A Numerical Approach to Estimate Natural Frequency of Trees with Variable Properties


Abstract
:1. Introduction
2. Methods
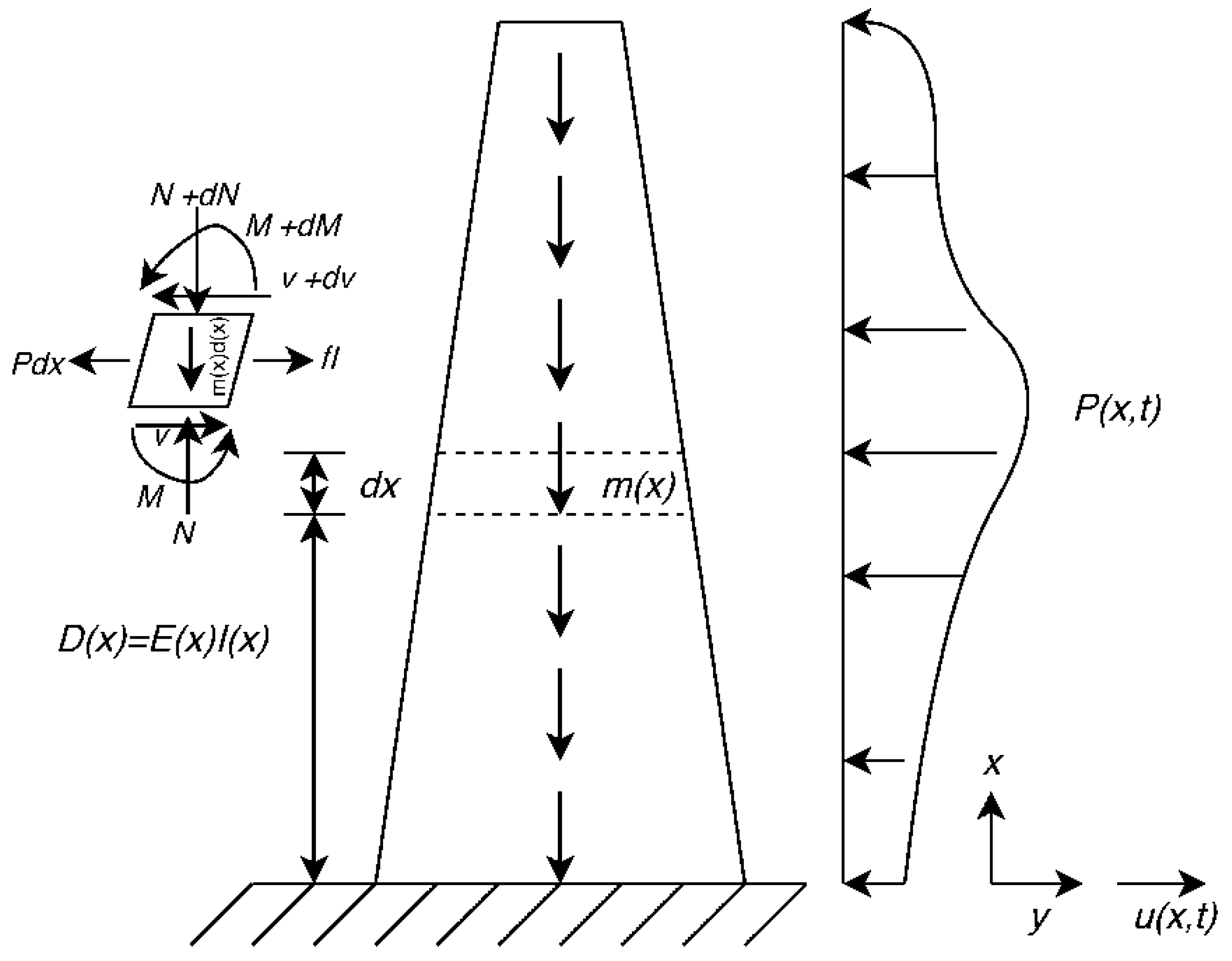
2.1. Theoretical Basis for Vibration Analysis
2.2. Calculation of Natural Frequency Using Numerical Analysis
2.2.1. Effect of Base Fixity
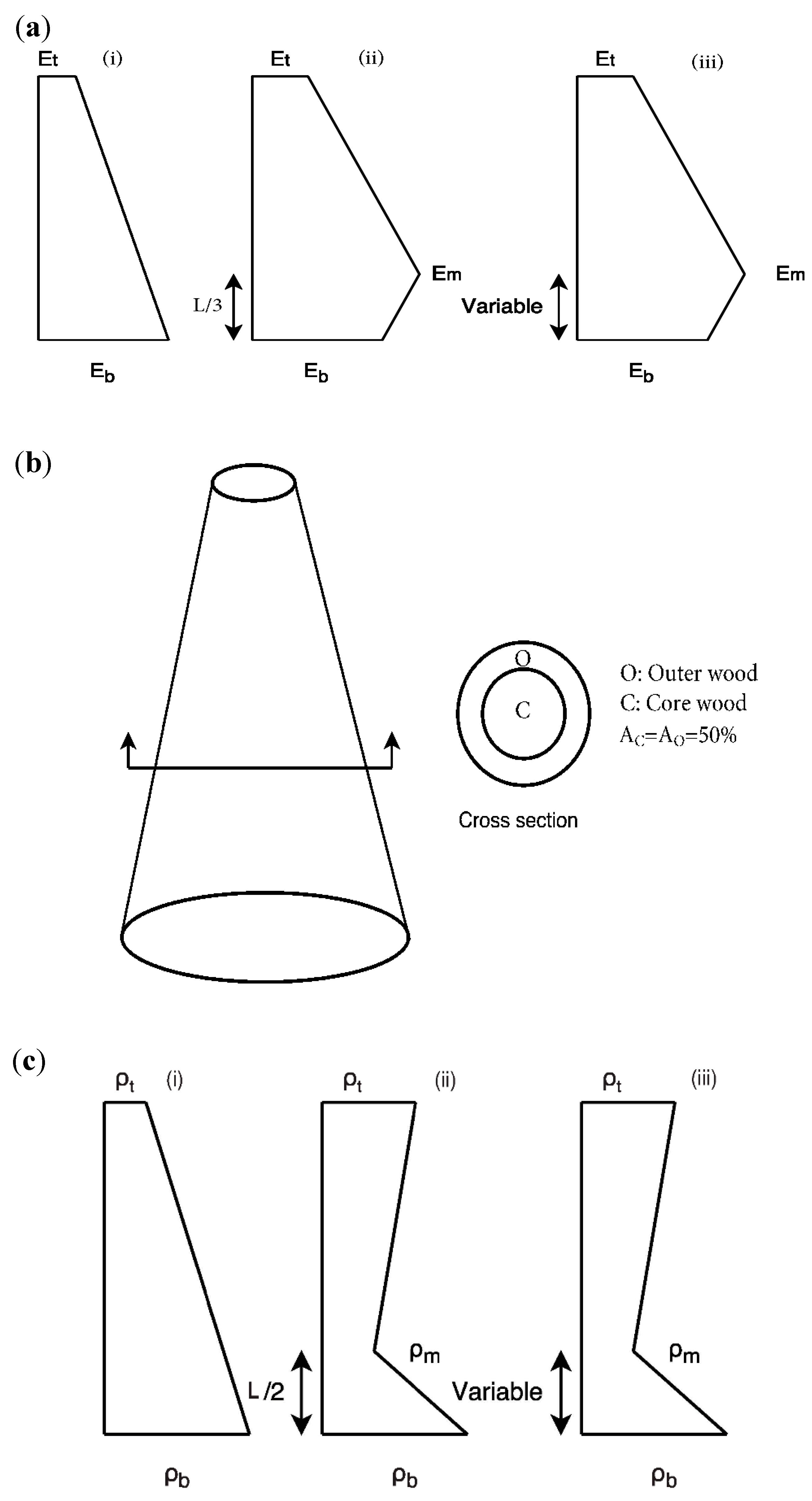
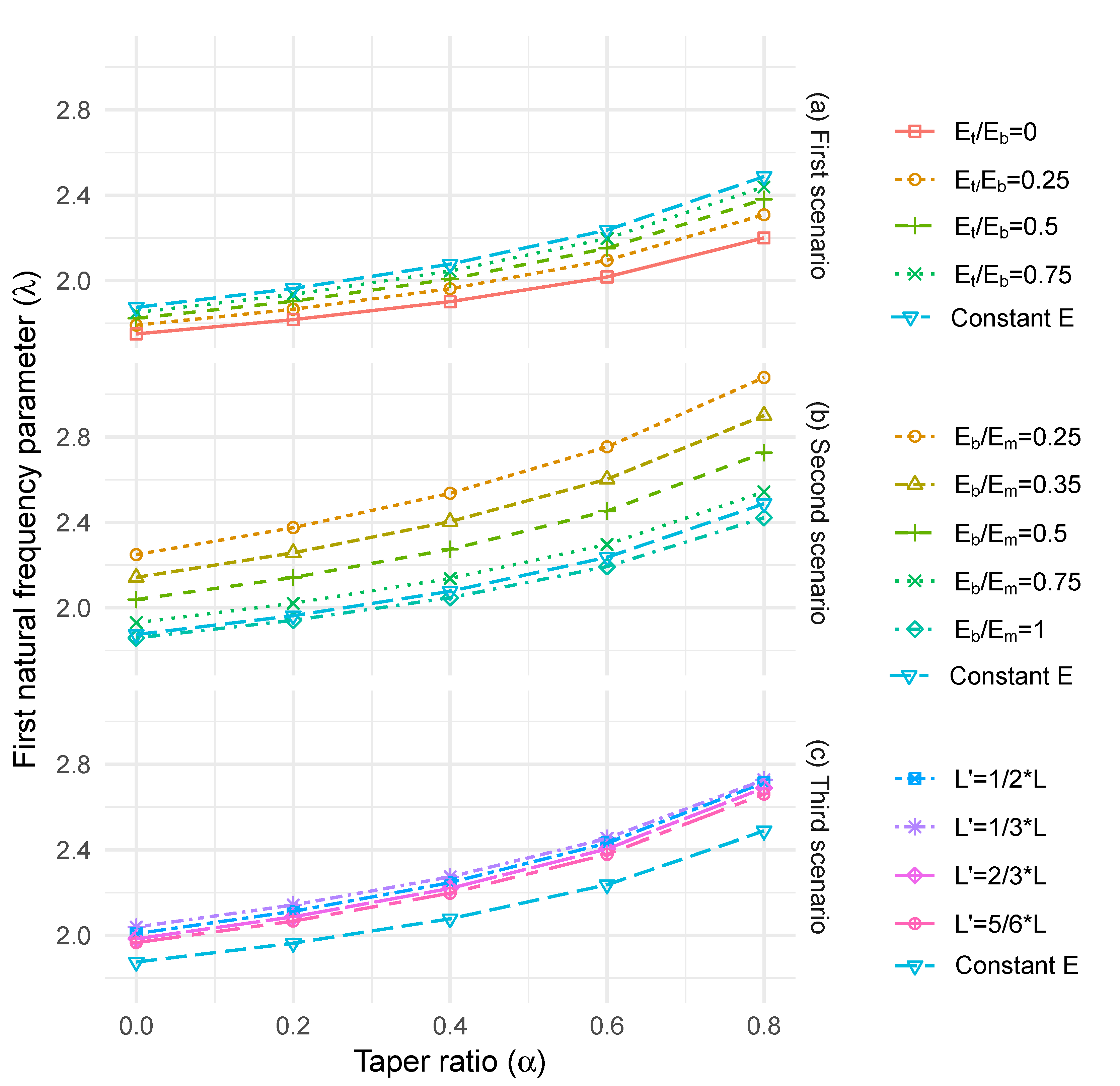
2.2.2. Effect of Longitudinal Stiffness Variation
2.2.3. Effect of Radial Stiffness Variation
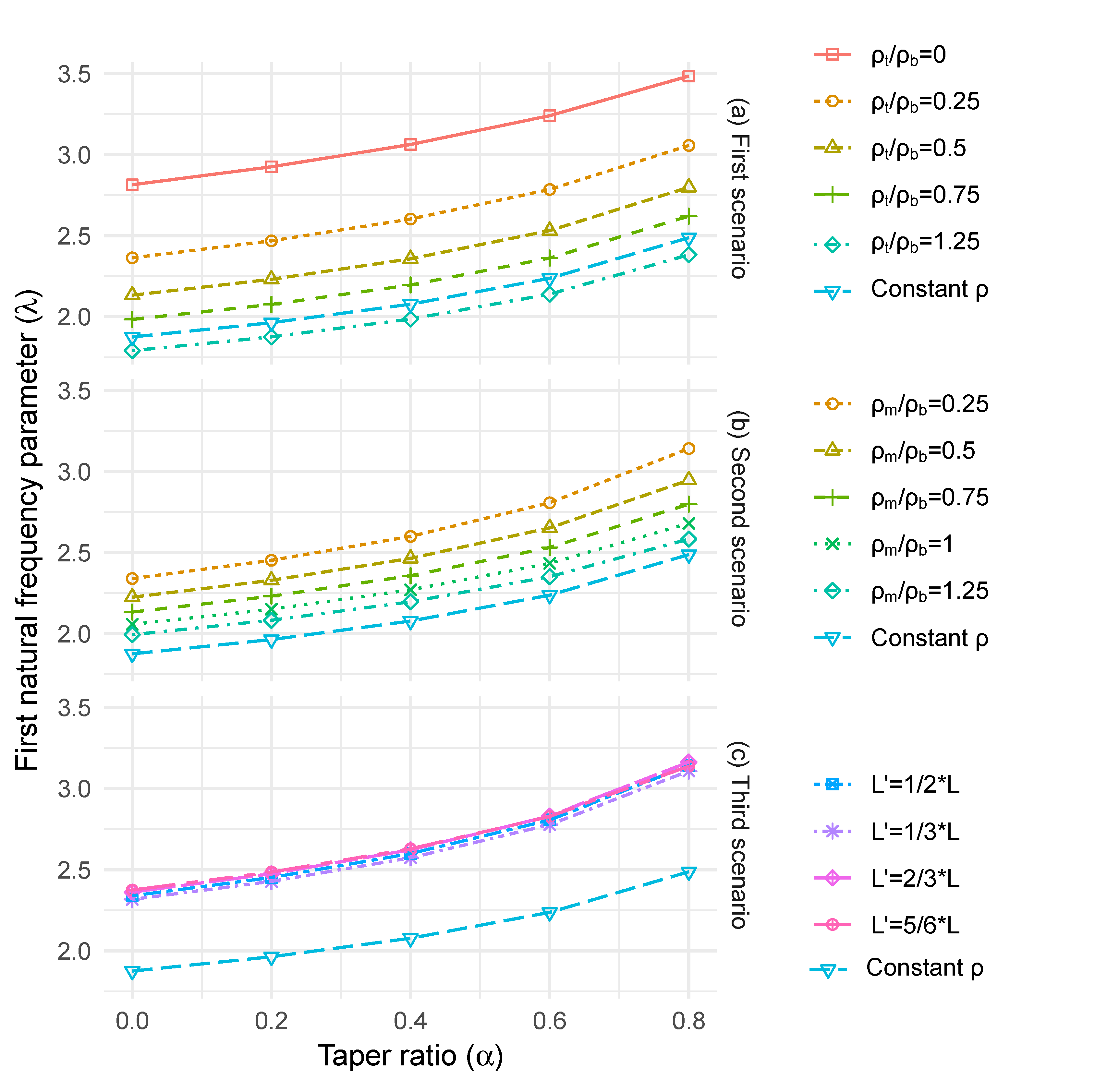
2.2.4. Effect of Longitudinal Density Variation
2.2.5. Effect of Radial Density Variation
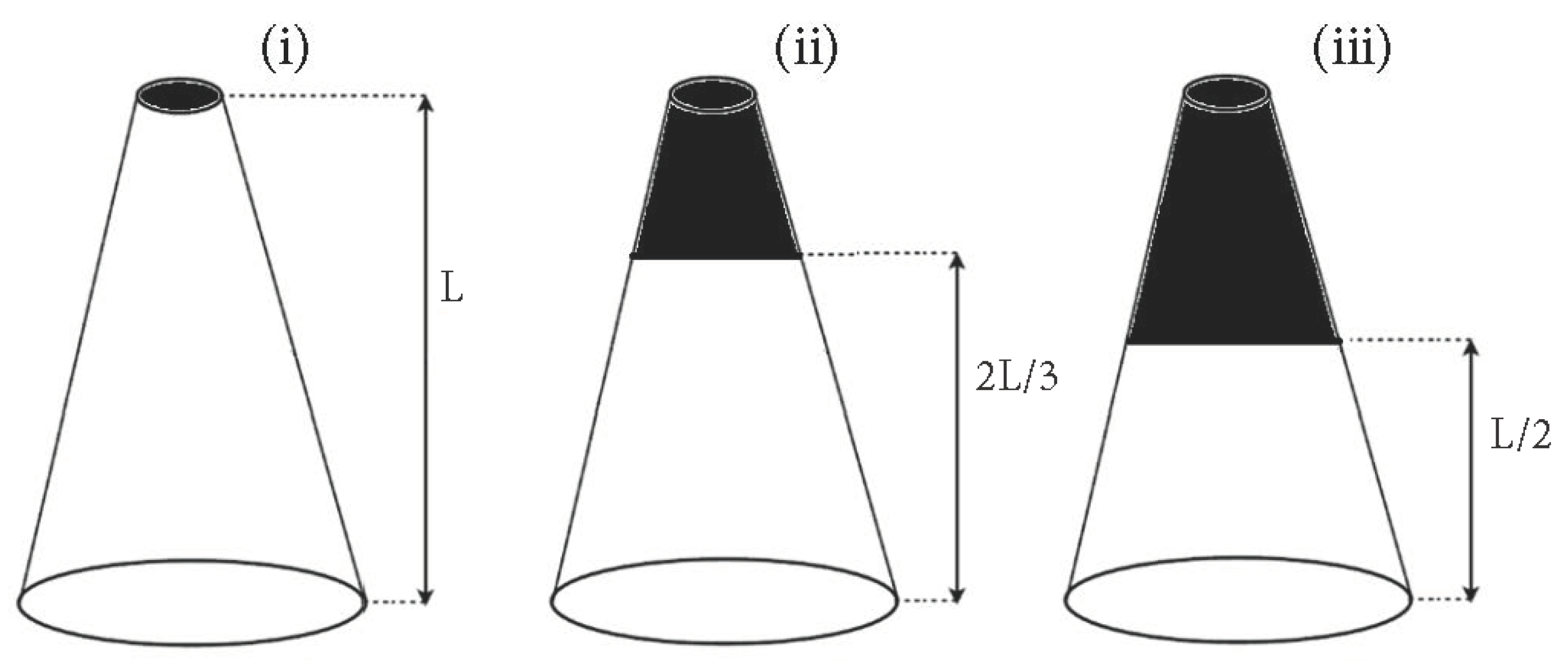
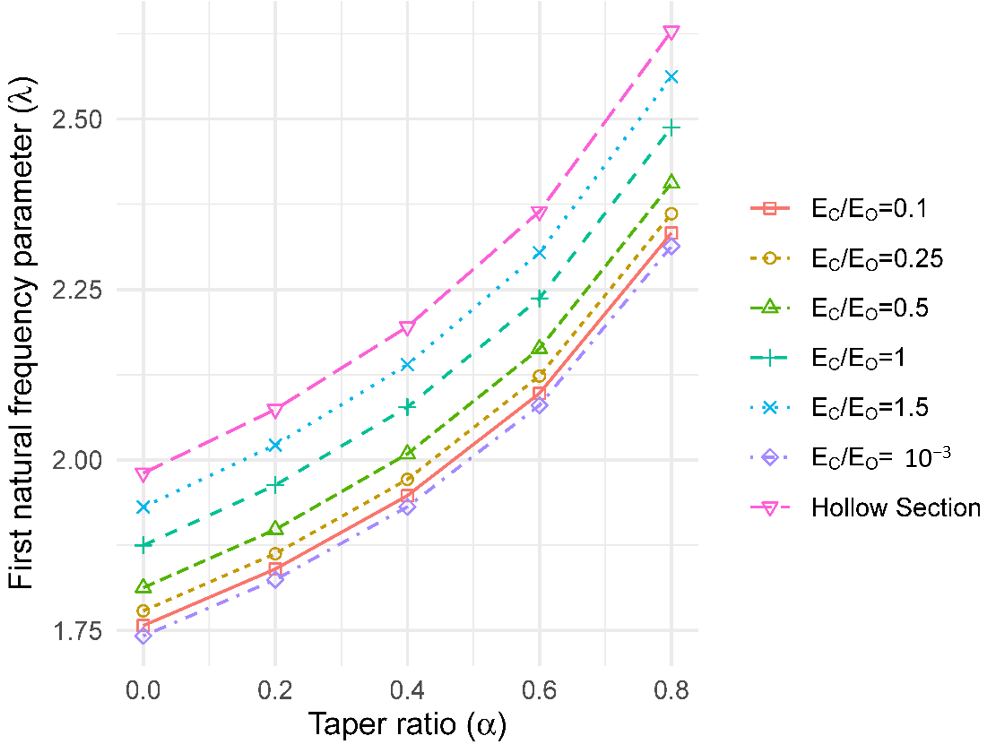
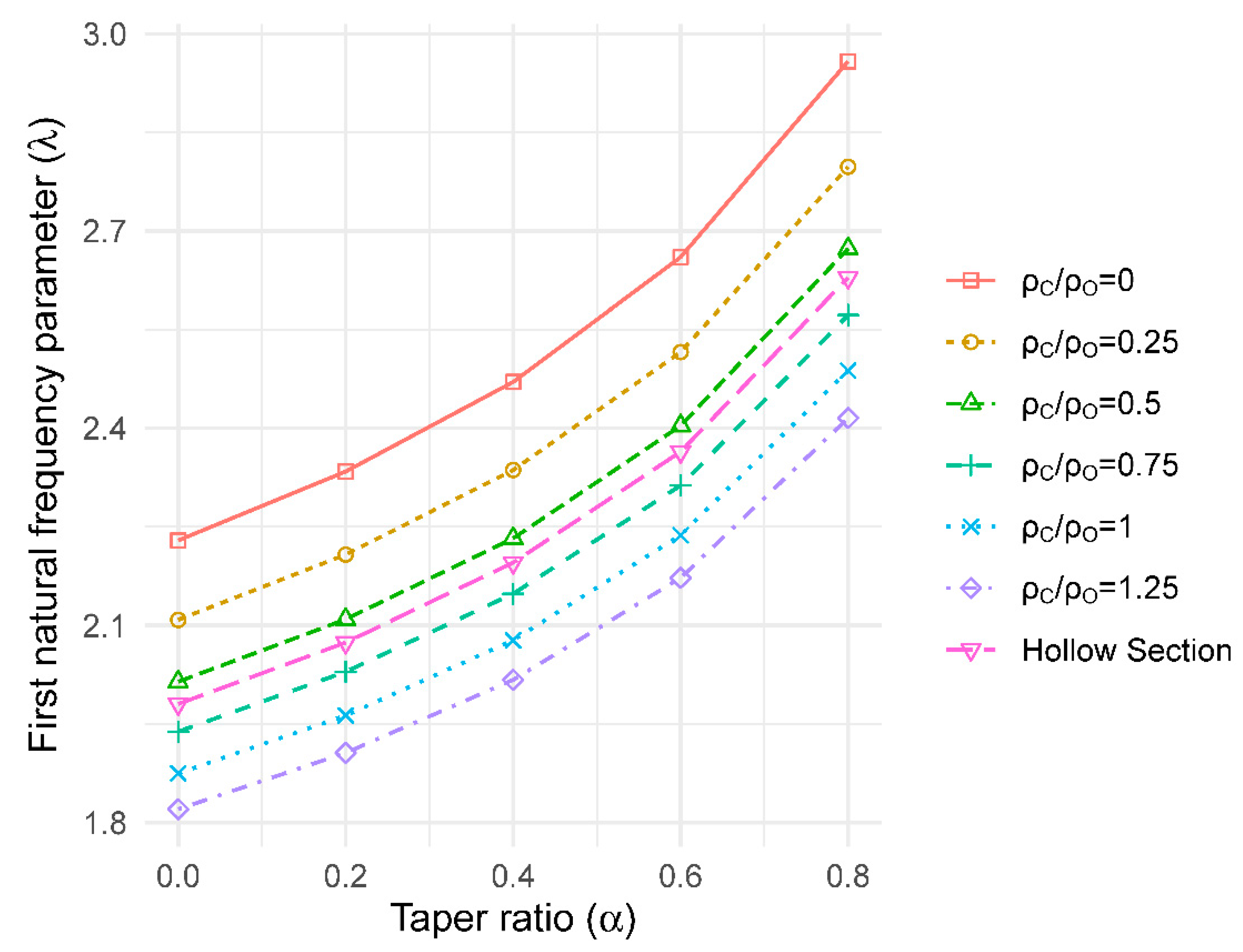
2.2.6. Effect of Crown Mass
2.3. Estimating Natural Frequency for Coniferous and Broadleaved Trees
3. Results
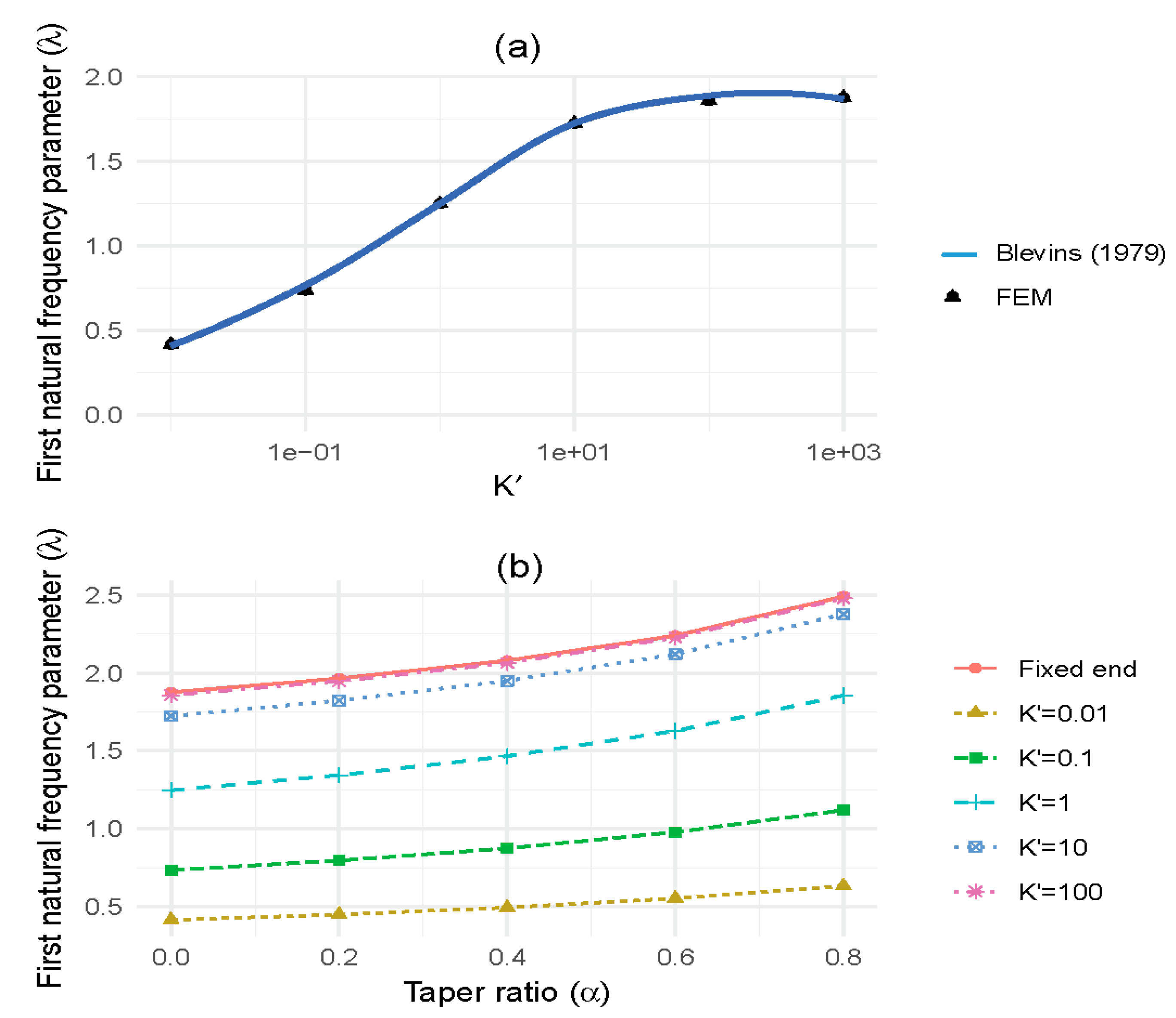
3.1. Effect of Base Fixity
3.2. Effect of Longitudinal Stiffness Variation
3.3. Effect of Radial Stiffness Variation
3.4. Effect of Longitudinal Density Variation
3.5. Effect of Radial Density Variation
3.6. Effect of Crown Mass
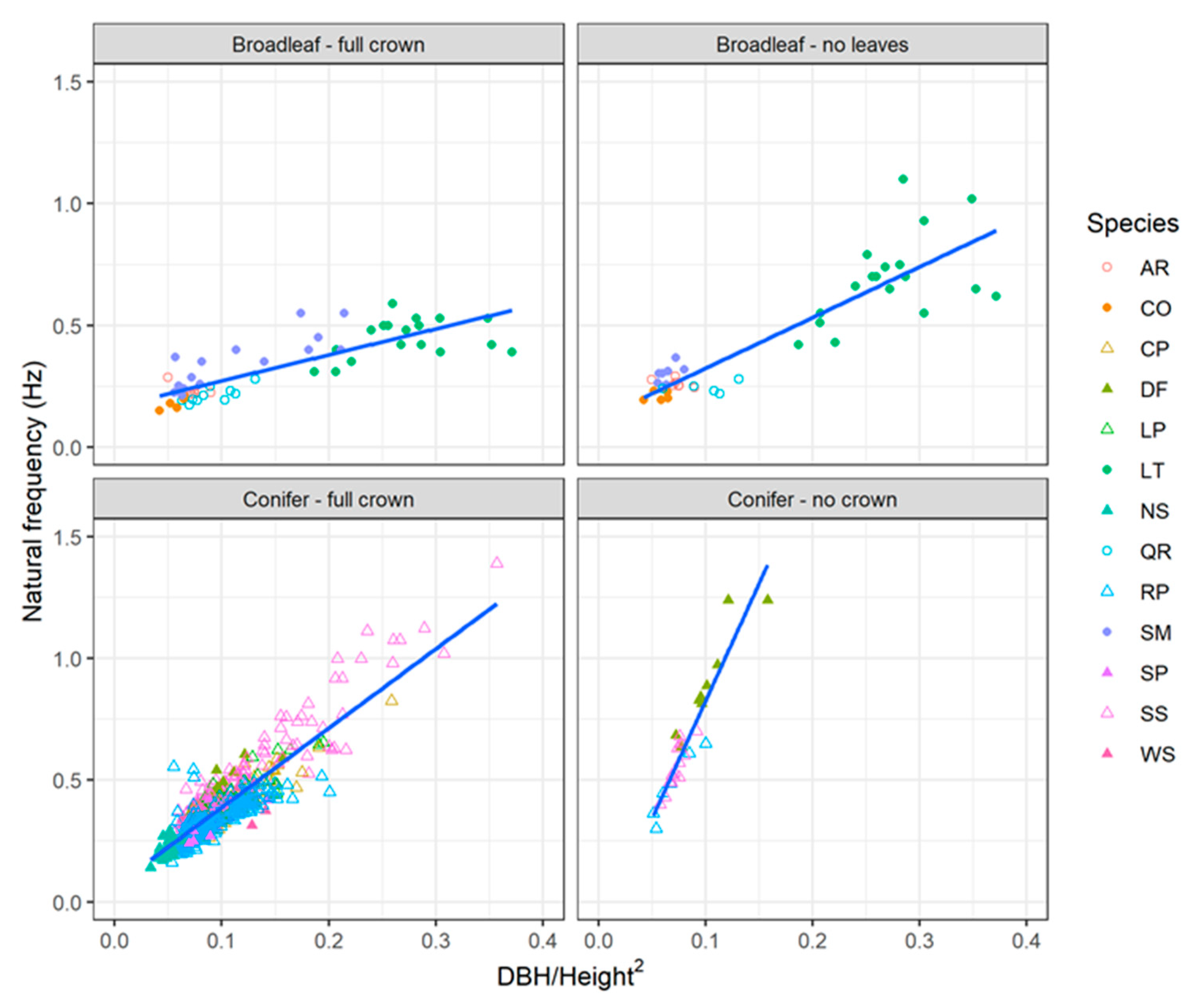
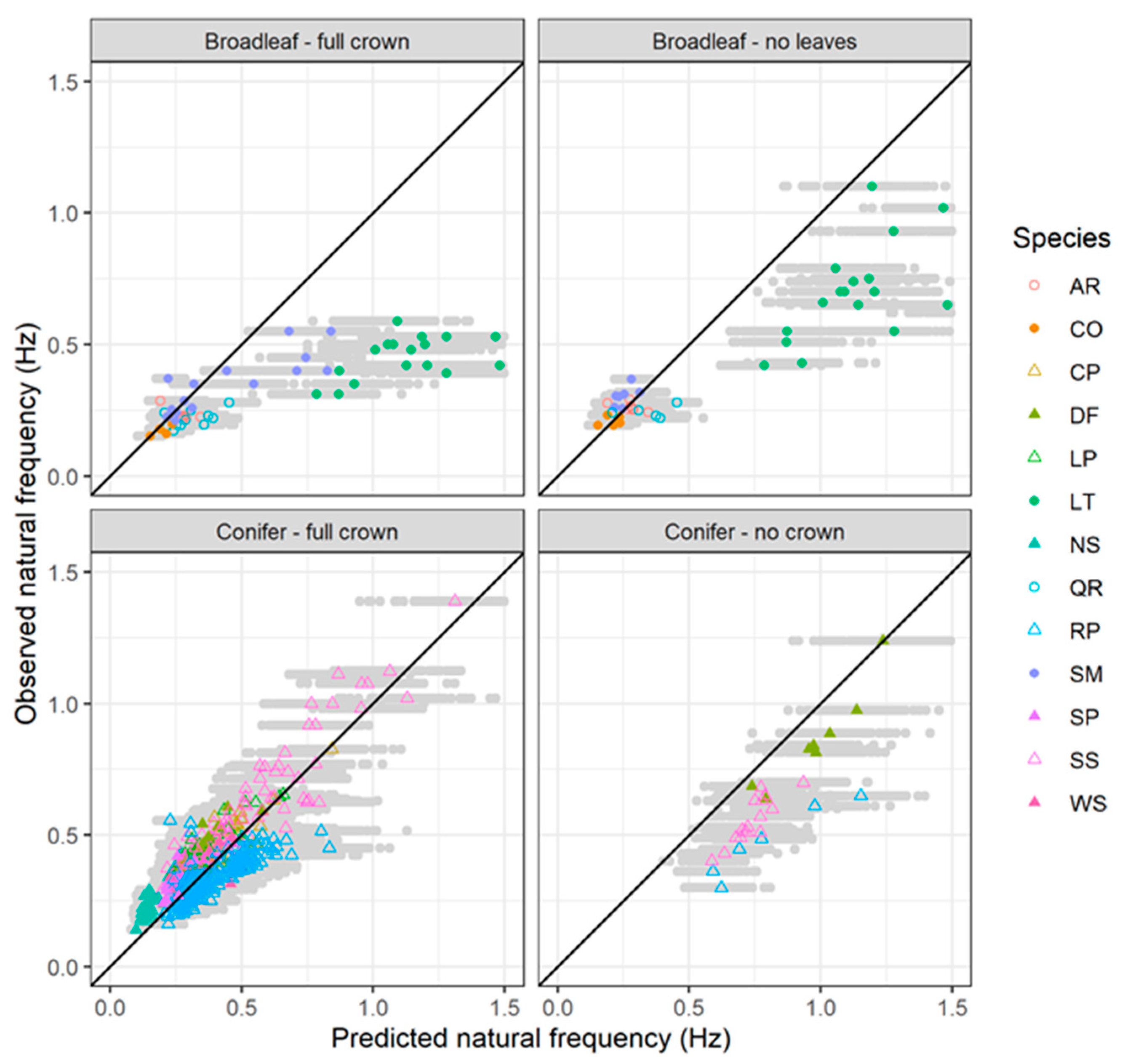
3.7. Estimating Natural Frequency for Coniferous and Broadleaved Trees
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- De Langre, E. Effects of wind on plants. Annu. Rev. Fluid Mech. 2008, 40, 141–168. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, B.; Berry, P.; Moulia, B. Review: Wind impacts on plant growth, mechanics and damage. Plant Sci. 2016, 245, 94–118. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.; Gardiner, B.; Sellier, D. Tree Mechanics and Wind Loading. In Plant Biomechanics; Geitmann, A., Gril, J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 79–106. ISBN 978-3-319-79098-5. [Google Scholar]
- Grace, J. Plant Response to Wind; Academic Press: London, UK, 1977. [Google Scholar]
- Telewski, F.W. Wind-induced physiological and developmental responses in trees. In Wind and Trees; Coutts, M.P., Grace, J., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 237–263. [Google Scholar]
- Moulia, B.; Coutand, C.; Julien, J.-L. Mechanosensitive control of plant growth: Bearing the load, sensing, transducing, and responding. Front. Plant Sci. 2015, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Fournier, M.; Dlouhá, J.; Jaouen, G.; Almeras, T. Integrative biomechanics for tree ecology: Beyond wood density and strength. J. Exp. Bot. 2013, 64, 4793–4815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardiner, B.A. Mathematical modelling of the static and dynamic characteristics of plantation trees. In Mathematical Modelling of Forest Ecosystems; Franke, J., Roeder, A., Eds.; Sauerlander’s Verlag: Frankfurt am Main, Germany, 1992; pp. 40–61. [Google Scholar]
- Gardiner, B.; Peltola, H.; Kellomäki, S. Comparison of two models for predicting the critical wind speeds required to damage coniferous trees. Ecol. Model. 2000, 129, 1–23. [Google Scholar] [CrossRef]
- Mayer, H. Wind-induced tree sways. Trees 1987, 1, 195–206. [Google Scholar] [CrossRef]
- Moore, J.R.; Maguire, D.A. Natural sway frequencies and damping ratios of trees: Concepts, review and synthesis of previous studies. Trees-Struct. Funct. 2004, 18, 195–203. [Google Scholar] [CrossRef]
- Jonsson, M.J.; Foetzki, A.; Kalberer, M.; Lundström, T.; Ammann, W.; Stöckli, V. Natural frequencies and damping ratios of Norway spruce (Picea abies (L.) Karst) growing on subalpine forested slopes. Trees 2007, 21, 541–548. [Google Scholar] [CrossRef]
- Blackwell, P.G.; Rennolls, K.; Coutts, M.P. A root anchorage model for shallowly rooted Sitka spruce. Forestry 1990, 63, 73–91. [Google Scholar] [CrossRef]
- Baker, C.J. Measurements of the natural frequencies of trees. J. Exp. Bot. 1997, 48, 1125–1132. [Google Scholar] [CrossRef]
- Moore, J.R.; Maguire, D.A. Natural sway frequencies and damping ratios of trees: Influence of crown structure. Trees 2005, 19, 363–373. [Google Scholar] [CrossRef]
- James, K.R.; Haritos, N.; Ades, P.K. Mechanical stability of trees under dynamic loads. Am. J. Bot. 2006, 93, 1522–1530. [Google Scholar] [CrossRef] [PubMed]
- Kane, B.; James, K.R. Dynamic properties of open-grown deciduous trees. Can. J. For. Res. 2011, 41, 321–330. [Google Scholar] [CrossRef]
- Horvath, E.; Sitkei, G. Energy consumption of selected tree shakers under different operational condition. J. Agric. Eng. Res. 2001, 80, 191–199. [Google Scholar] [CrossRef]
- Kerzenmacher, T.; Gardiner, B. A mathematical model to describe the dynamic response of a spruce tree to the wind. Trees 1998, 12, 385. [Google Scholar] [CrossRef]
- Saunderson, S.E.T.; England, A.H.; Baker, C.J. A Dynamic Model of the Behaviour of Sitka Spruce in High Winds. J. Theor. Biol. 1999, 200, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Sellier, D.; Fourcaud, T. A mechanical analysis of the relationship between free oscillations of Pinus pinaster Ait. saplings and their aerial architecture. J. Exp. Bot. 2005, 56, 1563–1573. [Google Scholar] [CrossRef] [Green Version]
- Sellier, D.; Fourcaud, T.; Lac, P. A finite element model for investigating effects of aerial architecture on tree oscillations. Tree Physiol. 2006, 26, 799–806. [Google Scholar] [CrossRef] [Green Version]
- Sellier, D.; Brunet, Y.; Fourcaud, T. A numerical model of tree aerodynamic response to a turbulent airflow. Forestry 2008, 81, 279–297. [Google Scholar] [CrossRef] [Green Version]
- Spatz, H.-C.; Bruchert, F.; Pfisterer, J. Multiple resonance damping or how do trees escape dangerously large oscillations? Am. J. Bot. 2007, 94, 1603–1611. [Google Scholar] [CrossRef]
- Moore, J.R.; Maguire, D.A. Simulating the dynamic behavior of Douglas-fir trees under applied loads by the finite element method. Tree Physiol. 2008, 28, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.; de Langre, E.; Moulia, B. A scaling law for the effects of architecture and allometry on tree vibration modes suggests a biological tuning to modal compartmentalization. Am. J. Bot. 2008, 95, 1523–1537. [Google Scholar] [CrossRef] [PubMed]
- Spatz, H.-C.; Speck, O. Oscillation frequencies of tapered plant stems. Am. J. Bot. 2002, 89, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milne, R. Dynamics of swaying of Picea sitchensis. Tree Physiol. 1991, 9, 383–399. [Google Scholar] [CrossRef]
- Humar, J.L. Dynamics of Structures, 3rd ed.; CRC Press-Taylor & Francis Croup: Boca Raton, FL, USA, 2012; ISBN 978-0-415-62086-4. [Google Scholar]
- Blevins, R.D. Formulas for Natural Frequency and Mode Shape; Van Nostrand Reinhold Co.: New York, NY, USA, 1979; ISBN 978-0-442-20710-6. [Google Scholar]
- Mayhead, G.J. Sway periods of forest trees. Scott. For. 1973, 27, 19–23. [Google Scholar]
- Jackson, T.; Shenkin, A.; Moore, J.; Bunce, A.; van Emmerik, T.; Kane, B.; Burcham, D.; James, K.; Selker, J.; Calders, K.; et al. An architectural understanding of natural sway frequencies in trees. J. R. Soc. Interface 2019, 16, 20190116. [Google Scholar] [CrossRef] [Green Version]
- Lachenbruch, B.; Moore, J.R.; Evans, R. Radial Variation in Wood Structure and Function in Woody Plants, and Hypotheses for Its Occurrence. In Size- and Age-Related Changes in Tree Structure and Function; Tree, Physiology; Meinzer, F.C., Lachenbruch, B., Dawson, T.E., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 4, pp. 121–164. ISBN 978-94-007-1241-6. [Google Scholar]
- Niklas, K.J. Plant Biomechanics: An Engineering Approach to Plant Form and Function; University of Chicago Press: Chicago, IL, USA, 1992; ISBN 978-0-226-58630-4. [Google Scholar]
- Niklas, K. Mechanical properties of black locust (Robinia pseudoacacia L.) wood. Size- and age-dependent variations in sap- and heartwood. Ann. Bot. 1997, 79, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Koch, P. Utilization of Hardwoods Growing on Southern Pine Sites-Volume 1; USDA-Forest Service, Southern Forest Experiment Station: Asheville, NC, USA, 1985.
- Rich, P.M. Mechanical structure of the stem of arborescent palms. Bot. Gaz. 1987, 148, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Wiemann, M.C.; Williamson, G.B. Wood specific gravity gradients in tropical dry and montane rain forest trees. Am. J. Bot. 1989, 76, 924–928. [Google Scholar] [CrossRef]
- Xu, P.; Walker, J.C.F. Stiffness gradients in radiata pine trees. Wood Sci. Technol. 2004, 38, 1–9. [Google Scholar] [CrossRef]
- Waghorn, M.J.; Mason, E.G.; Watt, M.S. Influence of initial stand density and genotype on longitudinal variation in modulus of elasticity for 17-year-old Pinus radiata. Forest Ecol. Manag. 2007, 252, 67–72. [Google Scholar] [CrossRef]
- Hirakawa, Y.; Fujisawa, Y. The S2 microfibril angle variations in the vertical direction of latewood tracheids in sugi (Cryptomeria japonica) trees. Mokuzai Gakkaishi/J. Jpn. Wood Res. Soc. 1996, 42, 107–114. [Google Scholar]
- Perstorper, M. Lengthwise variation in grading parameters and comparison with bending strength tests. In Proceedings of the 10th International Symposium on Nondestructive Testing of Wood; Santoz, J.L., Ed.; Presses Polytechniques et Universitaires Romandes: Lausanne, Switzerland, 1996; pp. 341–350. [Google Scholar]
- Gardiner, B.; Leban, J.-M.; Auty, D.; Simpson, H. Models for predicting wood density of British-grown Sitka spruce. Forestry 2011, 84, 119–132. [Google Scholar] [CrossRef]
- Auty, D.; Achim, A.; Macdonald, E.; Cameron, A.D.; Gardiner, B.A. Models for predicting wood density variation in Scots pine. Forestry 2014, 87, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Kimberley, M.O.; Cown, D.J.; McKinley, R.B.; Moore, J.R.; Dowling, L.J. Modelling variation in wood density within and among trees in stands of New Zealand-grown radiata pine. N. Z. J. For. Sci. 2015, 45, 22. [Google Scholar] [CrossRef] [Green Version]
- ght loading. For. Int. J. For. Res. 2019, 92, 393–405. [CrossRef]
- Hushmand, B. Experimental Studies of Dynamic Response of Foundations. Ph.D. Thesis, California Institute of Technology, Pasadena, CA, USA, 1983. [Google Scholar]
- Gardiner, B.A.; Stacey, G.R.; Belcher, R.E.; Wood, C.J. Field and wind tunnel assessments of the implications of respacing and thinning for tree stability. Forestry 1997, 70, 233–252. [Google Scholar] [CrossRef]
- Bunce, A.; Volin, J.C.; Miller, D.R.; Parent, J.; Rudnicki, M. Determinants of tree sway frequency in temperate deciduous forests of the Northeast United States. Agric. For. Meteorol. 2019, 266, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Flesch, T.K.; Wilson, J.D. Wind and remnant tree sway in forest cutblocks. II. Relating measured tree sway to wind statistics. Agric. For. Meteorol. 1999, 93, 243–258. [Google Scholar] [CrossRef] [Green Version]
- Webb, V.A.; Rudnicki, M.; Muppa, S.K. Analysis of tree sway and crown collisions for managed Pinus resinosa in southern Maine. For. Ecol. Manag. 2013, 302, 193–199. [Google Scholar] [CrossRef]
- Sugden, M.J. Tree sway period—A possible new parameter for crown classification and stand competition. For. Chron. 1962, 38, 336–344. [Google Scholar] [CrossRef]
- Kane, B.; Modarres-Sadeghi, Y.; James, K.R.; Reiland, M. Effects of crown structure on the sway characteristics of large decurrent trees. Trees 2014, 28, 151–159. [Google Scholar] [CrossRef]
- Lin, L.I.-K. A Concordance Correlation Coefficient to Evaluate Reproducibility. Biometrics 1989, 45, 255. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.I.-K. A note on the concordance correlation coefficient. Biometrics 2000, 56, 324–325. [Google Scholar] [CrossRef]
- McCurdy, D.R.; Spangenberg, W.G.; Doty, C.P. How to Choose Your Tree: A Guide to Parklike Landscaping in Illinois, Indiana, and Ohio; Southern Illinois University Press: Carbondale, IL, USA, 1972; ISBN 978-0-8093-0514-8. [Google Scholar]
- Cline, M.G. Apical dominance. Bot. Rev. 1991, 57, 318–358. [Google Scholar] [CrossRef]
- Ford, E.D. Branching, crown structure and the control of timber production. In Attributes of Trees as Crop Plants; Cannell, M.G.R., Jackson, J.E., Eds.; Institute of Terrestrial Ecology: Huntingdon, UK, 1985; pp. 228–252. [Google Scholar]
- Scannell, B. Quantification of the Interactive Motions of the Atmospheric Surface Layer and a Conifer Canopy. Ph.D. Thesis, Cranfield Institute of Technology, Bedford, UK, 1983. [Google Scholar]
- Moore, J.R. Mechanical Behavior of Coniferous Trees Subjected to Wind Loading. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 2002. [Google Scholar]
- Ciftci, C.; Brena, S.F.; Kane, B.; Arwade, S.R. The effect of crown architecture on dynamic amplification factor of an open-grown sugar maple (Acer saccharum L.). Trees 2013, 27, 1175–1189. [Google Scholar] [CrossRef]
- Antony, F.; Jordan, L.; Schimleck, L.R.; Clark, A.; Souter, R.A.; Daniels, R.F. Regional variation in wood modulus of elasticity (stiffness) and modulus of rupture (strength) of planted loblolly pine in the United States. Can. J. For. Res. 2011, 41, 1522–1533. [Google Scholar] [CrossRef]
- Kimberley, M.O.; Moore, J.R.; Dungey, H.S. Modelling the effects of genetic improvement on radiata pine wood density. N. Z. J. For. Sci. 2016, 46, 8. [Google Scholar] [CrossRef] [Green Version]
- Neild, S.A.; Wood, C.J. Estimating stem and root-anchorage flexibility in trees. Tree Physiol. 1999, 19, 141–151. [Google Scholar] [CrossRef]
- Borowicka, H. Uber ausmittig belaste starre Platten auf elastischisotorpem Untergrund. Ing. Arch. Berl. 1943, 1, 1–8. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Tree Species | n | Eg (GPa) | ρ (kg/m3) | Eg/ρ | Taper (α) | Wc/Ws |
|---|---|---|---|---|---|---|---|
| CP | Corsican pine (Pinus nigra) | 57 | 8.70 | 657 | 0.013 | 0.85 | 0.34 |
| DF | Douglas-fir (Pseudotsuga menziesii) | 17 | 9.83 | 583 | 0.017 | 0.77 | 0.16 |
| LP | Lodgepole pine (Pinus contorta) | 40 | 6.90 | 487 | 0.014 | 0.83 | 0.33 |
| NS | Norway spruce (Picea abies) | 32 | 6.23 | 598 | 0.010 | 0.90 | 0.32 |
| RP | Red pine (Pinus resinosa) | 300 | 8.80 | 410 | 0.021 | 0.81 | 0.22 |
| SP | Scots pine (Pinus sylvestris) | 20 | 7.33 | 700 | 0.010 | 0.90 | 0.29 |
| SS | Sitka spruce (Picea sitchensis) | 175 | 7.53 | 447 | 0.017 | 0.84 | 0.50 |
| WS | White spruce (Picea glauca) | 6 | 7.40 | 466 | 0.016 | 0.91 | 0.34 |
| Averages | 7.77 | 551 | 0.015 | 0.85 | 0.31 | ||
| LT | Lime (Tilia europaea) | 18 | 11.7 | 530 | 0.022 | 0.84 | 0.18 |
| AR | Red maple (Acer rubrum) | 7 | 9.6 | 524 | 0.018 | 0.67 | 0.22 |
| QR | Red oak (Quercus rubra) | 11 | 9.9 | 665 | 0.015 | 0.75 | 0.32 |
| CO | Shagbark hickory (Carya ovata) | 5 | 10.8 | 649 | 0.017 | 0.76 | 0.39 |
| SM | Sugar maple (Acer saccharum) | 15 | 10.7 | 560 | 0.019 | 0.79 | 0.18 |
| Averages | 10.54 | 585.6 | 0.018 | 0.76 | 0.26 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dargahi, M.; Newson, T.; R. Moore, J. A Numerical Approach to Estimate Natural Frequency of Trees with Variable Properties. Forests 2020, 11, 915. https://doi.org/10.3390/f11090915
Dargahi M, Newson T, R. Moore J. A Numerical Approach to Estimate Natural Frequency of Trees with Variable Properties. Forests. 2020; 11(9):915. https://doi.org/10.3390/f11090915
Chicago/Turabian StyleDargahi, Mojtaba, Timothy Newson, and John R. Moore. 2020. "A Numerical Approach to Estimate Natural Frequency of Trees with Variable Properties" Forests 11, no. 9: 915. https://doi.org/10.3390/f11090915
APA StyleDargahi, M., Newson, T., & R. Moore, J. (2020). A Numerical Approach to Estimate Natural Frequency of Trees with Variable Properties. Forests, 11(9), 915. https://doi.org/10.3390/f11090915