pollut. Bull. 2001, 149–156. [Google Scholar] [CrossRef]
- Bahadori, M.; Chen, C.; Lewis, S.; Rezaei Rashti, M.; Cook, F.; Parnell, A.; Esfandbod, M.; Boyd, S. A novel approach of combining isotopic and geochemical signatures to differentiate the sources of sediments and particulate nutrients from different land uses. Sci. Total Environ. 2019, 655, 129–140. [Google Scholar] [CrossRef]
- Adame, M.F.; Wright, A.; Grinham, A.; Lobb, K.; Raymond, C.E.; Lovelock, C.E. Terrestrial-marine connectivity: Patterns of terrestrial soil carbon deposition in coastal sediments determined by analysis of glomalin related soil protein. Limnol. Oceanogr. 2012, 57, 1492–1502. [Google Scholar] [CrossRef] [Green Version]
- López-Merino, L.; Colás-Ruiz, N.R.; Adame, M.F.; Serrano, O.; Martínez Cortizas, A.; Mateo, M.A. A six thousand-year record of climate and land-use change from Mediterranean seagrass mats. J. Ecol. 2017, 105, 1267–1278. [Google Scholar] [CrossRef]
- Reef, R.; Atwood, T.B.; Samper-Villarreal, J.; Adame, M.F.; Sampayo, E.M.; Lovelock, C.E. Using eDNA to determine the source of organic carbon in seagrass meadows. Limnol. Oceanogr. 2017, 62, 1254–1265. [Google Scholar] [CrossRef]
- Australian Bureau of Meteorology. 2020. Available online: http://www.bom.gov.au/ (accessed on 10 May 2020).
- Queensland Governement. WetlandInfo. 2020. Available online: https://wetlandinfo.des.qld.gov.au/ (accessed on 10 May 2020).
- Karim, F.; Kinsey-Henderson, A.; Wallace, J.; Godfrey, P.; Arthington, A.H.; Pearson, R.G. Modelling hydrological connectivity of tropical floodplain wetlands via a combined natural and artificial stream network. Hydrol. Process. 2013, 28, 5696–5710. [Google Scholar] [CrossRef]
- Rasiah, V.; Armour, J.D.; Cogle, A.L.; Florentine, S.K. Nitrate import—Export dynamics in groundwater interacting with surface-water in a wet-tropical environment. Soil Res. 2010, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Queensland Government. QSpatial Catalogue. Australia. 2020. Available online: http://qldspatial.information.qld.gov.au/catalogue/custom/index.page (accessed on 20 July 2020).
- Finlayson, C.M. Plant ecology of Australia’s tropical floodplain wetlands: A review. Ann. Bot. 2005, 96, 541–555. [Google Scholar] [CrossRef] [Green Version]
- Turner, C.E.; Center, T.D.; Burrows, D.W.; Buckingham, G.R. Ecology and management of Melaleuca quinqenervia, an invader of wetlands in Florida, U.S.A. Wetl. Ecol. Manag. 1998, 5, 165–178. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T. Stamatakis PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 1, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.; Haas, B.; Clemente, J.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 15, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.; Kuczynski, J.; Stomabaugh, J.; Bittinger, K.; Bushman, F.; Costello, E.; Fierer, N.; Peña, A.; Goodrich, J.; Gordon, J.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Fry, B. Alternative approaches for solving underdetermined isotope mixing problems. Mar. Ecol. Prog. Ser. 2013, 472, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Adame, M.; Jardine, T.; Fry, B.; Valdez, D.; Linder, G.; Nadji, J.; Bunn, S. Estuarine crocodiles in a tropical coastal floodplain obtain nutrition from terrestrial prey. PLoS ONE 2018, e0200983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, D.; Gregg, J. Uncertainty in source partitioning using stable isotopes. Oecologia 2011, 127, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Burrage, D.M.; Heron, M.L.; Hacker, J.M.; Stieglitz, T.C.; Steinberg, C.R.; Prytz, A. Evolution and dynamics of tropical river plumes in the Great Barrier Reef: An integrated remote sensing and in situ study. J. Geophys. Res. 2002, 107, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lovelock, C.E.; Duarte, C.M. Dimensions of blue carbon and emerging perspectives. Biol. Lett. 2019, 15, 20180781. [Google Scholar] [CrossRef] [PubMed]
- Saintilan, N.; Rogers, K.; Mazumder, D.; Woodroffe, C. Allochthonous and autochthonous contributions to carbon accumulation and carbon store in southeastern Australian coast. Estuar. Coast. Shelf Sci. 2013, 128, 84–92. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Site | Catchment | Sub-Catchment | Nearby Land Use and Impacts | Vegetation | Hydrology |
|---|---|---|---|---|---|
| (HR) Hull River National Park | Tully-Murray River | Hull River | Grazing Sugarcane Damaged during cyclone in 2011 | M. viridiflora Rainforest | Permanently flooded |
| (GI) Girramay National Park | Whitfield-Dallachy Creek | Grazing Protected rainforest | M. quinquenerva Rainforest | Flooded by rain and overbank flows | |
| (MG) Mungalla | Herbert River | Palm Creek | Grazing Treatment plant discharge | M. quinquenerva | Flooded by nearby lagoon |
| (HX) Halifax Wetlands National Park | Trebonne Creek | Grazing Sugar cane Protected wetlands | M. quinquenerva Mangroves Marsh | Flooded by rain and tidal flows | |
| (IC) Insulator Creek | Insulator Creek | Sugar cane Protected rainforest | M. quinquenerva Mangroves Marsh | Flooded by rain and overbank flows | |
| (HI) East Hinchinbrook Island | Hinchinbrook Island | East Hinchinbrook | Protected rainforest and beach | M. quinquenerva Rainforest | Flooded by rain and groundwater |
| (ZB) Zoe Bay | Southeast Hinchinbrook | Protected wetlands and rainforests | M. quinquenerva Mangroves Marsh | Flooded by fringing tidal creek |
| Sites | BD (g cm−3) | N (%) | C (%) | C:N | δ13C (‰) | δ 15N (‰) |
|---|---|---|---|---|---|---|
| HR | 0.34 ± 0.05 | 0.78 ± 0.04 | 14.3 ± 1.4 | 22.4 ± 2.2 | −30.3 ± 0.6 | −0.3 ± 0.6 |
| GI | 0.59 ± 0.02 | 0.34 ± 0.02 | 4.1 ± 0.4 | 23.8 ± 0.4 | −25.7 ± 1.0 | 2.4 ± 0.4 |
| MG | 0.61 ± 0.07 | 0.44 ± 0.15 | 7.6 ± 2.8 | 21.7 ± 0.7 | −25.5 ± 2.1 | 1.6 ± 0.9 |
| HX | 0.84 ± 0.12 | 0.06 ± 0.01 | 1.0 ± 0.2 | 20.6 ± 0.3 | −22.4 ± 1.1 | n.d. |
| IC | 0.65 ± 0.10 | 0.54 ± 0.14 | 7.8 ± 2.2 | 18.7 ± 1.2 | −25.5 ± 0.8 | 2.3 ± 0.4 |
| Site | Native Trees | Grass/Shrubs | Macrophyte | Microalgae | Non-Native | Cultivated |
|---|---|---|---|---|---|---|
| HR | 21.5 | 0.03 | 0.01 | 0.04 | 0.01 | 16.0 |
| GI | 50.7 | 4.9 | 0.1 | 20.0 | 0.1 | 20.7 |
| MG | 57.8 | 0.8 | 17.2 | 18.9 | 0.9 | 1.4 |
| HX | 57.1 | 0.2 | 9.6 | 22.1 | n.d. | 6.8 |
| IC | 30.6 | 35.7 | 0.02 | 15.1 | 0.01 | 17.4 |
| HI | 69.6 | 1.9 | 0.03 | 4.0 | 0.3 | 7.5 |
| ZB | 35.8 | 0.1 | 0.6 | 16.7 | 0.5 | 24.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adame, M.F.; Reef, R. Potential Pollution Sources from Agricultural Activities on Tropical Forested Floodplain Wetlands Revealed by Soil eDNA. Forests 2020, 11, 892. https://doi.org/10.3390/f11080892
Adame MF, Reef R. Potential Pollution Sources from Agricultural Activities on Tropical Forested Floodplain Wetlands Revealed by Soil eDNA. Forests. 2020; 11(8):892. https://doi.org/10.3390/f11080892
Chicago/Turabian StyleAdame, Maria Fernanda, and Ruth Reef. 2020. "Potential Pollution Sources from Agricultural Activities on Tropical Forested Floodplain Wetlands Revealed by Soil eDNA" Forests 11, no. 8: 892. https://doi.org/10.3390/f11080892
APA StyleAdame, M. F., & Reef, R. (2020). Potential Pollution Sources from Agricultural Activities on Tropical Forested Floodplain Wetlands Revealed by Soil eDNA. Forests, 11(8), 892. https://doi.org/10.3390/f11080892