Using a Trait-Based Approach to Compare Tree Species Sensitivity to Climate Change Stressors in Eastern Canada and Inform Adaptation Practices

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
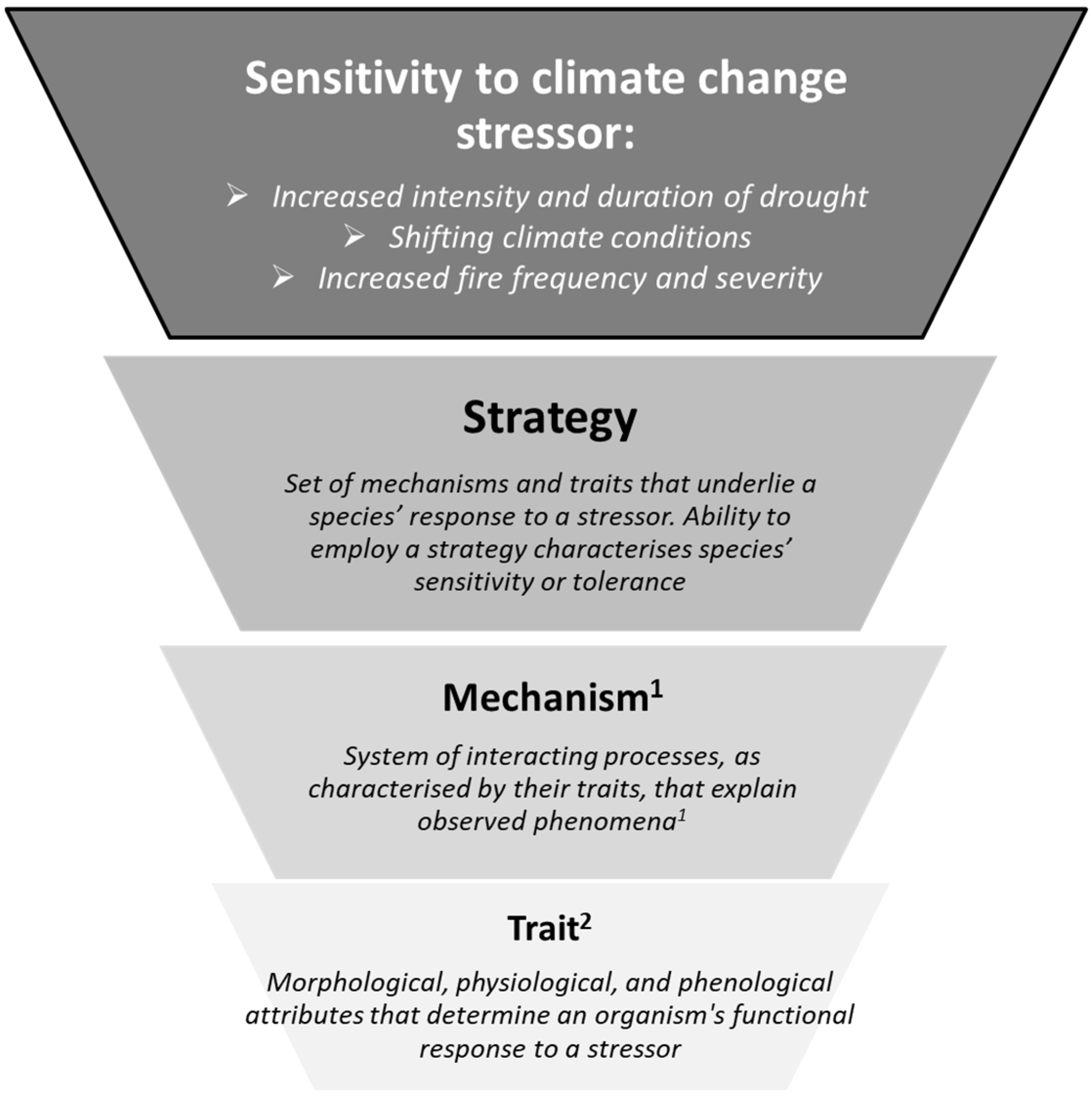
2.1. Trait Documentation
2.2. Index Development
- (1)
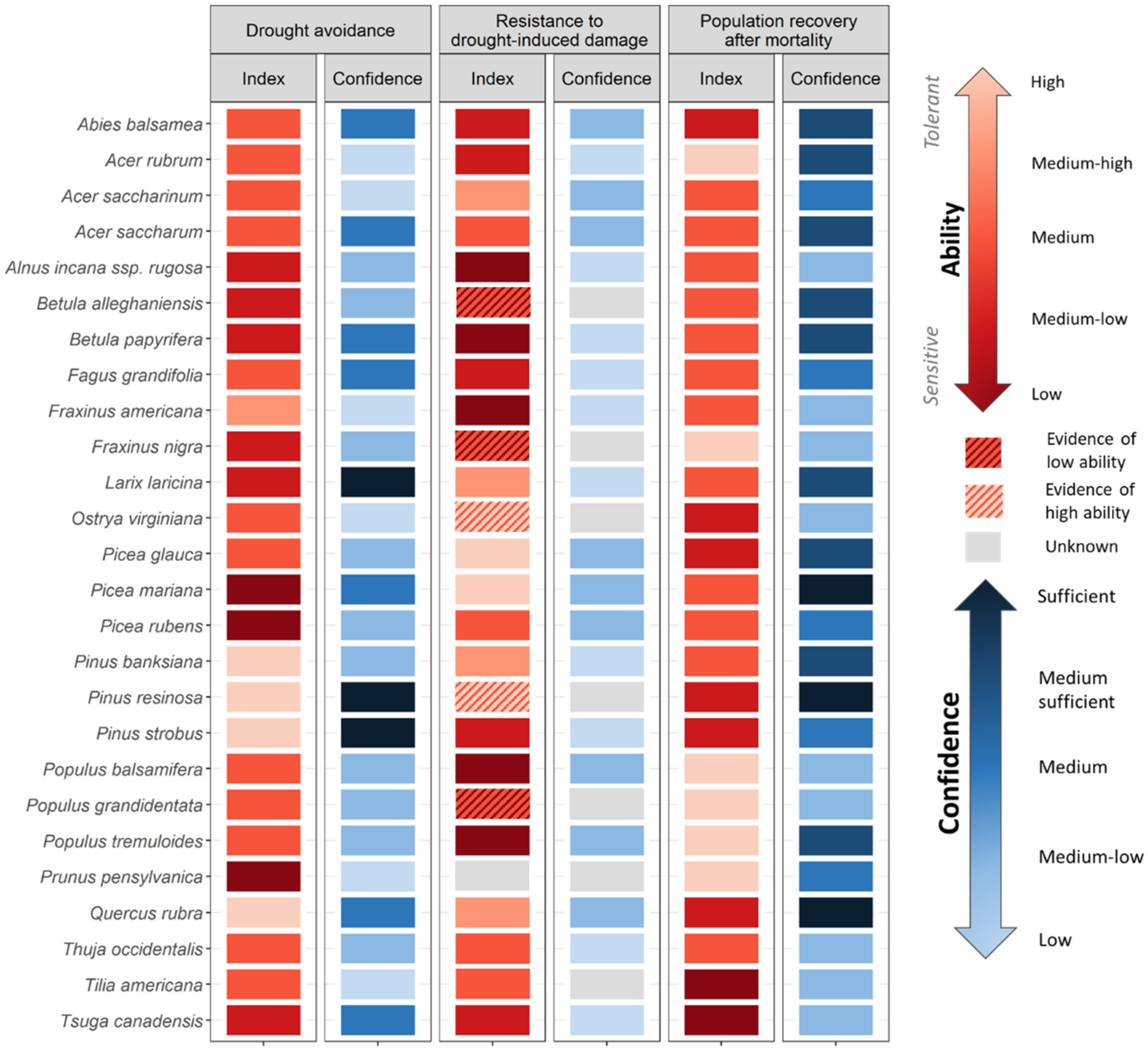
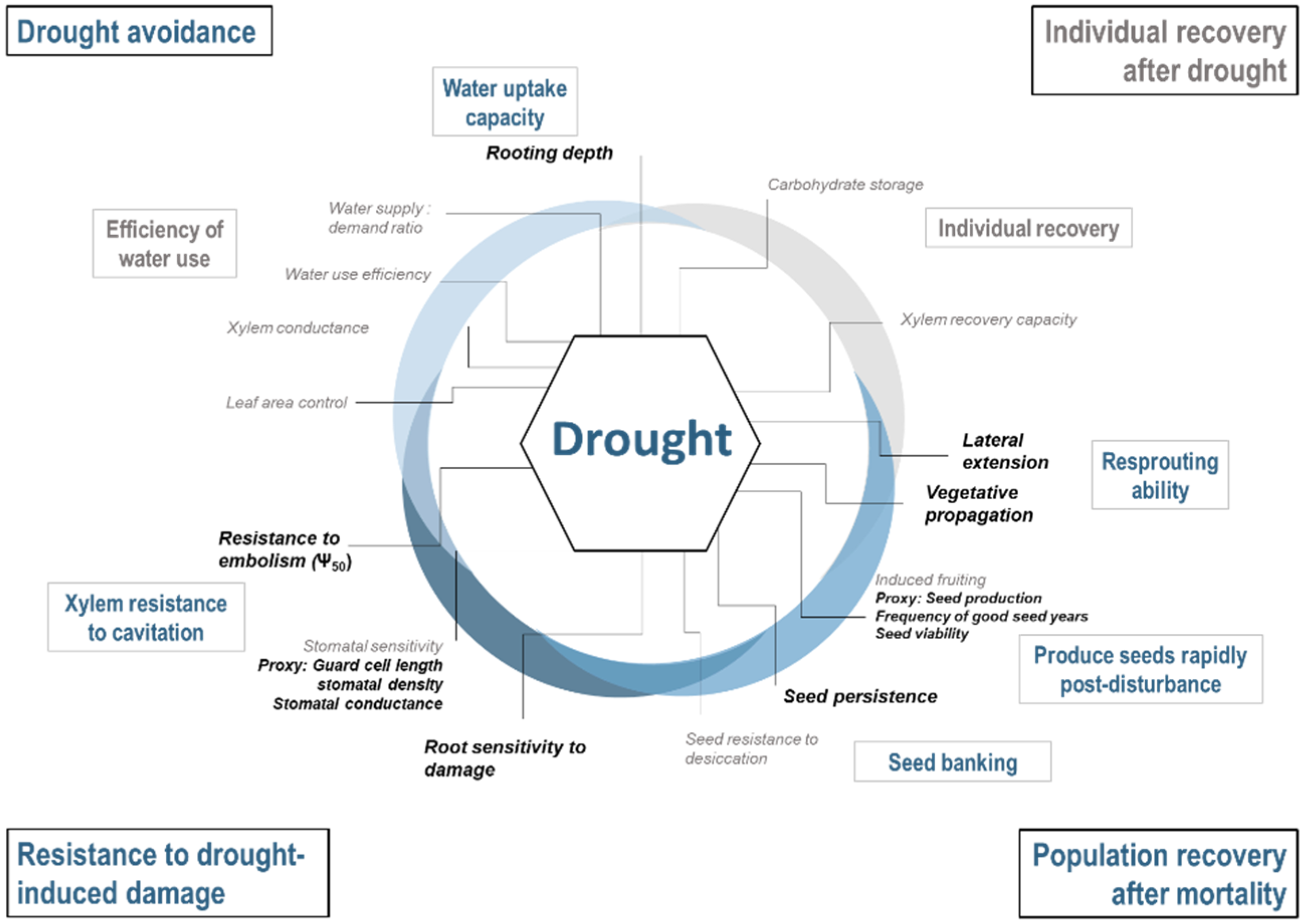
- Drought avoidance by maintaining internal water levels: As the water levels drop at the soil surface during drought, continued access to the water and uptake capacity from deeper horizons are crucial to avoid drought (Breda et al. 2006). Shallow rooting species will therefore be the most sensitive.
- (2)
- Resistance to drought-induced damage by maintaining circulation when internal water levels are low: When water access decreases but demand remains constant, mechanisms such as xylem resistance to cavitation become important to avoid mortality. Ψ50 (xylem water potential at which 50% of conductivity is lost) is a widely used metric of xylem resistance [45,46]. Traits that characterize stomatal sensitivity are also important to consider because they underlie species’ ability to limit water loss during prolonged droughts [47].
- (3)
- Population recovery after mortality: the ability of a species to rapidly recolonize an area after drought-induced mortality will influence its persistence in the landscape. Population may recover from several mechanisms including resprouting ability, ability to produce seed rapidly post-disturbance and to store seeds in the seed bank. Traits related to seedling survival are also of importance but was not considered here because of the lack of data on ontological differences in trait expression.
- (4)
- Individual recovery after drought: Several mechanisms influence the ability of individual stems to recover after drought. This includes the capacity to resume hydraulic conductivity after xylem have been embolised and/or produce new conductive tissues [48]. However, questions remain regarding which physiological mechanisms are responsible [49]. Survival may also be influenced by non-structural carbohydrate storage (NSC; [50]), though the exact relationship between NSC and recovery remains an active area of research [51,52,53]. Therefore, we did not develop an index for this strategy.
- (1)
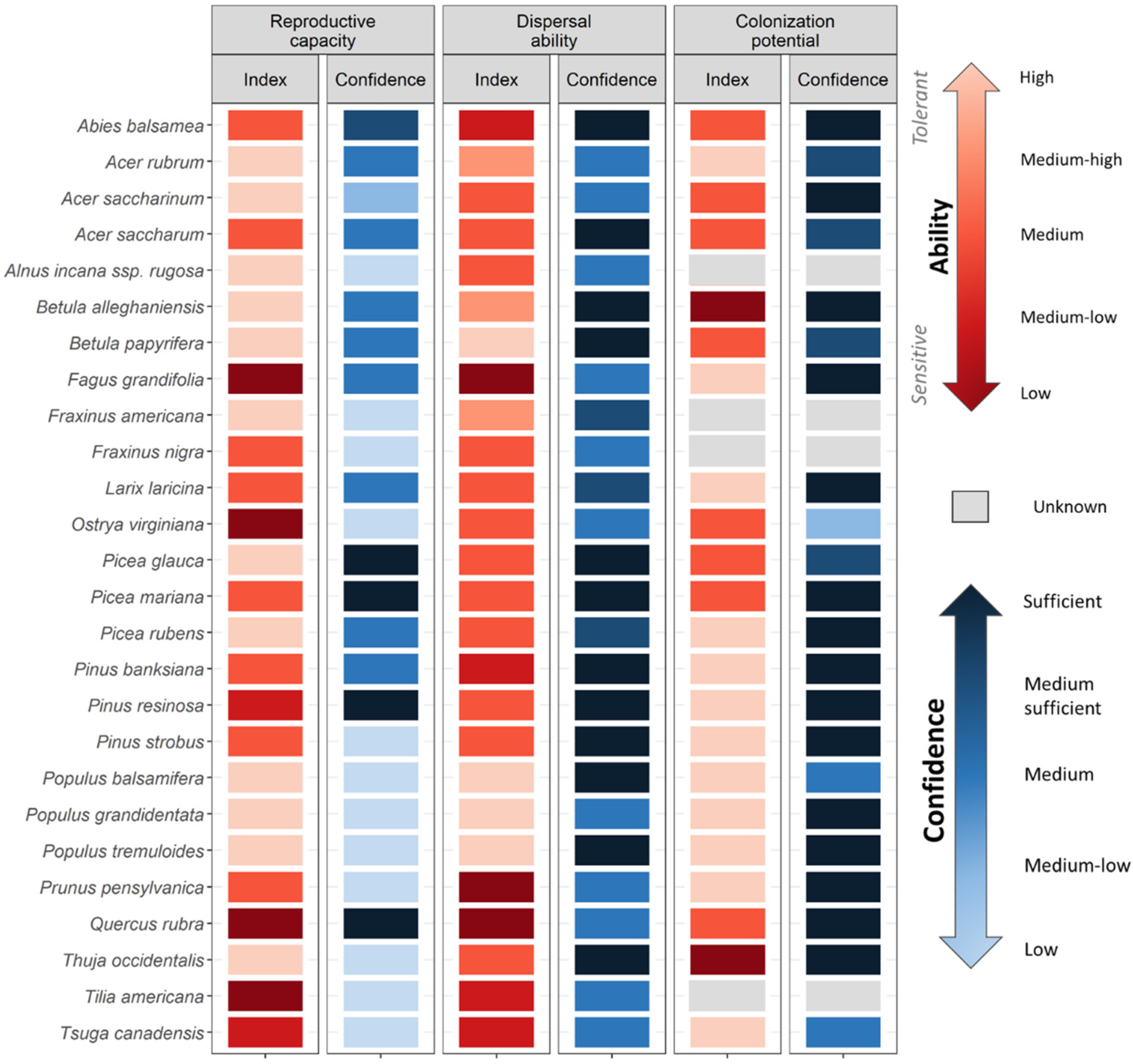
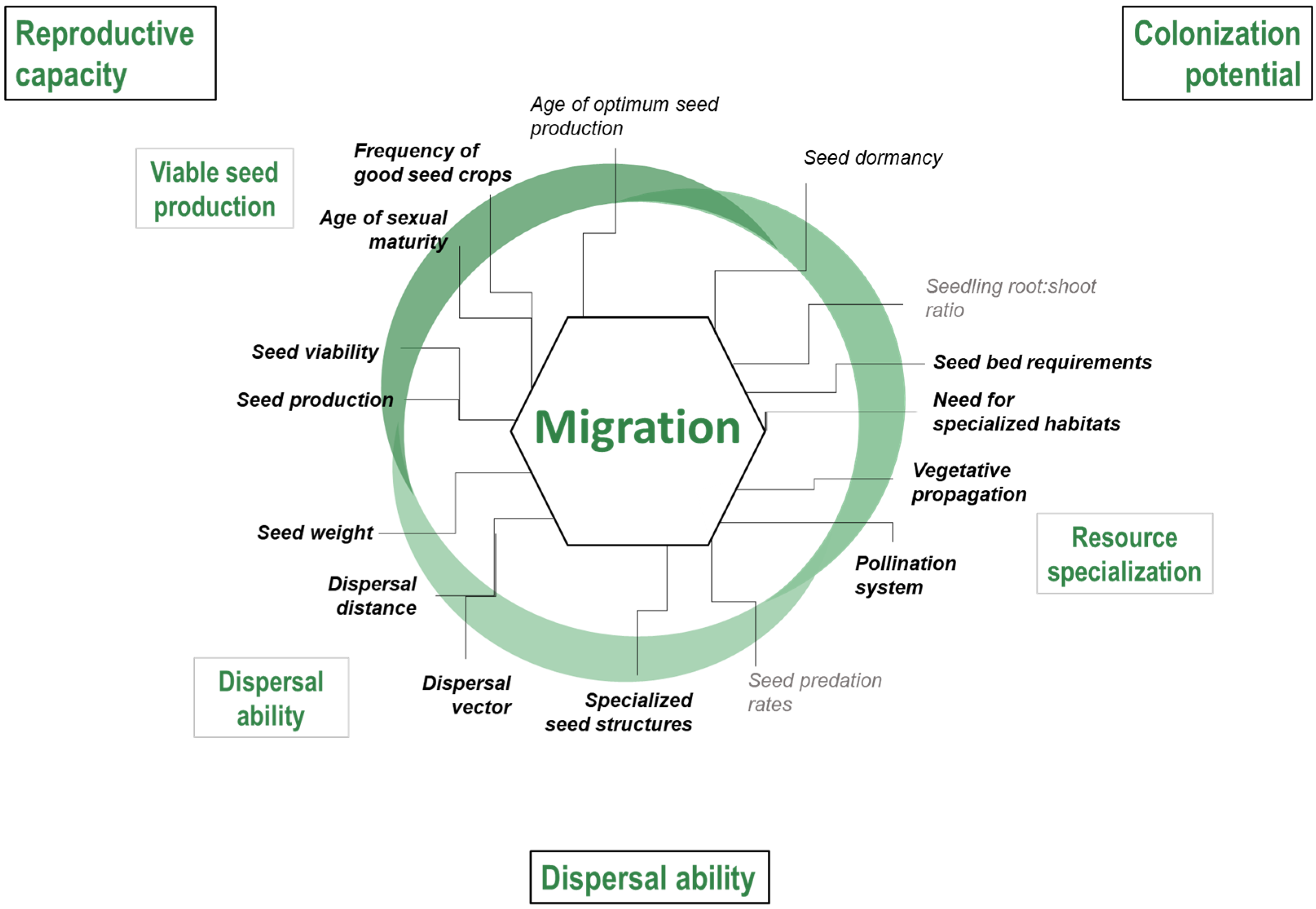
- Reproductive capacity: Successful migration is dependent on the reproductive capacity of the source population as well as the time to reach sexual maturity. Propagule pressure, determined by seed production, is an important determinant of the success of recruitment, locally and at their advancing front [57,58,59].
- (2)
- (3)
- Colonization potential at the advancing front: Once dispersules reach a new site, their ability to germinate, survive, and reproduce will largely determine which species can colonize [63]. Once colonised, species’ ability to tolerate inbreeding and successfully reproduce or propagate in small populations will play a critical role in its migration success [64,65].
- (1)
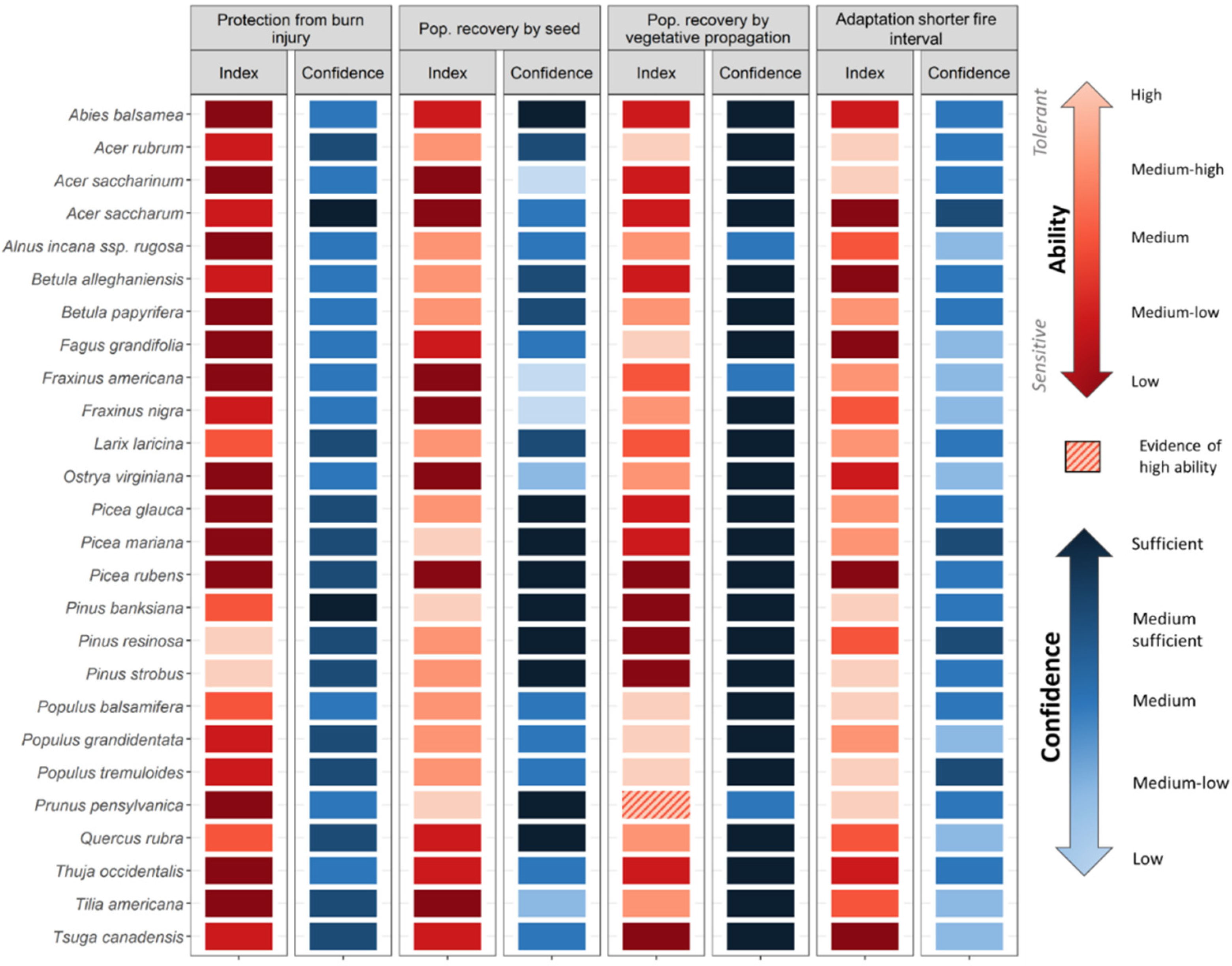
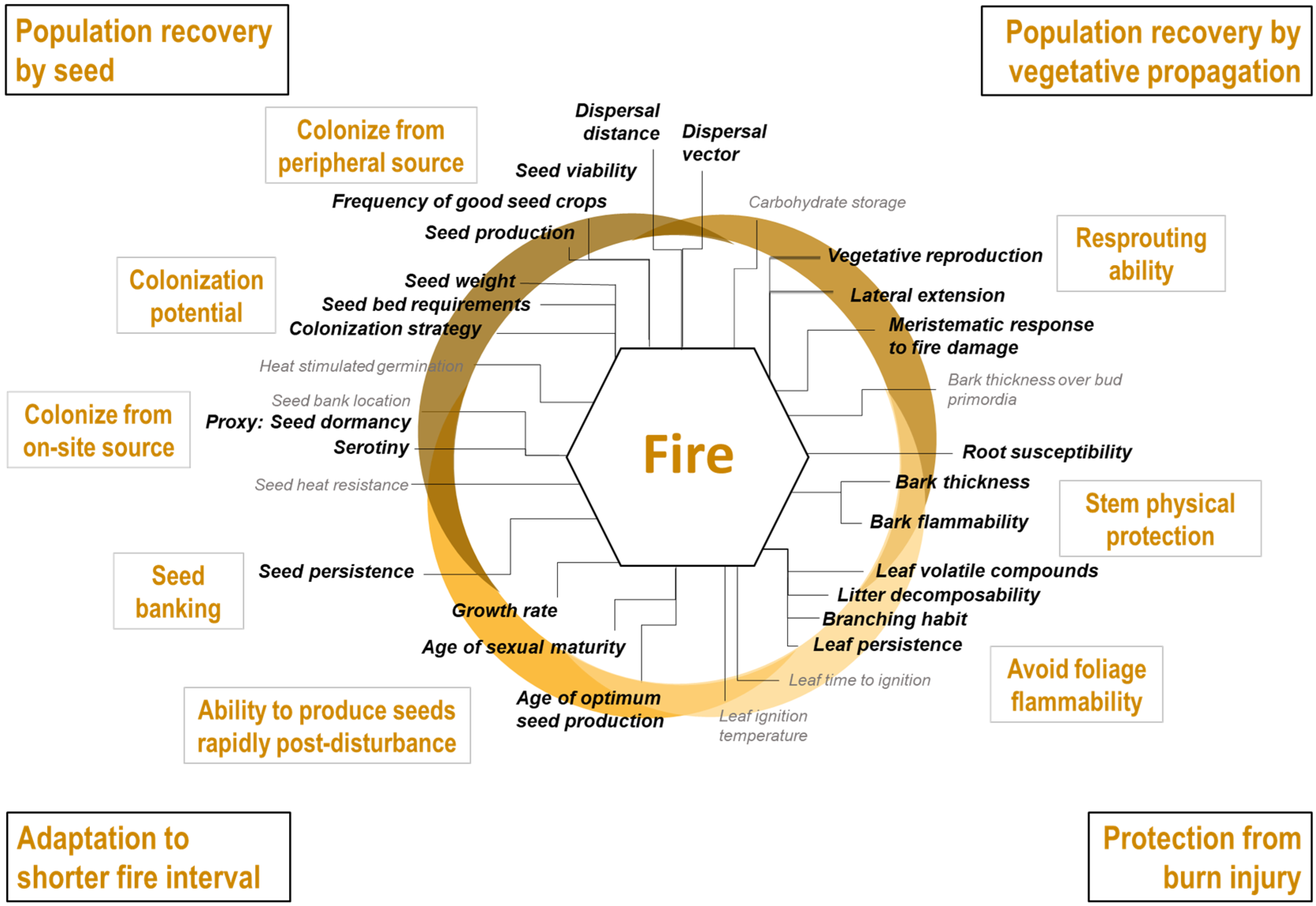
- Stem protection from burn injury: Surviving stems are contingent on maintaining intact vascular systems capable of circulating water and sap. Stem physical protection, or the ability to reduce heat transmission through the outer layers to the vascular system (e.g., thick bark), influences whether a stem will survive or not [68]. Additionally, certain leaf traits and crown properties influence flammability and may affect fire intensity and spread [69].
- (2)
- Population recovery by seed, from seed sources on-site or from peripheral areas not affected by the fire. Protecting seeds from fire, i.e., through aerial or soil banks or by protective structures, ensures a direct source of dispersules to re-establish populations [70,71]. When species lack adaptations to protect seeds or when subjected to very intense fires, seeds must originate from the unburnt forest along the edge and beyond, making dispersal ability and seed production crucial mechanisms [72]. Germination requirements also influence post-fire recruitment [73].
- (3)
- Population recovery by vegetative propagation. Resprouting ability is determined mainly by the location of meristematic tissues on the plant and how well protected these tissues are [74]. Like seeds in the soil bank, underground buds may be protected in the soil and consequently have higher survival probabilities, especially in low-intensity fires. The type of vegetative reproduction will also influence the rapidity of population recovery, particularly for species with extensive clonality [75].
- (4)
- Adaptation to shorter fire intervals. Multiple fires in a short time frame may prevent species from re-establishing from seed, even for fire-adapted species [76]. Hence, short fire return intervals favour species that can mature and start to produce seed rapidly after a fire has occurred. Seeds that remain viable for long periods in the soil bank could also provide a source of propagules [70].
2.3. Confidence Scores
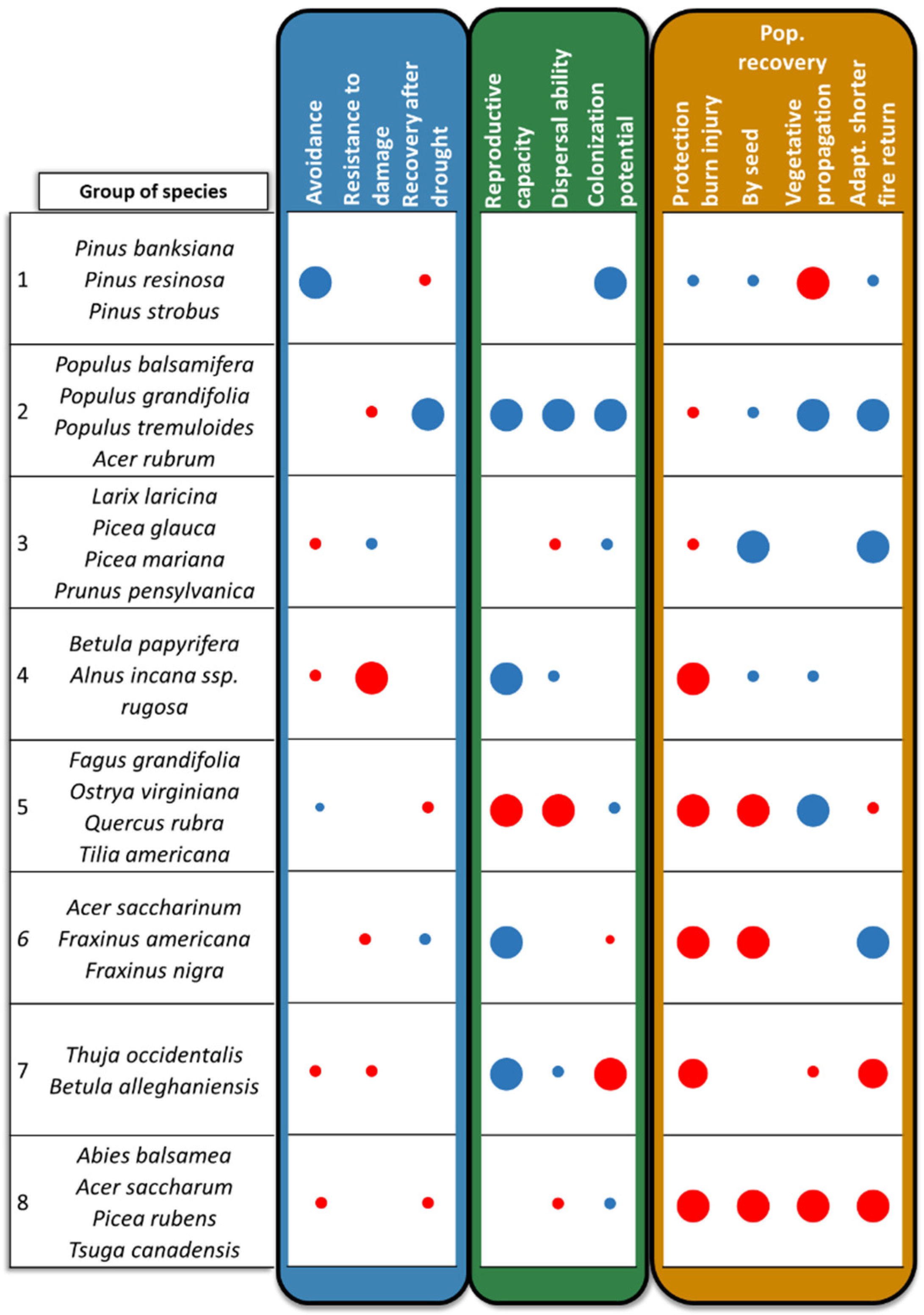
2.4. Defining Species Groups
2.5. Development of Integrated Indices of Drought Sensitivity and Comparison with Other Published Indices
3. Results
4. Discussion
4.1. Limitations of Trait-Based Indices
4.2. Species Sensitivity to Inform Adaptation Practices
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lemprière, T.C.; Bernier, P.; Carroll, A.; Flannigan, M.; Gilsenan, R.; McKenney, D.; Hogg, E.; Pedlar, J.; Blain, D. The Importance of Forest Sector Adaptation to Climate Change; Information Report NOR-X-416E; Canadian Forest Service, Natural Resources Canada: Québec, QC, Canada, 2008; 78p.
- Reyer, C.P.O.; Leuzinger, S.; Rammig, A.; Wolf, A.; Bartholomeus, R.P.; Bonfante, A.; de Lorenzi, F.; Dury, M.; Gloning, P.; Abou Jaoudé, R.; et al. A plant’s perspective of extremes: Terrestrial plant responses to changing climatic variability. Glob. Chang. Biol. 2013, 19, 75–89. [Google Scholar] [CrossRef] [PubMed]
- McKenney, D.W.; Pedlar, J.H.; Rood, R.B.; Price, D. Revisiting projected shifts in the climate envelopes of North American trees using updated general circulation models. Glob. Chang. Biol. 2011, 17, 2720–2730. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Flato, G.; Kirchmeier-Young, M.; Vincent, L.; Wan, H.; Wang, X.; Rong, R.; Fyfe, J.; Li, G.; Kharin, V. Changes in Temperature and Precipitation across Canada; Canada’s Changing Climate Report; Bush, E., Lemmen, D.S., Eds.; Government of Canada: Ottawa, ON, Canada, 2019; pp. 112–193. Available online: https://changingclimate.ca/CCCR2019/ (accessed on 5 September 2019).
- Bouchard, M.; Aquilué, N.; Périé, C.; Lambert, M.-C. Tree species persistence under warming conditions: A key driver of forest response to climate change. Ecol. Manag. 2019, 442, 96–104. [Google Scholar] [CrossRef]
- Boucher, D.; Boulanger, Y.; Aubin, I.; Bernier, P.Y.; Beaudoin, A.; Guindon, L.; Gauthier, S. Current and projected cumulative impacts of fire, drought, and insects on timber volumes across Canada. Ecol. Appl. 2018, 28, 1245–1259. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L. Temperate forest health in an era of emerging megadisturbance. Science 2015, 349, 823–826. [Google Scholar] [CrossRef]
- Edwards, J.; Pearce, C.; Ogden, A.; Williamson, T. Climate Change and Sustainable Forest Management in Canada: A Guidebook for Assessing Vulnerability and Mainstreaming Adaptation into Decision Making; Canadian Council of Forest Ministers: Ottawa, ON, Canada, 2015; p. 160. Available online: https://cfs.nrcan.gc.ca/publications/download-pdf/35956 (accessed on 5 September 2019).
- Swanston, C.W.; Janowiak, M.K.; Brandt, L.A.; Butler, P.R.; Handler, S.D.; Shannon, P.D.; Lewis, A.D.; Hall, K.; Fahey, R.T.; Scott, L. Forest Adaptation Resources: Climate Change Tools and Approaches for Land Managers; General Technical Reports; US Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2016; p. 161. [CrossRef]
- Nadeau, C.P.; Fuller, A.K.; Rosenblatt, D.L. Climate-smart management of biodiversity. Ecosphere 2015, 6, 1–17. [Google Scholar] [CrossRef]
- Rogers, B.M.; Jantz, P.; Goetz, S.J. Vulnerability of eastern US tree species to climate change. Glob. Chang. Biol. 2017, 23, 3302–3320. [Google Scholar] [CrossRef] [PubMed]
- Zolkos, S.G.; Jantz, P.; Cormier, T.; Iverson, L.R.; McKenney, D.W.; Goetz, S.J. Projected Tree Species Redistribution Under Climate Change: Implications for Ecosystem Vulnerability Across Protected Areas in the Eastern United States. Ecosystems 2015, 18, 202–220. [Google Scholar] [CrossRef]
- Aubin, I.; Munson, A.D.; Cardou, F.; Burton, P.J.; Isabel, N.; Pedlar, J.H.; Paquette, A.; Taylor, A.R.; Delagrange, S.; Kebli, H.; et al. Traits to stay, traits to move: A review of functional traits to assess sensitivity and adaptive capacity of temperate and boreal trees to climate change. Environ. Rev. 2016, 24, 164–186. [Google Scholar] [CrossRef]
- Stahl, U.; Reu, B.; Wirth, C. Predicting species’range limits from functional traits for the tree flora of North America. Proc. Natl. Acad. Sci. USA 2014, 111, 13739–13744. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.C.; Bocedi, G.; Hendry, A.P.; Mihoub, J.B.; Pe’er, G.; Singer, A.; Bridle, J.R.; Crozier, L.G.; De Meester, L.; Godsoe, W.; et al. Improving the forecast for biodiversity under climate change. Science 2016, 353, aad8466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glick, P.; Stein, B.; Edelson, N. Scanning the Conservation Horizon: A Guide to Climate Change Vulnerability Assessment; National Wildlife Federation: Washington, DC, USA, 2011; p. 168. Available online: https://www.fs.usda.gov/treesearch/pubs/37406 (accessed on 5 September 2019).
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.L. Spatial and temporal variation in plant hydraulic traits and their relevance for climate change impacts on vegetation. New Phytol. 2015, 205, 1008–1014. [Google Scholar] [CrossRef] [PubMed]
- Gilman, S.E.; Urban, M.C.; Tewksbury, J.; Gilchrist, G.W.; Holt, R.D. A framework for community interactions under climate change. Trends Ecol. Evol. 2010, 25, 325–331. [Google Scholar] [CrossRef]
- Helmuth, B.; Kingsolver, J.G.; Carrington, E. BIOPHYSICS, PHYSIOLOGICAL ECOLOGY, AND CLIMATE CHANGE: Does Mechanism Matter? Annu. Rev. Physiol. 2005, 67, 177–201. [Google Scholar] [CrossRef]
- Bussotti, F.; Pollastrini, M.; Holland, V.; Brüggemann, W. Functional traits and adaptive capacity of European forests to climate change. Environ. Exp. Bot. 2015, 111, 91–113. [Google Scholar] [CrossRef]
- Alberto, F.J.; Aitken, S.N.; Alía, R.; González-Martínez, S.C.; Hänninen, H.; Kremer, A.; Lefèvre, F.; Lenormand, T.; Yeaman, S.; Whetten, R.; et al. Potential for evolutionary responses to climate change—Evidence from tree populations. Glob. Chang. Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef] [Green Version]
- Lamhamedi, M.S.; Bernier, P.Y. Ecophysiology and field performance of black spruce (Picea mariana): A review. Ann. Sci. 1994, 51, 529–551. [Google Scholar] [CrossRef] [Green Version]
- Craver, C.; Tabery, J. Mechanisms in Science. In The Stanford Encyclopedia of Philosophy; Zalta, E.N., Ed.; Metaphysics Research Lab, Stanford University: Stanford, CA, USA, 2019; Available online: https://plato.stanford.edu/entries/science-mechanisms/ (accessed on 7 September 2019).
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Shipley, B. Comparative Plant Ecology as a Tool for Integrating Across Scales. Ann. Bot. 2007, 99, 965–966. [Google Scholar] [CrossRef]
- Willis, S.G.; Foden, W.; Baker, D.J.; Belle, E.; Burgess, N.D.; Carr, J.A.; Doswald, N.; Garcia, R.A.; Hartley, A.; Hof, C.; et al. Integrating climate change vulnerability assessments from species distribution models and trait-based approaches. Biol. Conserv. 2015, 190, 167–178. [Google Scholar] [CrossRef]
- Angert, A.L.; Crozier, L.G.; Rissler, L.J.; Gilman, S.E.; Tewksbury, J.J.; Chunco, A.J. Do species’traits predict recent shifts at expanding range edges? Ecol. Lett. 2011, 14, 677–689. [Google Scholar] [CrossRef]
- Pausas, J.G.; Verdú, M. Plant Persistence Traits in Fire-Prone Ecosystems of the Mediterranean Basin: A Phylogenetic Approach. Oikos 2005, 109, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, A.F.A.; Anderegg, W.R.L.; Paine, C.E.T.; Hoffmann, W.A.; Kartzinel, T.; Rabin, S.S.; Sheil, D.; Franco, A.C.; Pacala, S.W. Convergence of bark investment according to fire and climate structures ecosystem vulnerability to future change. Ecol. Lett. 2017, 20, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Garnier, E.; Navas, M.-L.; Grigulis, K. Plant Functional Diversity: Organism Traits, Community Structure, and Ecosystem Properties; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Lavorel, S.; McIntyre, S.; Landsberg, J.; Forbes, T.D.A. Plant functional classifications: From general groups to specific groups based on response to disturbance. Trends Ecol. Evol. 1997, 12, 474–478. [Google Scholar] [CrossRef]
- Foden, W.B.; Butchart, S.H.M.; Stuart, S.N.; Vié, J.-C.; Akçakaya, H.R.; Angulo, A.; DeVantier, L.M.; Gutsche, A.; Turak, E.; Cao, L.; et al. Identifying the World’s Most Climate Change Vulnerable Species: A Systematic Trait-Based Assessment of all Birds, Amphibians and Corals. PLoS ONE 2013, 8, e65427. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought, and waterlogging of temperate northern hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Butt, N.; Gallagher, R. Using species traits to guide conservation actions under climate change. Clim. Chang. 2018, 151, 317–332. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef] [PubMed]
- Aubin, I.; Cardou, F.; Boisvert-Marsh, L.; Garnier, E.; Strukelj, M.; Munson, A.D. Managing data locally to answer questions globally: The role of collaborative science in ecology. J. Veg. Sci. 2020, 31, 509–517. [Google Scholar] [CrossRef]
- Hicks, C.C.; Fitzsimmons, C.; Polunin, N.V.C. Interdisciplinarity in the environmental sciences: Barriers and frontiers. Env. Conserv. 2010, 37, 464–477. [Google Scholar] [CrossRef]
- Beaudoin, A.; Bernier, P.Y.; Guindon, L.; Villemaire, P.; Guo, X.J.; Stinson, G.; Bergeron, T.; Magnussen, S.; Hall, R.J. Mapping attributes of Canada’s forests at moderate resolution through kNN and MODIS imagery. Can. J. Res. 2014, 44, 521–532. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.S.; Iverson, L.; Woodall, C.W.; Allen, C.D.; Bell, D.M.; Bragg, D.C.; D'Amato, A.W.; Davis, F.W.; Hersh, M.H.; Ibanez, I.; et al. The impacts of increasing drought on forest dynamics, structure, and biodiversity in the United States. Glob. Chang. Biol. 2016, 22, 2329–2352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.P.; OsÓRio, M.L.; Carvalho, I.; Faria, T.; Pinheiro, C. How Plants Cope with Water Stress in the Field? Photosynthesis and Growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, H.; Ziegler, W.; Trumbore, S. Lethal drought leads to reduction in nonstructural carbohydrates in Norway spruce tree roots but not in the canopy. Funct. Ecol. 2013, 27, 413–427. [Google Scholar] [CrossRef]
- Meinzer, F.C.; Johnson, D.M.; Lachenbruch, B.; McCulloh, K.A.; Woodruff, D.R. Xylem hydraulic safety margins in woody plants: Coordination of stomatal control of xylem tension with hydraulic capacitance. Funct. Ecol. 2009, 23, 922–930. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Klein, T.; Jansen, S.; Choat, B.; Sack, L. The correlations and sequence of plant stomatal, hydraulic, and wilting responses to drought. Proc. Natl. Acad. Sci. USA 2016, 113, 13098. [Google Scholar] [CrossRef] [Green Version]
- Brodribb, T.J.; Holbrook, N.M. Stomatal protection against hydraulic failure: A comparison of coexisting ferns and angiosperms. New Phytol. 2004, 162, 663–670. [Google Scholar] [CrossRef]
- Martin-StPaul, N.; Delzon, S.; Cochard, H. Plant resistance to drought depends on timely stomatal closure. Ecol. Lett. 2017, 20, 1437–1447. [Google Scholar] [CrossRef]
- Johnson, D.M.; McCulloh, K.A.; Woodruff, D.R.; Meinzer, F.C. Hydraulic safety margins and embolism reversal in stems and leaves: Why are conifers and angiosperms so different? Plant Sci. 2012, 195, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Yakir, D.; Buchmann, N.; Grünzweig, J.M. Towards an advanced assessment of the hydrological vulnerability of forests to climate change-induced drought. New Phytol. 2014, 201, 712–716. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M.J.; Leuzinger, S.; Philipson, C.D.; Tay, J.; Hector, A. Drought survival of tropical tree seedlings enhanced by non-structural carbohydrate levels. Nat. Clim. Chang. 2014, 4, 710–714. [Google Scholar] [CrossRef]
- Dietze, M.C.; Sala, A.; Carbone, M.S.; Czimczik, C.I.; Mantooth, J.A.; Richardson, A.D.; Vargas, R. Nonstructural Carbon in Woody Plants. Annu. Rev. Plant Biol. 2014, 65, 667–687. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.G.; Beerling, D.J.; Breshears, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef]
- Tague, C.L.; McDowell, N.G.; Allen, C.D. An Integrated Model of Environmental Effects on Growth, Carbohydrate Balance, and Mortality of Pinus ponderosa Forests in the Southern Rocky Mountains. PLoS ONE 2013, 8, e80286. [Google Scholar] [CrossRef]
- Périé, C.; de Blois, S. Dominant forest tree species are potentially vulnerable to climate change over large portions of their range even at high latitudes. PeerJ 2016, 4, e2218. [Google Scholar] [CrossRef] [Green Version]
- Aubin, I.; Boisvert-Marsh, L.; Kebli, H.; McKenney, D.; Pedlar, J.; Lawrence, K.; Hogg, E.H.; Boulanger, Y.; Gauthier, S.; Ste-Marie, C. Tree vulnerability to climate change: Improving exposure-based assessments using traits as indicators of sensitivity. Ecosphere 2018, 9, e02108. [Google Scholar] [CrossRef]
- Boisvert-Marsh, L.; Périé, C.; de Blois, S. Shifting with climate? Evidence for recent changes in tree species distribution at high latitudes. Ecosphere 2014, 5, 1–33. [Google Scholar] [CrossRef]
- Clark, J.S. Why trees migrate so fast: Confronting theory with dispersal biology and the Paleorecord. Am. Nat. 1998, 152, 204–224. [Google Scholar] [CrossRef]
- Hampe, A. Plants on the move: The role of seed dispersal and initial population establishment for climate-driven range expansions. Acta Oecol. 2011, 37, 666–673. [Google Scholar] [CrossRef]
- Nathan, R.; Horvitz, N.; He, Y.; Kuparinen, A.; Schurr, F.M.; Katul, G.G. Spread of North American wind-dispersed trees in future environments. Ecol. Lett. 2011, 14, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Broennimann, O.; Thuiller, W.; Hughes, G.; Midgley, G.F.; Alkemade, J.M.R.; Guisan, A. Do geographic distribution, niche property and life form explain plants’vulnerability to global change? Glob. Chang. Biol. 2006, 12, 1079–1093. [Google Scholar] [CrossRef]
- Corlett, R.T.; Westcott, D.A. Will plant movements keep up with climate change? Trends Ecol. Evol. 2013, 28, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Estrada, A.; Morales-Castilla, I.; Caplat, P.; Early, R. Usefulness of Species Traits in Predicting Range Shifts. Trends Ecol. Evol. 2016, 31, 190–203. [Google Scholar] [CrossRef] [Green Version]
- Godoy, O.; Valladares, F.; Castro-Díez, P. The relative importance for plant invasiveness of trait means, and their plasticity and integration in a multivariate framework. New Phytol. 2012, 195, 912–922. [Google Scholar] [CrossRef] [Green Version]
- Pannell, J.R.; Barrett, S.C.H. Baker’s law revisited: Reproductive assurance in a metapopulation. Evolution 1998, 52, 657–668. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Boulanger, Y.; Gauthier, S.; Burton, P.J. A refinement of models projecting future Canadian fire regimes using homogeneous fire regime zones. Can. J. Res. 2014, 44, 365–376. [Google Scholar] [CrossRef]
- Girardin, M.P.; Ali, A.A.; Carcaillet, C.; Gauthier, S.; Hély, C.; Le Goff, H.; Terrier, A.; Bergeron, Y. Fire in managed forests of eastern Canada: Risks and options. Ecol. Manag. 2013, 294, 238–249. [Google Scholar] [CrossRef]
- van Mantgem, P.; Schwartz, M. Bark heat resistance of small trees in Californian mixed conifer forests: Testing some model assumptions. Ecol. Manag. 2003, 178, 341–352. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E.; Schwilk, D.W. Flammability as an ecological and evolutionary driver. J. Ecol. 2017, 105, 289–297. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Buma, B.; Brown, C.D.; Donato, D.C.; Fontaine, J.B.; Johnstone, J.F. The Impacts of Changing Disturbance Regimes on Serotinous Plant Populations and Communities. BioScience 2013, 63, 866–876. [Google Scholar] [CrossRef]
- Greene, D.F.; Zasada, J.C.; Sirois, L.; Kneeshaw, D.; Morin, H.; Charron, I.; Simard, M.J. A review of the regeneration dynamics of North American boreal forest tree species. Can. J. Res. 1999, 29, 824–839. [Google Scholar] [CrossRef]
- Zasada, J.C.; Norum, R.A.; Veldhuizen, R.M.V.; Teutsch, C.E. Artificial regeneration of trees and tall shrubs in experimentally burned upland black spruce/feather moss stands in Alaska. Can. J. Res. 1983, 13, 903–913. [Google Scholar] [CrossRef]
- Clarke, P.J.; Lawes, M.J.; Midgley, J.J.; Lamont, B.B.; Ojeda, F.; Burrows, G.E.; Enright, N.J.; Knox, K.J.E. Resprouting as a key functional trait: How buds, protection and resources drive persistence after fire. New Phytol. 2013, 197, 19–35. [Google Scholar] [CrossRef] [Green Version]
- Bond, W.J.; Midgley, J.J. Ecology of sprouting in woody plants: The persistence niche. Trends Ecol. Evol. 2001, 16, 45–51. [Google Scholar] [CrossRef]
- Keeley, J.E.; Keeley, M.B.; Bond, W.J. Stem demography and post-fire recruitment of a resprouting serotinous conifer. J. Veg. Sci. 1999, 10, 69–76. [Google Scholar] [CrossRef]
- Boulet, B.; Huot, M. Le Guide Sylvicole Du Québec: Les Fondements Biologiques De La Sylviculture; Publications du Québec: Québec, QC, Canada, 2013; Volume 1, p. 1044. [Google Scholar]
- USDA NRCS. The PLANTS Database; National Plant Data Team: Greensboro, NC, USA, 2009. Available online: http://plants.usda.gov (accessed on 19 September 2019).
- Hightshoe, G.L. Native Trees, Shrubs, and Vines for Urban and Rural America: A Planting Design Manual for Environmental Designers; Van Nostrand Reinhold: New York, NY, USA, 1988. [Google Scholar]
- Ottawa Forests and Greenspace Committee. OFGAC Native Trees and Shrubs Database. Available online: https://ofnc.ca/programs/fletcher-wildlife-garden/flora-and-fauna-at-the-fwg/trees-and-shrubs-for-wildlife-gardens (accessed on 12 September 2019).
- Matthews, S.N.; Iverson, L.R.; Prasad, A.M.; Peters, M.P.; Rodewald, P.G. Modifying climate change habitat models using tree species-specific assessments of model uncertainty and life history-factors. Ecol. Manag. 2011, 262, 1460–1472. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Kane, J.M.; Anderegg, L.D.L. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 2013, 3, 30–36. [Google Scholar] [CrossRef]
- Adams, H.D.; Zeppel, M.J.; Anderegg, W.R.; Hartmann, H.; Landhäusser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.L.; Flint, A.; Huang, C.-y.; Flint, L.; Berry, J.A.; Davis, F.W.; Sperry, J.S.; Field, C.B. Tree mortality predicted from drought-induced vascular damage. Nat. Geosci. 2015, 8, 367–371. [Google Scholar] [CrossRef] [Green Version]
- Wesołowski, T.; Rowiński, P.; Maziarz, M. Interannual variation in tree seed production in a primeval temperate forest: Does masting prevail? Eur. J. Res. 2015, 134, 99–112. [Google Scholar] [CrossRef] [Green Version]
- Morin, X.; Augspurger, C.; Chuine, I. Process-based modeling of species’ distributions: What limits temperate tree species’ range boundaries? Ecology 2007, 88, 2280–2291. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, M.F.; Bergeron, Y.; Lalonde, D.; Mauffette, Y. The potential effects of sexual reproduction and seedling recruitment on the maintenance of red maple (Acer rubrum L.) populations at the northern limit of the species range. J. Biogeogr. 2002, 29, 365–373. [Google Scholar] [CrossRef]
- Kattge, J.; Bönisch, G.; Díaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Tautenhahn, S.; Werner, G.D.A.; Aakala, T.; Abedi, M.; et al. TRY plant trait database—Enhanced coverage and open access. Glob. Chang. Biol. 2020, 26, 119–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urli, M.; Porté, A.J.; Cochard, H.; Guengant, Y.; Burlett, R.; Delzon, S. Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees. Tree Physiol. 2013, 33, 672–683. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [Green Version]
- Flannigan, M.D.; Krawchuk, M.A.; de Groot, W.J.; Wotton, B.M.; Gowman, L.M. Implications of changing climate for global wildland fire. Int. J. Wildland Fire 2009, 18, 483–507. [Google Scholar] [CrossRef]
- Leuschner, C.; Meier, I.C. The ecology of Central European tree species: Trait spectra, functional trade-offs, and ecological classification of adult trees. Perspect. Plant Ecol. Evol. Syst. 2018, 33, 89–103. [Google Scholar] [CrossRef]
- Canadell, J.; Jackson, R.B.; Ehleringer, J.B.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. Maximum rooting depth of vegetation types at the global scale. Oecologia 1996, 108, 583–595. [Google Scholar] [CrossRef]
- Farrar, J.L. Trees in Canada; Fitzhenry & Whiteside Ltd. and Canadian Forest Service: Ottawa, ON, Canada, 1995.
- Minore, D. Comparative Autecological Characteristics of Northwestern Tree Species: A Literature Review. General Technical Reports PNW-GTR-087; Department of Agriculture, Forest Service, Pacific Northwest Forest Station: Portland, OR, USA, 1979; p. 72. [Google Scholar]
- Stone, E.L.; Kalisz, P.J. On the maximum extent of tree roots. Ecol. Manag. 1991, 46, 59–102. [Google Scholar] [CrossRef]
- Bullock, J.M.; Mallada González, L.; Tamme, R.; Götzenberger, L.; White, S.M.; Pärtel, M.; Hooftman, D.A.P. A synthesis of empirical plant dispersal kernels. J. Ecol. 2017, 105, 6–19. [Google Scholar] [CrossRef]
- Vittoz, P.; Engler, R. Seed dispersal distances: A typology based on dispersal modes and plant traits. Bot. Helv. 2007, 117, 109–124. [Google Scholar] [CrossRef] [Green Version]
- Tamme, R.; Götzenberger, L.; Zobel, M.; Bullock, J.M.; Hooftman, D.A.P.; Kaasik, A.; Pärtel, M. Predicting species’ maximum dispersal distances from simple plant traits. Ecology 2014, 95, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, S.; Bernier, P.; Burton, P.J.; Edwards, J.; Isaac, K.; Isabel, N.; Jayen, K.; Le Goff, H.; Nelson, E.A. Climate change vulnerability and adaptation in the managed Canadian boreal forest. Environ. Rev. 2014, 22, 256–285. [Google Scholar] [CrossRef]
- Messier, C.; Bauhus, J.; Doyon, F.; Maure, F.; Sousa-Silva, R.; Nolet, P.; Mina, M.; Aquilué, N.; Fortin, M.-J.; Puettmann, K. The functional complex network approach to foster forest resilience to global changes. For. Ecosyst. 2019, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Park, A.; Puettmann, K.; Wilson, E.; Messier, C.; Kames, S.; Dhar, A. Can Boreal and Temperate Forest Management be Adapted to the Uncertainties of 21st Century Climate Change? Crit. Rev. Plant Sci. 2014, 33, 251–285. [Google Scholar] [CrossRef]
- Hogg, E.H.; Brandt, J.P.; Kochtubajda, B. Growth and dieback of aspen forests in northwestern Alberta, Canada, in relation to climate and insects. Can. J. Res. 2002, 32, 823–832. [Google Scholar] [CrossRef]
- Wade, A.A.; Hand, B.K.; Kovach, R.P.; Luikart, G.; Whited, D.C.; Muhlfeld, C.C. Accounting for adaptive capacity and uncertainty in assessments of species’ climate-change vulnerability. Conserv. Biol. 2017, 31, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Belote, R.T.; Carroll, C.; Martinuzzi, S.; Michalak, J.; Williams, J.W.; Williamson, M.A.; Aplet, G.H. Assessing agreement among alternative climate change projections to inform conservation recommendations in the contiguous United States. Sci. Rep. 2018, 8, 9441. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.D.; Fahrig, L.; Henein, K.; Merriam, G. Connectivity is a vital element of landscape structure. Oikos 1993, 68, 571–573. [Google Scholar] [CrossRef] [Green Version]
- Fischer, A.P. Adapting and coping with climate change in temperate forests. Glob. Env. Chang. 2019, 54, 160–171. [Google Scholar] [CrossRef]
- Pedlar, J.H.; McKenney, D.W.; Aubin, I.; Beardmore, T.; Beaulieu, J.; Iverson, L.; O’Neill, G.A.; Winder, R.S.; Ste-Marie, C. Placing Forestry in the Assisted Migration Debate. BioScience 2012, 62, 835–842. [Google Scholar] [CrossRef]
- Nagel, L.M.; Palik, B.J.; Battaglia, M.A.; D’Amato, A.W.; Guldin, J.M.; Swanston, C.W.; Janowiak, M.K.; Powers, M.P.; Joyce, L.A.; Millar, C.I.; et al. Adaptive Silviculture for Climate Change: A National Experiment in Manager-Scientist Partnerships to Apply an Adaptation Framework. J. For. 2017, 115, 167–178. [Google Scholar] [CrossRef]
- Morelli, T.L.; Daly, C.; Dobrowski, S.Z.; Dulen, D.M.; Ebersole, J.L.; Jackson, S.T.; Lundquist, J.D.; Millar, C.I.; Maher, S.P.; Monahan, W.B.; et al. Managing Climate Change Refugia for Climate Adaptation. PLoS ONE 2016, 11, e0159909. [Google Scholar] [CrossRef] [Green Version]
- Prévost, M.; Raymond, P.; Lussier, J.-M. Regeneration dynamics after patch cutting and scarification in yellow birch—Conifer stands. Can. J. Res. 2010, 40, 357–369. [Google Scholar] [CrossRef]
- Brice, M.-H.; Vissault, S.; Vieira, W.; Gravel, D.; Legendre, P.; Fortin, M.-J. Moderate disturbances accelerate forest transition dynamics under climate change in the temperate–boreal ecotone of eastern North America. Glob. Chang. Biol. 2020, 26, 4418–4435. [Google Scholar] [CrossRef]
- Lafleur, B.; Paré, D.; Munson, A.D.; Bergeron, Y. Response of northeastern North American forests to climate change: Will soil conditions constrain tree species migration? Env. Rev. 2010, 18, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Boisvert-Marsh, L.; de Blois, S. Unravelling potential northward migration pathways for tree species under climate change. J. Biogeogr. 2020. in review. [Google Scholar]
- Lindenmayer, D.B.; Likens, G.E.; Krebs, C.J.; Hobbs, R.J. Improved probability of detection of ecological “surprises”. Proc. Natl. Acad. Sci. USA 2010, 107, 21957. [Google Scholar] [CrossRef] [Green Version]
- Janowiak, M.K.; Swanston, C.W.; Nagel, L.M.; Brandt, L.A.; Butler, P.R.; Handler, S.D.; Shannon, P.D.; Iverson, L.R.; Matthews, S.N.; Prasad, A.; et al. A Practical Approach for Translating Climate Change Adaptation Principles into Forest Management Actions. J. For. 2014, 112, 424–433. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.C. Climate-tracking species are not invasive. Nat. Clim. Chang. 2020, 10, 382–384. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drought Sensitivity—Stem and Population 1 | Drought Sensitivity—Stem Only 2 | Niinemets and Valladares (2006) [34] 3 | Matthews et al. (2011) [81] 4 | Boulet and Huot (2013) [77] 5 | USDA NRCS (2009) [78] 6 | Hightshoe (1988) [79] 7 | OFGAC Native Trees and Shrubs Database [80] 8 | |
|---|---|---|---|---|---|---|---|---|
| Drought sensitivity—stem and population 1 | 0.72 *** | 0.40 ** | 0.18 | 0.63 *** | −0.061 | 0.59 *** | 0.028 | |
| Drought sensitivity—stem only 2 | 0.61 *** | 0.35 * | 0.49 ** | 0.0034 | 0.26 | 0.30 | ||
| Niinemets and Valladares (2006) 3 | 0.34 * | 0.53 *** | 0.11 | 0.22 | 0.37 * | |||
| Matthews et al. (2011) 4 | 0.21 | 0.50 ** | 0.077 | 0.27 | ||||
| Boulet and Huot (2013) 5 | 0.019 | 0.65 *** | 0.17 | |||||
| USDA NRCS (2009) 6 | −0.13 | 0.19 | ||||||
| Hightshoe (1988) 7 | −0.27 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boisvert-Marsh, L.; Royer-Tardif, S.; Nolet, P.; Doyon, F.; Aubin, I. Using a Trait-Based Approach to Compare Tree Species Sensitivity to Climate Change Stressors in Eastern Canada and Inform Adaptation Practices. Forests 2020, 11, 989. https://doi.org/10.3390/f11090989
Boisvert-Marsh L, Royer-Tardif S, Nolet P, Doyon F, Aubin I. Using a Trait-Based Approach to Compare Tree Species Sensitivity to Climate Change Stressors in Eastern Canada and Inform Adaptation Practices. Forests. 2020; 11(9):989. https://doi.org/10.3390/f11090989
Chicago/Turabian StyleBoisvert-Marsh, Laura, Samuel Royer-Tardif, Philippe Nolet, Frédérik Doyon, and Isabelle Aubin. 2020. "Using a Trait-Based Approach to Compare Tree Species Sensitivity to Climate Change Stressors in Eastern Canada and Inform Adaptation Practices" Forests 11, no. 9: 989. https://doi.org/10.3390/f11090989
APA StyleBoisvert-Marsh, L., Royer-Tardif, S., Nolet, P., Doyon, F., & Aubin, I. (2020). Using a Trait-Based Approach to Compare Tree Species Sensitivity to Climate Change Stressors in Eastern Canada and Inform Adaptation Practices. Forests, 11(9), 989. https://doi.org/10.3390/f11090989