
Dothistroma septosporum Not Detected in Pinus sylvestris Seed Trees from Investigated Stands in Southern Poland


 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
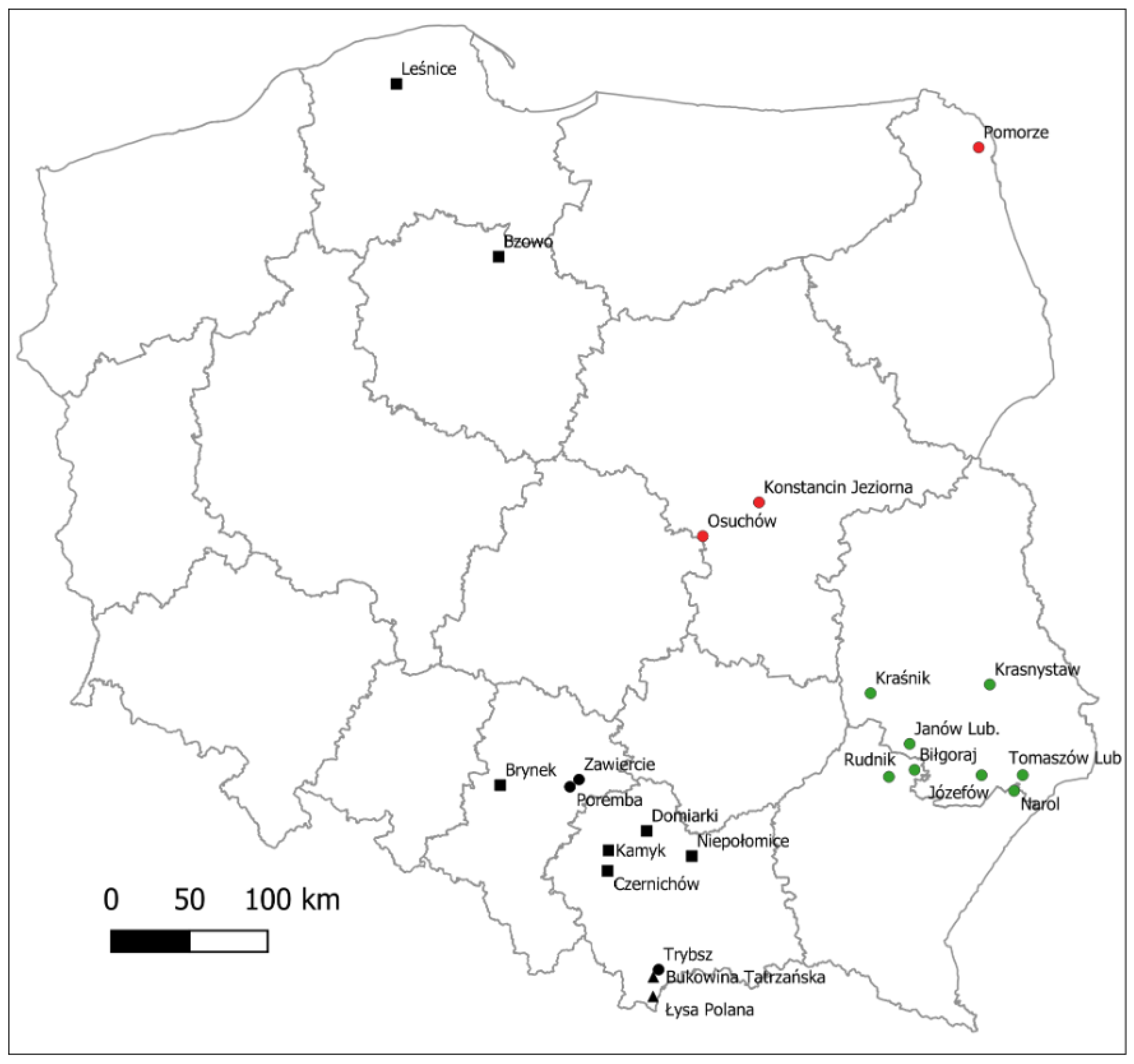
2. Materials and Methods
2.1. Preliminary Assessment
2.2. Main Study to Detect Pathogens in Seed Tree Needles
2.2.1. Sampling of Plant Material
2.2.2. Preparation of Fungal Positive Control
2.3. Genomic DNA Isolation
2.4. Molecular Analysis of the Sampled Material
2.5. Sequencing of the PCR Amplicons
3. Results
4. Discussion
4.1. Emerging Pine Needle Pathogens
4.2. Accuracy of the Molecular Methods in D. septosporum Detection
4.3. Pathway of Spread of D. septosporum in Europe
4.4. European Approach Problem Solving
4.5. Influence of Climatic Changes on Dothistroma Distribution
5. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raitelaitytė, K.; Markovskaja, S.; Paulauskas, A.; Hsiang, T.; Oszako, T. First molecular detection of Lecanosticta acicola from Poland on Pinus mugo. For. Pathol. 2020, 50, e12589. [Google Scholar] [CrossRef]
- Drenkhan, R.; Ganley, B.; Martín-García, J.; Vahalík, P.; Adamson, K.; Adamcíková, K.; Ahumada, R.; Blank, L.; Bragança, H.; Capretti, P.; et al. Global geographic distribution and host range of Fusarium circinatum, the causal agent of Pine Pitch Canker. Forests 2020, 11, 724. [Google Scholar] [CrossRef]
- Elvira-Recuenco, M.; Cacciola, S.O.; Sanz-Ros, A.V.; Garbelotto, M.; Aguayo, J.; Solla, A.; Mullett, M.; Drenkhan, T.; Oskay, F.; Kaya, A.G.A.; et al. Potential interactions between invasive Fusarium circinatum and other pine pathogens in Europe. Forests 2020, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Davydenko, K.; Nowakowska, J.A.; Kaluski, T.; Gawlak, M.; Sadowska, K.; García, J.M.; Diez, J.J.; Okorski, A.; Oszako, T. A Comparative study of the pathogenicity of Fusarium circinatum and other fusarium species in Polish provenances of P. sylvestris L. Forests 2018, 9, 560. [Google Scholar] [CrossRef] [Green Version]
- Barnes, I.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Multigene phylogenies reveal that red band needle blight of Pinus is caused by two distinct species of Dothistroma, D. septosporum and D. pini. Stud. Mycol. 2004, 50, 551–565. [Google Scholar]
- Drenkhan, R.; Tomešová-Haataja, V.; Fraser, S.; Bradshaw, R.E.; Vahalík, P.; Mullett, M.S.; Martín-García, J.; Bulman, L.S.; Wingfield, M.J.; Kirisits, T.; et al. Global geographic distribution and host range of Dothistroma species: A comprehensive review. For. Pathol. 2016, 46, 408–442. [Google Scholar] [CrossRef]
- Watt, M.S.; Kriticos, D.J.; Alcaraz, S.; Brown, A.V.; Leriche, A. The hosts and potential geographic range of Dothistroma needle blight. For. Ecol. Manag. 2009, 257, 1505–1519. [Google Scholar] [CrossRef]
- Barnes, I.; Kirisits, T.; Akulov, A.; Chhetri, D.B.; Wingfield, B.D.; Bulgakov, T.S.; Wingfield, M.J. New host and country records of the Dothistroma needle blight pathogens from Europe and Asia. For. Pathol. 2008, 38, 178–195. [Google Scholar] [CrossRef]
- Funk, A.; Parker, A.K. Scirrhia pini n. sp., the perfect state of Dothistroma pini Hulbary. Can. J. Bot. 1966, 44, 1171–1176. [Google Scholar] [CrossRef]
- Doroguine, M.D.M. A Cryptogamic disease of pines. Bull. Société Mycol. Fr. 1911, 27, 105–106. [Google Scholar]
- Morelet, M. Une maladie des pins, nouvelle pour la France, due à Scirrhia pini Funk et Parker et à son stade conidien Dothistroma pini Hulbary. Bull. Mens. Société Linnéenne Lyon 1967, 36, 361–367. [Google Scholar] [CrossRef]
- Bradshaw, R.E. Dothistroma (Red-Band) needle blight of pines and the dothistromin toxin: A review. For. Pathol. 2004, 34, 163–185. [Google Scholar] [CrossRef]
- Gibson, I.A.S. Impact and control of Dothistroma blight of pines. Eur. J. For. Pathol. 1974, 4, 89–100. [Google Scholar] [CrossRef]
- Hanso, M.; Drenkhan, R. Diplodia pinea is a new pathogen on Austrian pine (Pinus nigra) in Estonia. Plant. Pathol. 2009, 58, 797. [Google Scholar] [CrossRef]
- Woods, A.J. Species diversity and forest health in northwest British Columbia. For. Chron. 2003, 79, 892–897. [Google Scholar] [CrossRef]
- Ghelardini, L.; Aglietti, C.; Loria, F.; Cerboneschi, M.; Gionni, A.; Goti, E.; Maresi, G.; Moricca, S.; Marchi, G. Dothistroma needle blight in protected pine forests in Italy. Manag. Biol. Invasions 2020, 11, 689–702. [Google Scholar] [CrossRef]
- Fabre, B.; Ioos, R.; Piou, D.; Marçais, B. Is the emergence of Dothistroma needle blight of pine in France caused by the cryptic species Dothistroma pini? Phytopathology 2012, 102, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullett, M.S.; Brown, A.V.; Barnes, I. Population structure and reproductive mode of Dothistroma septosporum in the Brittany peninsula of France. Eur. J. Plant Pathol. 2015, 143, 261–275. [Google Scholar] [CrossRef]
- Murray, J.S.; Batko, S. Dothistroma pini Hulbary: A new disease on pine in Britain. Forestry 1962, 34, 57–65. [Google Scholar] [CrossRef]
- Karadžić, D. Study on Bioecology of Fungus Dothistroma pini—Cause Forestry, of Needle Defoliation of Austrian Pine. Ph.D. Thesis, University of Belgrade, Belgrade, Serbia, 1986; p. 328. (In Serbian). [Google Scholar]
- Kailidis, D.S.; Markalas, S. The fungus Dothistroma pini and the dasika, lichen Evernia prunastri on Pinus brutia at the “Valley of Death”. For. Chron. 1981, 24, 257–259. [Google Scholar]
- Maček, J. Scirrhia pini, the pathogen of a new disease of pine in Slovenia. Gozdarski Vestn. 1975, 33, 9–11. [Google Scholar]
- Fernández, F.J. Dothistroma Needle Blight in P. radiata; Ministerio de Agricultura, Alimentación y Medio Ambiente: Madrid, Spain, 1975. (In Spanish)
- Magnani, G. Presence of Dothistroma pini on Pinus radiata. Cellul. Carta 1977, 28, 37–42. [Google Scholar]
- Butin, H.; Richter, J.X. Dothistroma needle blight: A new pine disease in the German Federal Republic. Nachr. Dtsch. Pflanzenschutzd. 1983, 35, 129–131. [Google Scholar]
- Szabó, I. Occurrence of Dothistroma septospora in Austrian pine plantation. Erdészeti Gazdászati Lapok 1997, 132, 44–45. [Google Scholar]
- Kunca, A.; Foffová, E. The risk of infestation of Pinus nigra stands in quarantine pathogen Dothistroma septospora (Dorog.) Morelet. In Proceedings of the Current Problems in Forest Protection 2000; Zvolen, Lesnícky Výskumný Ústav; Slovak Forest Research Institute (LVU): Zvolen, Slovakia, 2000; pp. 136–139. (In Slovak) [Google Scholar]
- Jankovský, L.; Palovčíková, D.B.M. Picea pungens engelm—A new host of Dothistroma needle blight Mycosphaerella pini E. rostrup detected in the Czech Republic. J. For. Sci. 2009, 50, 235–236. [Google Scholar] [CrossRef] [Green Version]
- Hanso, M.; Drenkhan, R. First observations of Mycosphaerella pini in Estonia. Plant Pathol. 2008, 57, 1177. [Google Scholar] [CrossRef]
- Davydenko, K.; Baturkyn, D.; Hnoievyi, I.; Shcherbak, O. New data on host range and geographical distribution of Dothistroma needle blight in Ukraine. Environ. Sci. Proc. 2020, 3, 89. [Google Scholar] [CrossRef]
- Musolin, D.; Bulgakov, T.; Selikhovkkin, V.A. Dothistroma needle blight in Russia and adjacent countries. In Proceedings of the Forests of Russia: Politics, Industry, Science, Education, Saint-Petersburg, Russia, 24–26 May 2017. (In Russian). [Google Scholar]
- Fay, D.L. Dotistromosis—A new dangerous disease of the Crimean pine in the south of Russia. Angew. Chem. Int. Ed. 1967, 6, 951–952. [Google Scholar]
- Millberg, H.; Hopkins, A.J.M.; Boberg, J.; Davydenko, K.; Stenlid, J. Disease development of Dothistroma needle blight in seedlings of Pinus sylvestris and Pinus contorta under Nordic conditions. For. Pathol. 2016, 46, 515–521. [Google Scholar] [CrossRef]
- Kowalski, T.; Jankowiak, R. First record of Dothistroma septospora (Dorog.) morelet in Poland: A contribution to the symptomatology and epidemiology. Phytopathol. Pol. 1998, 16, 15–29. [Google Scholar]
- Boroń, P.; Lenart-Boroń, A.; Mullett, M. The distribution of Dothistroma septosporum and its mating types in Poland. For. Pathol. 2016, 46, 489–496. [Google Scholar] [CrossRef]
- Mullett, M.S.; Drenkhan, R.; Adamson, K.; Boroń, P.; Lenart-Boroń, A.; Barnes, I.; Tomšovský, M.; Jánošíková, Z.; Adamčíková, K.; Ondrušková, E.; et al. Worldwide genetic structure elucidates the Eurasian origin and invasion pathways of Dothistroma septosporum, causal agent of Dothistroma needle blight. J. Fungi 2021, 7, 111. [Google Scholar] [CrossRef]
- Ioos, R.; Fabre, B.; Saurat, C.; Fourrier, Ć.; Frey, P.; Marçais, B. Development, comparison, and validation of real-time and conventional PCR tools for the detection of the fungal pathogens causing brown spot and red band needle blights of pine. Phytopathology 2010, 100, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karadzic, D.; Milijasevic, T. The most important parasitic and saprophytic fungi in Austrian pine and Scots pine plantations in Serbia. Glas. Sumar. Fak. 2008, 97, 147–170. [Google Scholar] [CrossRef]
- van der Nest, A.; Wingfield, M.J.; Janoušek, J.; Barnes, I. Lecanosticta acicola: A growing threat to expanding global pine forests and plantations. Mol. Plant Pathol. 2019, 20, 1327–1364. [Google Scholar] [CrossRef] [Green Version]
- The State Forests Information Centre. Forests in Poland 2017; The State Forests Information Centre: Warszawa, Poland, 2017.
- Kowalski, T.; Zych, P. Fungi isolated from living symptomless shoots of Pinus nigra growing in different site conditions. Osterr. Pilzkd. 2002, 11, 107–116. [Google Scholar]
- Frank, A.; Saldierna Guzmán, J.; Shay, J. Transmission of bacterial endophytes. Microorganisms 2017, 5, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truyens, S.; Weyens, N.; Cuypers, A.; Vangronsveld, J. Bacterial seed endophytes: Genera, vertical transmission and interaction with plants. Environ. Microbiol. Rep. 2015, 7, 40–50. [Google Scholar] [CrossRef]
- Rajković, S.; Tabaković-Tošić, M.; Rakonjac, L.J.; Ratknić, M.; Veselinović, M. Fungicide in controlling powdery mildews. In Proceedings of the International Conference: Forestry in Achieving Millennium Goals; Slovak Forest Research Institute (LVU): Zvolen, Slovakia, 2009; pp. 399–405. [Google Scholar]
- Janoušek, J.; Wingfield, M.J.; Marmolejo Monsivais, J.G.; Jankovský, L.; Stauffer, C.; Konečný, A.; Barnes, I. Genetic analyses suggest separate introductions of the pine pathogen Lecanosticta acicola into Europe. Phytopathology 2016, 106, 1413–1425. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, S.; de Cates, C.; Hodgson, J.; Morley, N.J.; Sutton, B.C.; Gange, A.C. Vertical transmission of fungal endophytes is widespread in forbs. Ecol. Evol. 2014, 4, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- McDougal, R.; Stewart, A.; Bradshaw, R. Transformation of Cyclaneusma minus with Green Fluorescent Protein (GFP) to enable screening of fungi for biocontrol activity. Forests 2012, 3, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Flowers, J.; Nuckles, E.; Hartman, J.; Vaillancourt, L. Latent infection of Austrian and Scots pine tissues by Sphaeropsis sapinea. Plant Dis. 2001, 85, 1107–1112. [Google Scholar] [CrossRef] [Green Version]
- Duda, B.; Sierota, Z. Diseases caused by Gremmeniella Abietina (Lagerb.) Schlapfer−Bernhard and Cenangium ferruginosum Fr. Ex Fr. in Scots pine (Pinus sylvestris L.) stands in Poland. In Foliage, Shoot and Stem Diseases of Trees; Canadian Forest Service: Quebec City, QC, Canada, 1997; pp. 90–94. [Google Scholar]
- Drenkhan, R.; Hantula, J.; Vuorinen, M.; Jankovský, L.; Müller, M.M. Genetic diversity of Dothistroma septosporum in Estonia, Finland and Czech Republic. Eur. J. Plant Pathol. 2013, 136, 71–85. [Google Scholar] [CrossRef]
- Koziol, C.; Matras, J. The First Country Report on Forest Genetic Resources; Lasy Państowe: Warszawa, Poland, 2013.
- Matras, J. A Review of the seed orchard programme in Poland. In Proceedings of the In Seed Orchard Conference, Umea, Sweden, 26–28 September 2008; pp. 165–171. [Google Scholar]
- Nowakowska, J.A. Genetic Variation of Polish Scots Pine (Pinus sylvestris L.) Populations Assessed with DNA Polymorphism Markers; Prace Instytutu Badawczego Leśnictwa—Rozprawy i Monografie; IBL: Sękocin Stary, Poland, 2007; Volume 9, 112p. [Google Scholar]
- Wright, S. The interpretation of population structure by f-statistics with special regard to systems of mating. Evolution 1965, 19, 395–420. [Google Scholar] [CrossRef]
- Normes OEPP EPPO Standards. Lecanosticta acicola (Formerly Mycosphaerella dearnessii), Dothistroma septosporum (Formerly Mycosphaerella pini) and Dothistroma pini. Bull. OEPP/EPPO Bull. 2015, 45, 163–182. [Google Scholar] [CrossRef]
- Lee, S.K.; Lee, S.K.; Bae, H.; Seo, S.T.; Lee, J.K. Effects of water stress on the endophytic fungal communities of Pinus koraiensis needles infected by Cenangium ferruginosum. Mycobiology 2014, 42, 331–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowakowska, J.A. Application of DNA markers against illegal logging as a new tool for the forest guard service. Folia For. Pol. 2011, 53, 142–149. [Google Scholar]
- Barnes, I.; Wingfield, M.J.; Carbone, I.; Kirisits, T.; Wingfield, B.D. Population structure and diversity of an invasive pine needle pathogen reflects anthropogenic activity. Ecol. Evol. 2014, 4, 3642–3661. [Google Scholar] [CrossRef] [PubMed]
- Ioos, R.; Fourrier, C.; Iancu, G.; Gordon, T.R. Sensitive detection of Fusarium circinatum in pine seed by combining an enrichment procedure with a real-time polymerase chain reaction using dual-labeled probe chemistry. Phytopathology 2009, 99, 582–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlikowski, L.B.; Oszako, T.; Trzewik, A.; Orlikowska, T. Occurrence of Phytophthora ramorum and other Phytophthora species in nurseries, trade stands, forests and water. J. Plant Prot. Res. 2007, 47, 443–455. [Google Scholar]
- Eschen, R.; O’Hanlon, R.; Santini, A.; Vannini, A.; Roques, A.; Kirichenko, N.; Kenis, M. Safeguarding global plant health: The rise of sentinels. J. Pest. Sci. 2019, 92, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Morales-Rodríguez, C.; Anslan, S.; Auger-Rozenberg, M.A.; Augustin, S.; Baranchikov, Y.; Bellahirech, A.; Burokiene, D.; Čepukoit, D.; Çota, E.; Davydenko, K.; et al. Forewarned is forearmed: Harmonized approaches for early detection of potentially invasive pests and pathogens in sentinel plantings. NeoBiota 2019, 123, 95–123. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Plant Health. Scientific Opinion on the Risk to Plant Health Posed by Dothistroma septosporum (Dorog.) M. Morelet (Mycosphaerella pini E. Rostrup, Syn. Scirrhia pini) and Dothistroma pini Hulbary to the EU Territory with the Identification and Evaluation of Risk Reduction. EFSA J. 2013, 11, 3026. [Google Scholar]
- Rodriguez, R.J.; White, J.F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles: Tansley review. N. Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Walters, D.R.; Avrova, A.; Bingham, I.J.; Burnett, F.J.; Fountaine, J.; Havis, N.D.; Hoad, S.P.; Hughes, G.; Looseley, M.; Oxley, S.J.P.; et al. Control of foliar diseases in barley: Towards an integrated approach. Eur. J. Plant Pathol. 2012, 133, 33–73. [Google Scholar] [CrossRef]
- Ripple, W.J.; Wolf, C.; Newsome, T.M.; Gregg, J.W.; Lenton, T.M.; Palomo, I.; Eikelboom, J.A.J.; Law, B.E.; Huq, S.; Duffy, P.B.; et al. World scientists’ warning of a climate emergency 2021. BioScience 2021, 71, 894–898. [Google Scholar] [CrossRef]
- Nowakowska, J.A.; Hsiang, T.; Patynek, P.; Stereńczak, K.; Olejarski, I.; Oszako, T. Health assessment and genetic structure of monumental Norway spruce trees during a bark beetle (Ips typographus L.) outbreak in the Białowieża Forest District, Poland. Forests 2020, 11, 647. [Google Scholar] [CrossRef]
- Hanso, M.; Drenkhan, R. Kliimamuutuste visualiseerimise Lihtne viis seoste paremaks tajumiseks metsapatoloogias. For. Stud. 2013, 58, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Bulman, L.S.; Bradshaw, R.E.; Fraser, S.; Martín-García, J.; Barnes, I.; Musolin, D.L.; la Porta, N.; Woods, A.J.; Diez, J.J.; Koltay, A.; et al. A worldwide perspective on the management and control of Dothistroma needle blight. For. Pathol. 2016, 46, 472–488. [Google Scholar] [CrossRef] [Green Version]
- Linde, C.C.; Zhan, J.; McDonald, B.A. Population structure of Mycosphaerella graminicola: From lesions to continents. Phytopathology 2002, 92, 946–955. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Species | Gene | Primer Name | Primer Sequence | Expected Fragment Length (bp) | Ref. |
|---|---|---|---|---|---|
| Dothistroma septosporum | β-tubulin | DStub2-F | 5′-CGAACATGGACTGAGCAAAAC-3′ | 231 | [37,55] |
| DStub2-R | 5′-GCACGGCTCTTTCAAATGAC-3′ | ||||
| D. pini | EF1-α | DPtef-F | 5′-ATTTTTCGCTGCTCGTCACT-3′ | 193 | [37,55] |
| DPtef-R | 5′-CAATGTGAGATGTTCGTCGTG-3′ | ||||
| Lecanosticta acicola | EF1-α | LAtef-F | 5′-GCAAATTTTCGCCGTTTATC-3′ | 237 | [37,55] |
| LAtef-F | 5′-TGTGTTCCAAGAGTGCTTGC-3′ | ||||
| Cenangium ferruginosum | ITS | CfF | 5′-GA*TCATTAC CAGAAGTGTCC-3′ | 477 | [56] |
| CfR | 5′-CCTAGGTGA GTTGGGGTTGC-3′ |
| Sample * | Host Species | Age of Tree | Dothistroma septosporum | D. pini | Lecanosticta Acicola | Cenangium ferruginosum |
|---|---|---|---|---|---|---|
| β-tubulin | EF1-α | EF1-α | ITS | |||
| 1 | P. sylvestris | ~20 | +/99% | - | - | - |
| 2 | P. sylvestris | ~80 | - | - | - | - |
| 3 | P. nigra | ~20 | - | - | - | - |
| 4 | P. sylvestris | ~80 | - | - | - | - |
| 5 | P. nigra | ~25 | - | - | - | - |
| 6 | P. nigra | ~25 | - | - | - | - |
| 7 | P. sylvestris | ~80 | - | - | - | - |
| 8 | P. nigra | ~25 | +/98% | +/99% | - | - |
| 9 | P. sylvestris | ~80 | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wartalska, P.; Oszako, T.; Bakier, S.; Belbahri, L.; Malewski, T.; Hsiang, T.; Popowska-Nowak, E.; Nowakowska, J. Dothistroma septosporum Not Detected in Pinus sylvestris Seed Trees from Investigated Stands in Southern Poland. Forests 2021, 12, 1323. https://doi.org/10.3390/f12101323
Wartalska P, Oszako T, Bakier S, Belbahri L, Malewski T, Hsiang T, Popowska-Nowak E, Nowakowska J. Dothistroma septosporum Not Detected in Pinus sylvestris Seed Trees from Investigated Stands in Southern Poland. Forests. 2021; 12(10):1323. https://doi.org/10.3390/f12101323
Chicago/Turabian StyleWartalska, Pola, Tomasz Oszako, Sławomir Bakier, Lassaâd Belbahri, Tadeusz Malewski, Tom Hsiang, Elżbieta Popowska-Nowak, and Justyna Nowakowska. 2021. "Dothistroma septosporum Not Detected in Pinus sylvestris Seed Trees from Investigated Stands in Southern Poland" Forests 12, no. 10: 1323. https://doi.org/10.3390/f12101323
APA StyleWartalska, P., Oszako, T., Bakier, S., Belbahri, L., Malewski, T., Hsiang, T., Popowska-Nowak, E., & Nowakowska, J. (2021). Dothistroma septosporum Not Detected in Pinus sylvestris Seed Trees from Investigated Stands in Southern Poland. Forests, 12(10), 1323. https://doi.org/10.3390/f12101323