Fungi Detected in the Previous Year’s Leaf Petioles of Fraxinus excelsior and Their Antagonistic Potential against Hymenoscyphus fraxineus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Sampling
2.2. Fungal Isolation and Identification
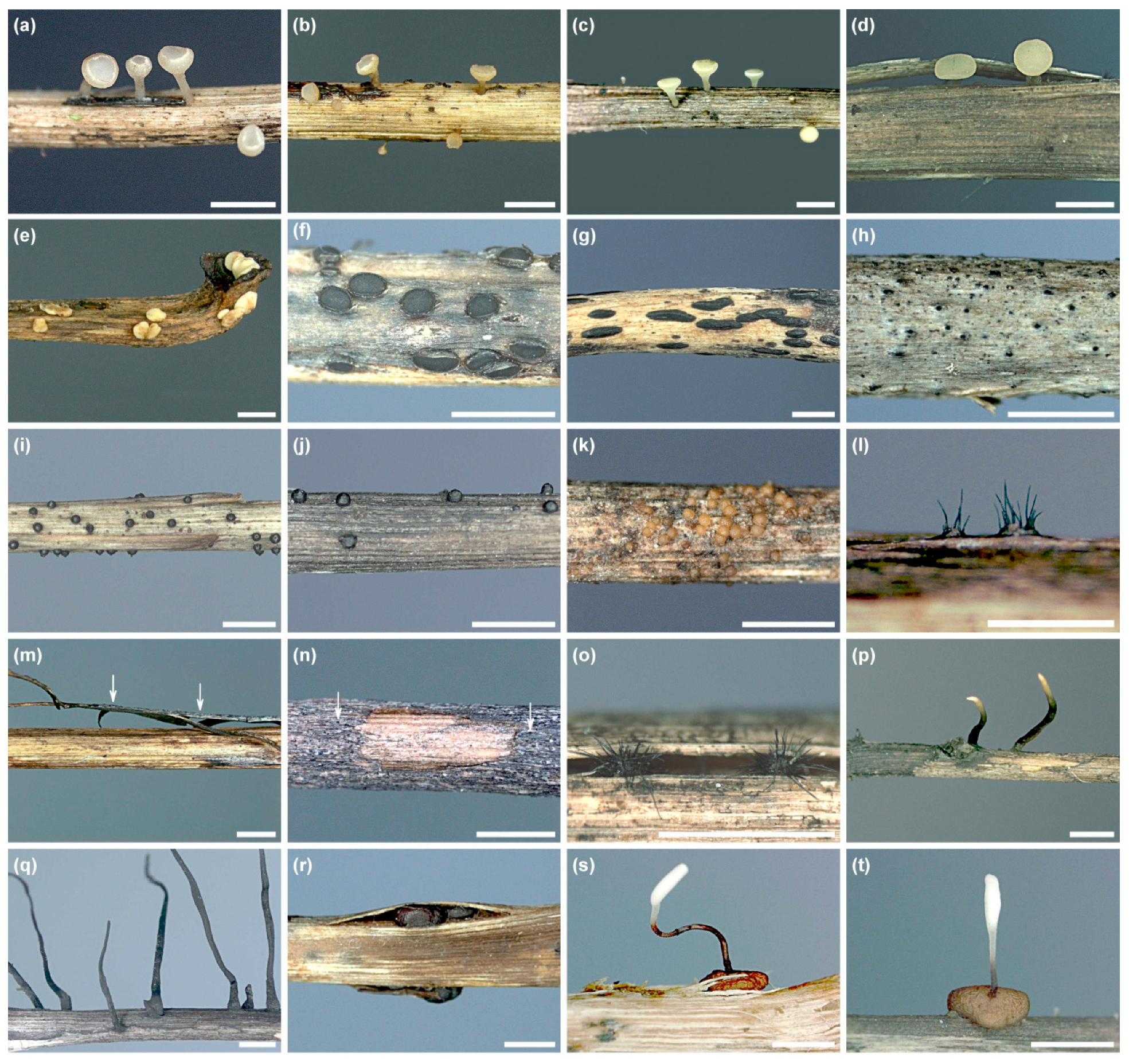
2.3. In Situ Inventory of Fungi from Fruit Bodies on Petioles
2.4. DNA Extraction, PCR and Sequencing
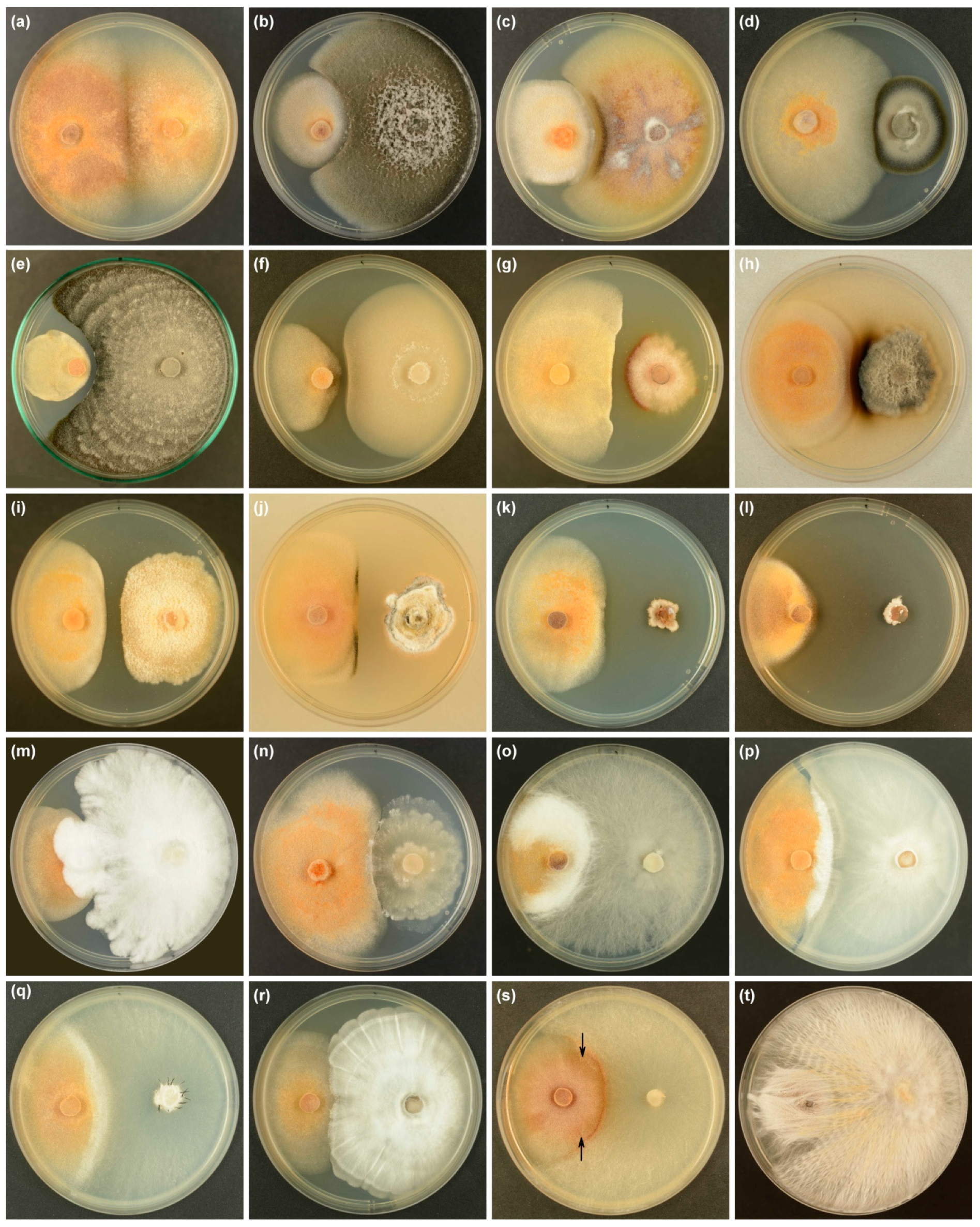
2.5. Dual-Culture Assays
2.6. Data Analyses
3. Results
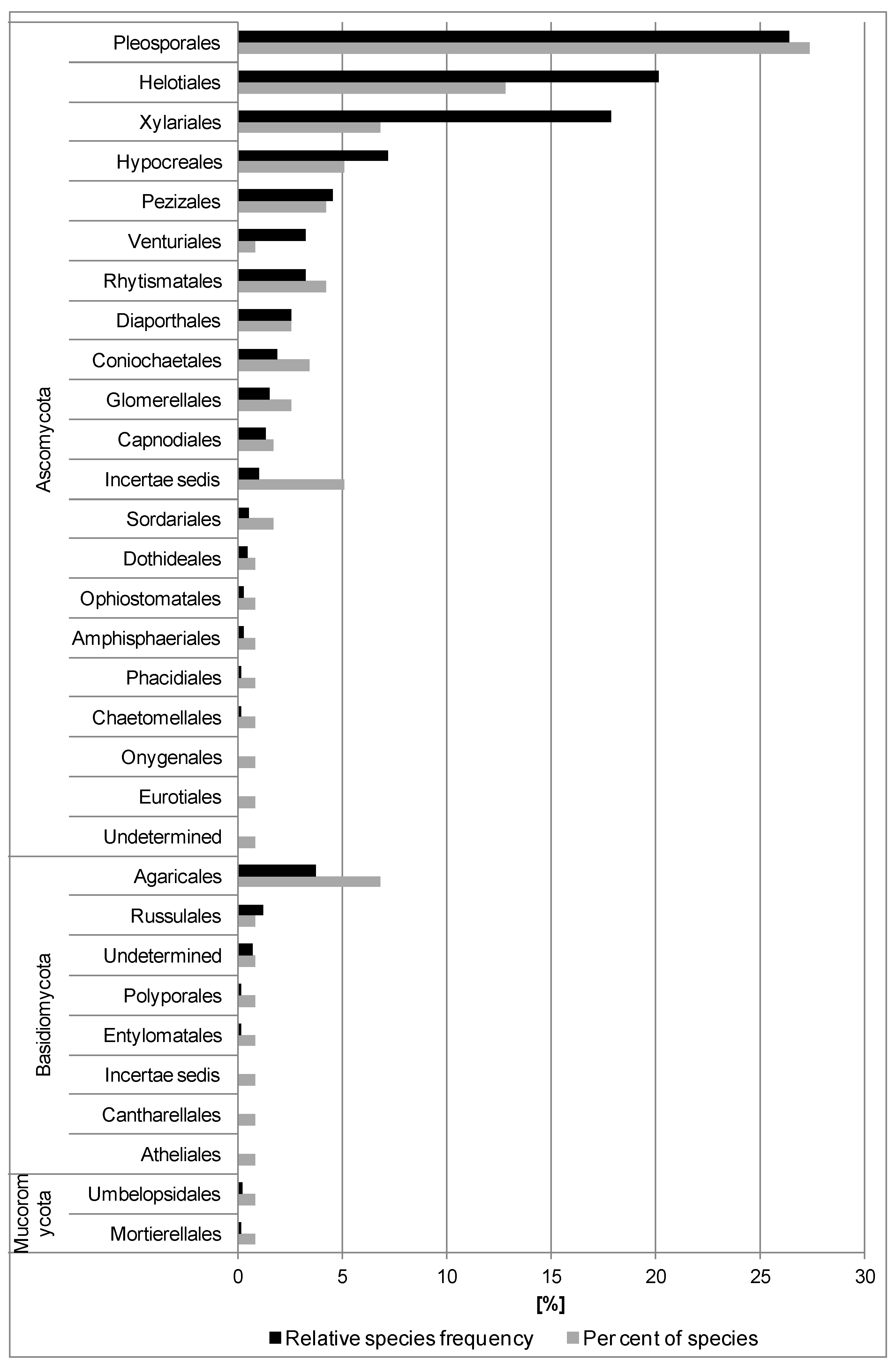
3.1. Fungi Isolated from the Previous Year’s Leaf Petioles
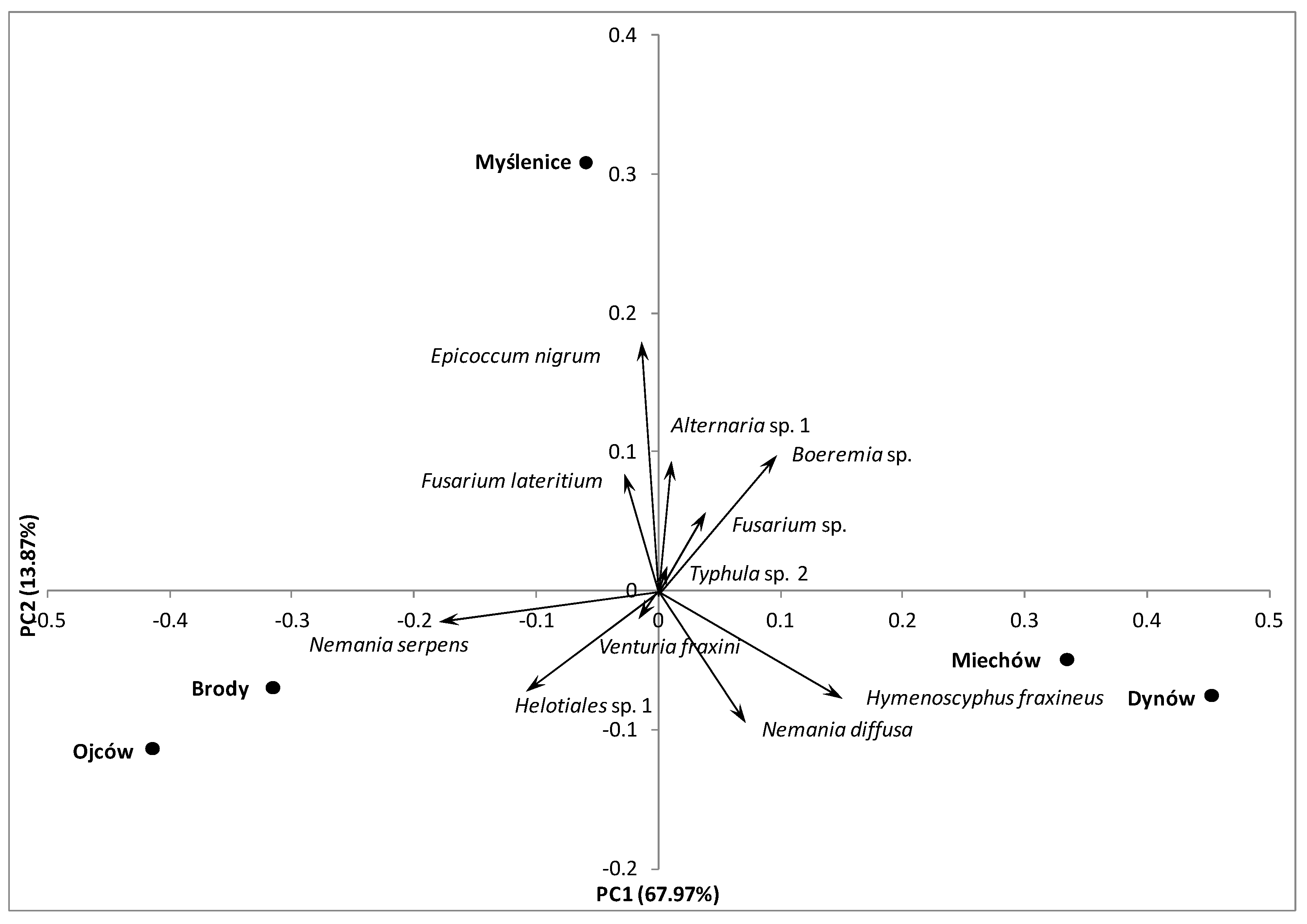
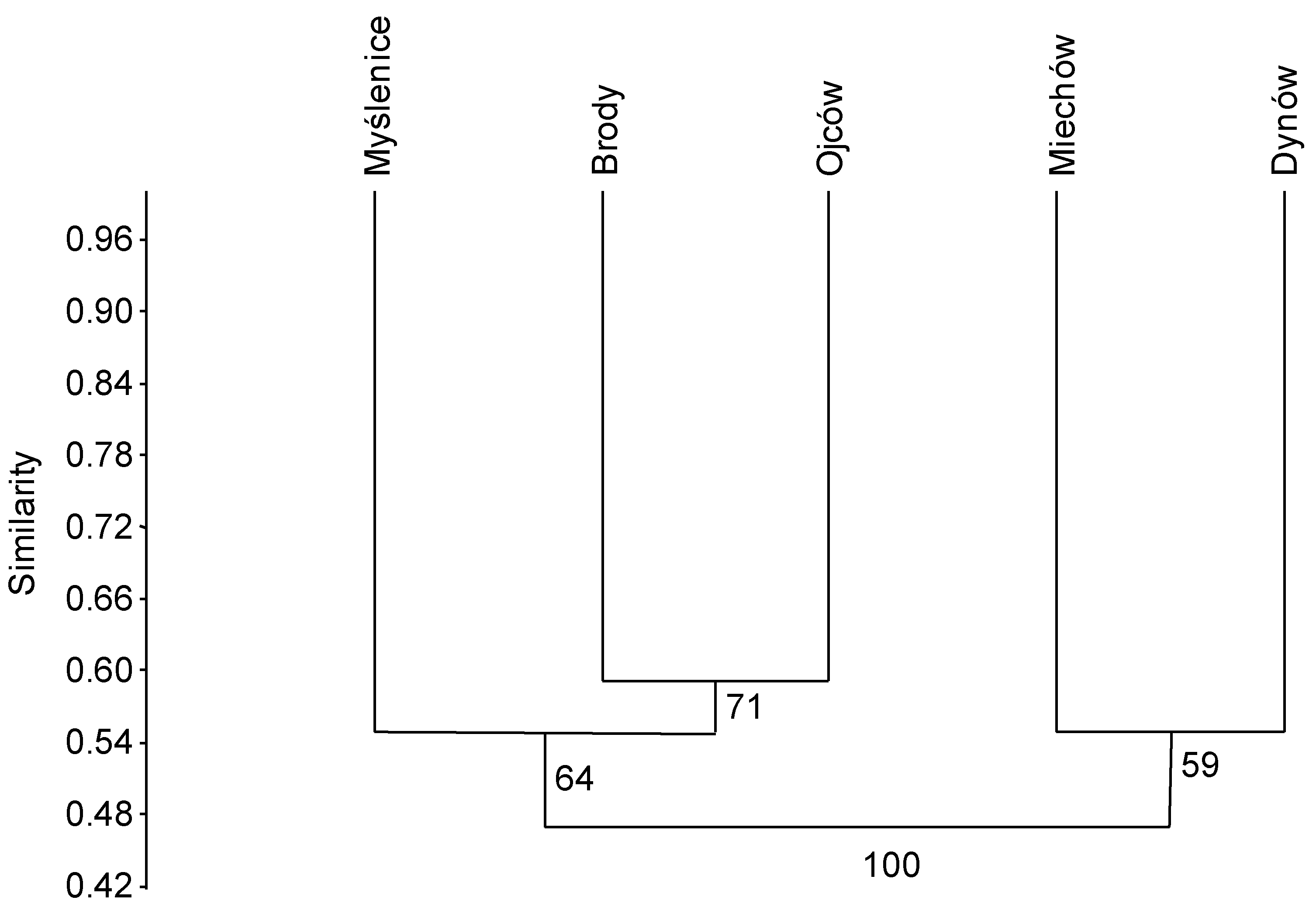
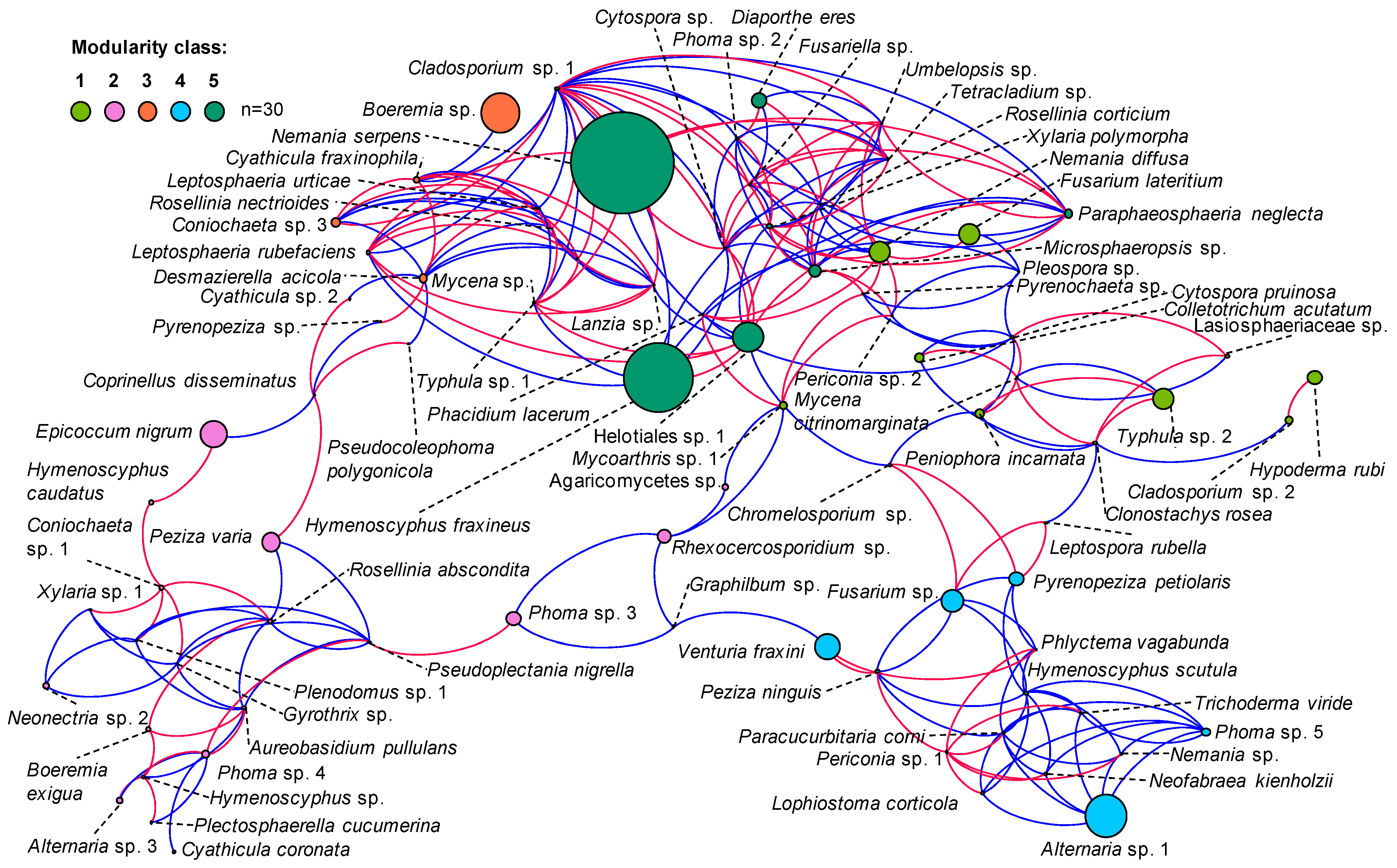
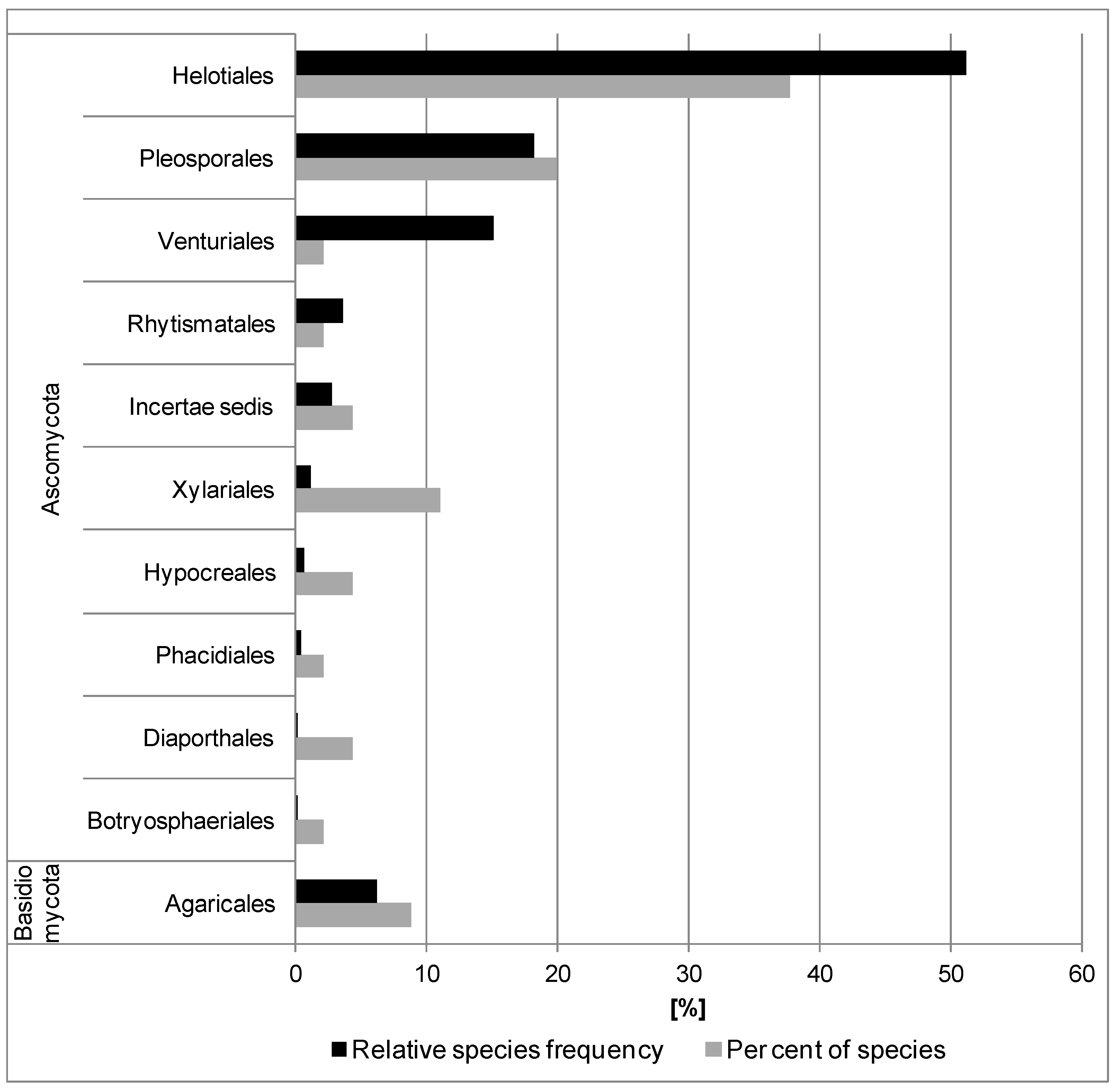
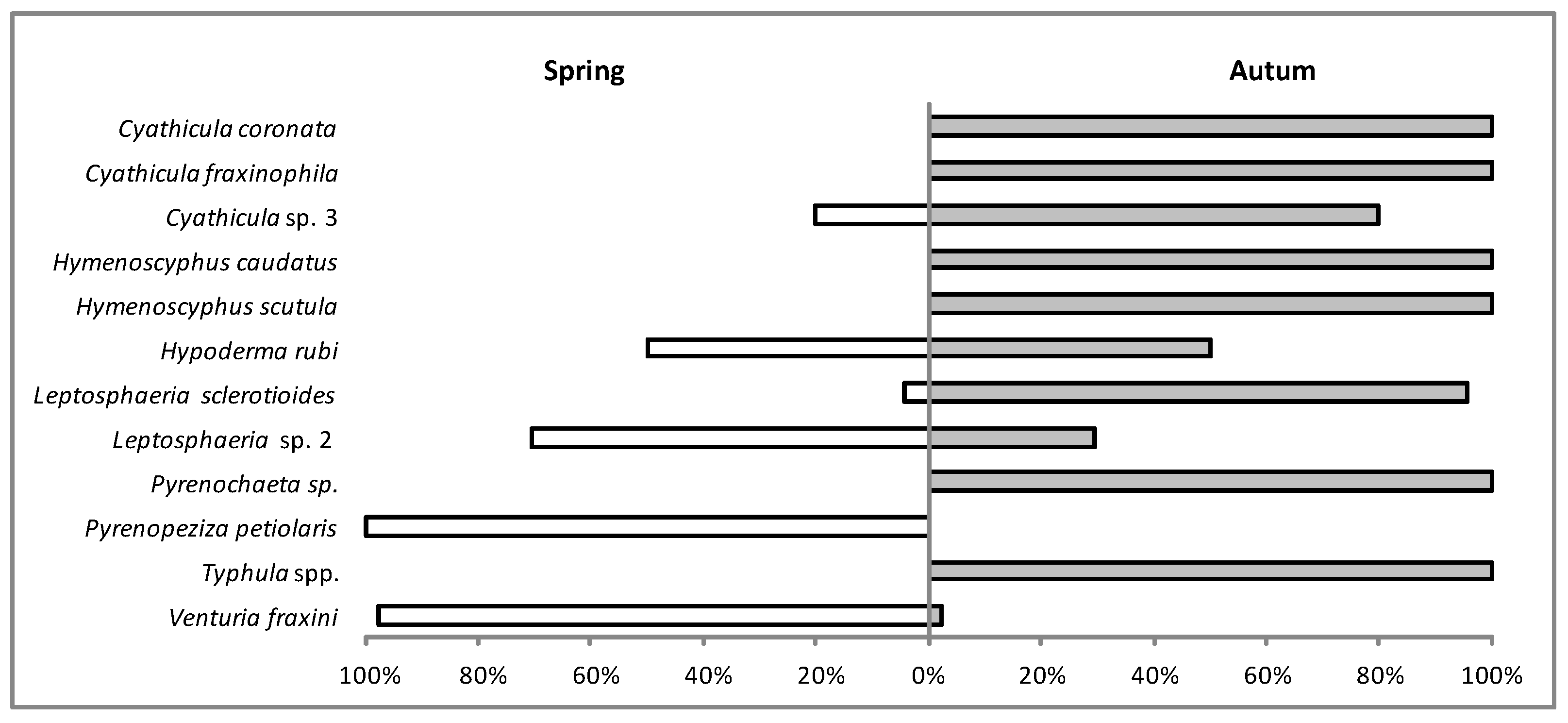
3.2. In Situ Inventory of Fungi Based on Fruit Bodies on Petioles
3.3. Dual-Culture Assays
4. Discussion
4.1. Fungal Diversity
4.2. Competition between H. fraxineus and Saprotrophs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kowalski, T. O zamieraniu jesionów. Tryb. Leśnika 2001, 4, 6–7. [Google Scholar]
- Kowalski, T. Chalara fraxinea sp. nov. associated with dieback of ash (Fraxinus excelsior) in Poland. For. Pathol. 2006, 36, 264–270. [Google Scholar] [CrossRef]
- Przybył, K. Fungi associated with necrotic apical parts of Fraxinus excelsior shoots. For. Pathol. 2002, 32, 387–394. [Google Scholar] [CrossRef]
- Halmschlager, E.; Kirisits, T. First report of the ash dieback pathogen Chalara fraxinea on Fraxinus excelsior in Austria. Plant Pathol. 2008, 57, 1177. [Google Scholar] [CrossRef]
- Gross, A.; Holdenrieder, O.; Pautasso, M.; Queloz, V.; Sieber, T.N. Hymenoscyphus pseudoalbidus, the causal agent of European ash dieback. Mol. Plant Pathol. 2014, 15, 5–21. [Google Scholar] [CrossRef]
- Timmermann, V.; Nagy, N.E.; Hietala, A.M.; Børja, I.; Solheim, H. Progression of ash dieback in Norway related to tree age, disease history and regional aspects. Balt. For. 2017, 23, 150–158. [Google Scholar]
- Hietala, A.M.; Børja, I.; Cross, H.; Nagy, N.E.; Solheim, H.; Timmermann, V.; Vivian-Smith, A. Dieback of European Ash: What Can We Learn from the Microbial Community and Species-Specific Traits of Endophytic Fungi Associated with Ash? In Endophytes of Forest Trees. Biology and Applications; Pirttilä, A.M., Frank, A.C., Eds.; Forestry Sciences; Springer International Publishing: Cham, Switzerland, 2018; Volume 86, pp. 229–258. ISBN 978-3-319-89832-2. [Google Scholar]
- Enderle, R. An overview of ash (Fraxinus spp.) and the ash dieback disease in Europe. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2019, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Baral, H.-O.; Queloz, V.; Hosoya, T. Hymenoscyphus fraxineus, the correct scientific name for the fungus causing ash dieback in Europe. IMA Fungus 2014, 5, 79–80. [Google Scholar] [CrossRef] [PubMed]
- Cleary, M.; Nguyen, D.; Marčiulynienė, D.; Berlin, A.; Vasaitis, R.; Stenlid, J. Friend or foe? Biological and ecological traits of the European ash dieback pathogen Hymenoscyphus fraxineus in its native environment. Sci. Rep. 2016, 6, 21895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, T.; Okane, I.; Ishiga, Y.; Degawa, Y.; Hosoya, T.; Yamaoka, Y. The life cycle of Hymenoscyphus fraxineus on Manchurian ash, Fraxinus mandshurica, in Japan. Mycoscience 2019, 60, 89–94. [Google Scholar] [CrossRef]
- Kowalski, T. Zamieranie jesionu—Aspekty taksonomiczne sprawcy choroby. Sylwan 2012, 156, 262–269. [Google Scholar]
- Hauptman, T.; Piškur, B.; de Groot, M.; Ogris, N.; Ferlan, M.; Jurc, D. Temperature effect on Chalara fraxinea: Heat treatment of saplings as a possible disease control method. For. Pathol. 2013, 43, 360–370. [Google Scholar] [CrossRef]
- Cleary, M.R.; Daniel, G.; Stenlid, J. Light and scanning electron microscopy studies of the early infection stages of Hymenoscyphus pseudoalbidus on Fraxinus excelsior. Plant Pathol. 2013, 62, 1294–1301. [Google Scholar] [CrossRef]
- Haňáčková, Z.; Koukol, O.; Čmoková, A.; Zahradník, D.; Havrdová, L. Direct evidence of Hymenoscyphus fraxineus infection pathway through the petiole-shoot junction. For. Pathol. 2017, 47, e12370. [Google Scholar] [CrossRef]
- Gross, A.; Holdenrieder, O. On the longevity of Hymenoscyphus pseudoalbidus in petioles of Fraxinus excelsior. For. Pathol. 2013, 43, 168–170. [Google Scholar] [CrossRef]
- Kirisits, T. Ascocarp formation of Hymenoscyphus fraxineus on several-year-old pseudosclerotial leaf rachises of Fraxinus excelsior. For. Pathol. 2015, 45, 254–257. [Google Scholar] [CrossRef]
- Kowalski, T.; Kehr, R. Stand des Eschentriebsterbens und zunehmende Schäden an Buche und Berg-Ahorn. In Jahrbuch der Baumpflege; Dujesiefken, D., Ed.; Haymarket Media: Braunschweig, Germany, 2016; pp. 63–83. [Google Scholar]
- Hauptman, T.; Celar, F.; de Groot, M.; Jurc, D. Application of fungicides and urea for control of ash dieback. iFor. Biogeosci. For. 2015, 8, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Bartha, B.; Mayer, A.; Lenz, H.D. Acceleration of ash petiole decomposition to reduce Hymenoscyphus fraxineus apothecia growth—A feasible method for the deprivation of fungal substrate. Balt. For. 2017, 23, 82–88. [Google Scholar]
- Marzano, M.; Woodcock, P.; Quine, C.P. Dealing with dieback: Forest manager attitudes towards developing resistant ash trees in the United Kingdom. Forestry 2019, 92, 554–567. [Google Scholar] [CrossRef]
- Noble, R.; Woodhall, J.W.; Dobrovin-Pennington, A.; Perkins, K.; Somoza-Valdeolmillos, E.; Gómez, H.L.; Lu, Y.; Macarthur, R.; Henry, C.M. Control of Hymenoscyphus fraxineus, the causal agent of ash dieback, using composting. For. Pathol. 2019, 49, e12568. [Google Scholar] [CrossRef]
- Berg, G.; Köberl, M.; Rybakova, D.; Müller, H.; Grosch, R.; Smalla, K. Plant microbial diversity is suggested as the key to future biocontrol and health trends. FEMS Microbiol. Ecol. 2017, 93, 50. [Google Scholar] [CrossRef]
- Terhonen, E.; Kovalchuk, A.; Zarsav, A.; Asiegbu, F.O. Biocontrol Potential of Forest Tree Endophytes. In Endophytes of Forest Trees. Biology and Applications; Pirttilä, A.M., Frank, A.C., Eds.; Forestry Sciences; Springer International Publishing: Cham, Switzerland, 2018; Volume 86, pp. 283–318. ISBN 978-3-319-89832-2. [Google Scholar]
- Collinge, D.B.; Jørgensen, H.J.L.; Latz, M.A.C.; Manzotti, A.; Ntana, F.; Rojas, E.C.; Jensen, B. Searching for novel fungal biological control agents for plant disease control among endophytes. In Endophytes for a Growing World; Cambridge University Press: Cambridge, UK, 2019; pp. 25–51. ISBN 9781108607667. [Google Scholar]
- Schlegel, M.; Dubach, V.; von Buol, L.; Sieber, T.N. Effects of endophytic fungi on the ash dieback pathogen. FEMS Microbiol. Ecol. 2016, 92, fiw142. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Haas, S.; Junker, C.; Andrée, N.; Schobert, M. Fungal endophytes are involved in multiple balanced antagonisms. Curr. Sci. 2015, 109, 39–45. [Google Scholar] [CrossRef]
- Haňáčková, Z.; Havrdová, L.; Černý, K.; Zahradník, D.; Koukol, O. Fungal endophytes in ash shoots—Diversity and inhibition of Hymenoscyphus fraxineus. Balt. For. 2017, 23, 89–106. [Google Scholar]
- Kosawang, C.; Amby, D.B.; Bussaban, B.; McKinney, L.V.; Xu, J.; Kjær, E.D.; Collinge, D.B.; Nielsen, L.R. Fungal communities associated with species of Fraxinus tolerant to ash dieback, and their potential for biological control. Fungal Biol. 2018, 122, 110–120. [Google Scholar] [CrossRef]
- Halecker, S.; Wennrich, J.-P.; Rodrigo, S.; Andrée, N.; Rabsch, L.; Baschien, C.; Steinert, M.; Stadler, M.; Surup, F.; Schulz, B. Fungal endophytes for biocontrol of ash dieback: The antagonistic potential of Hypoxylon rubiginosum. Fungal Ecol. 2020, 45, 100918. [Google Scholar] [CrossRef]
- Gartner, T.B.; Cardon, Z.G. Decomposition dynamics in mixed-species leaf litter. Oikos 2004, 104, 230–246. [Google Scholar] [CrossRef]
- Scherer-Lorenzen, M.; Luis Bonilla, J.; Potvin, C. Tree species richness affects litter production and decomposition rates in a tropical biodiversity experiment. Oikos 2007, 116, 2108–2124. [Google Scholar] [CrossRef]
- Jonard, M.; Andre, F.; Ponette, Q. Tree species mediated effects on leaf litter dynamics in pure and mixed stands of oak and beech. Can. J. For. Res. 2008, 38, 528–538. [Google Scholar] [CrossRef]
- Purahong, W.; Hyde, K.D. Effects of fungal endophytes on grass and non-grass litter decomposition rates. Fungal Divers. 2011, 47, 1–7. [Google Scholar] [CrossRef]
- Hoeber, S.; Fransson, P.; Weih, M.; Manzoni, S. Leaf litter quality coupled to Salix variety drives litter decomposition more than stand diversity or climate. Plant Soil 2020, 453, 313–328. [Google Scholar] [CrossRef]
- Melillo, J.M.; Aber, J.D.; Muratore, J.F. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 1982, 63, 621–626. [Google Scholar] [CrossRef]
- Osono, T.; Takeda, H. Decomposition of organic chemical components in relation to nitrogen dynamics in leaf litter of 14 tree species in a cool temperate forest. Ecol. Res. 2005, 20, 41–49. [Google Scholar] [CrossRef]
- Nykvist, N. Leaching and decomposition of litter I. Experiments on leaf litter of Fraxinus excelsior. Oikos 1959, 10, 190. [Google Scholar] [CrossRef]
- Błońska, E.; Piaszczyk, W.; Staszel, K.; Lasota, J. Enzymatic activity of soils and soil organic matter stabilization as an effect of components released from the decomposition of litter. Appl. Soil Ecol. 2021, 157, 103723. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Kull, O.; Tenhunen, J.D. An analysis of light effects on foliar morphology, physiology, and light interception in temperate deciduous woody species of contrasting shade tolerance. Tree Physiol. 1998, 18, 681–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotrufo, M.F.; Briones, M.J.I.; Ineson, P. Elevated CO2 affects field decomposition rate and palatability of tree leaf litter: Importance of changes in substrate quality. Soil Biol. Biochem. 1998, 30, 1565–1571. [Google Scholar] [CrossRef]
- Jacob, M.; Viedenz, K.; Polle, A.; Thomas, F.M. Leaf litter decomposition in temperate deciduous forest stands with a decreasing fraction of beech (Fagus sylvatica). Oecologia 2010, 164, 1083–1094. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, Ü. Differences in chemical composition relative to functional differentiation between petioles and laminas of Fraxinus excelsior. Tree Physiol. 1999, 19, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Conway, D.R.; Frankland, J.C.; Saunders, V.A.; Wilson, D.R. Effects of elevated atmospheric CO2 on fungal competition and decomposition of Fraxinus excelsior litter in laboratory microcosms. Mycol. Res. 2000, 104, 187–197. [Google Scholar] [CrossRef]
- Dilly, O.; Bartsch, S.; Rosenbrock, P.; Buscot, F.; Munch, J.C. Shifts in physiological capabilities of the microbiota during the decomposition of leaf litter in a black alder (Alnus glutinosa (Gaertn.) L.) forest. Soil Biol. Biochem. 2001, 33, 921–930. [Google Scholar] [CrossRef]
- Osono, T.; Takeda, H. Comparison of litter decomposing ability among diverse fungi in a cool temperate deciduous forest in Japan. Mycologia 2002, 94, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Shirouzu, T.; Hirose, D.; Fukasawa, Y.; Tokumasu, S. Fungal succession associated with the decay of leaves of an evergreen oak, Quercus myrsinaefolia. Fungal Divers. 2009, 34, 87–109. [Google Scholar]
- Voříšková, J.; Baldrian, P. Fungal community on decomposing leaf litter undergoes rapid successional changes. ISME J. 2013, 7, 477–486. [Google Scholar] [CrossRef]
- Osono, T. Leaf litter decomposition of 12 tree species in a subtropical forest in Japan. Ecol. Res. 2017, 32, 413–422. [Google Scholar] [CrossRef]
- Rosales-Castillo, J.A.; Oyama, K.; Vázquez-Garcidueñas, M.S.; Aguilar-Romero, R.; García-Oliva, F.; Vázquez-Marrufo, G. Fungal community and ligninolytic enzyme activities in Quercus deserticola Trel. litter from forest fragments with increasing levels of disturbance. Forests 2017, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Tennakoon, D.S.; Gentekaki, E.; Jeewon, R.; Kuo, C.H.; Promputtha, I.; Hyde, K.D. Life in leaf litter: Fungal community succession during decomposition. Mycosphere 2021, 12, 406–429. [Google Scholar] [CrossRef]
- Davydenko, K.; Vasaitis, R.; Stenlid, J.; Menkis, A. Fungi in foliage and shoots of Fraxinus excelsior in eastern Ukraine: A first report on Hymenoscyphus pseudoalbidus. For. Pathol. 2013, 43, 462–467. [Google Scholar] [CrossRef]
- Koukol, O.; Haňáčková, Z.; Dvořák, M.; Havrdová, L. Unseen, but still present in Czechia: Hymenoscyphus albidus detected by real-time PCR, but not by intensive sampling. Mycol. Prog. 2016, 15, 6. [Google Scholar] [CrossRef]
- Cross, H.; Sønstebø, J.H.; Nagy, N.E.; Timmermann, V.; Solheim, H.; Børja, I.; Kauserud, H.; Carlsen, T.; Rzepka, B.; Wasak, K.; et al. Fungal diversity and seasonal succession in ash leaves infected by the invasive ascomycete Hymenoscyphus fraxineus. New Phytol. 2017, 213, 1405–1417. [Google Scholar] [CrossRef]
- Trapiello, E.; Schoebel, C.N.; Rigling, D. Fungal community in symptomatic ash leaves in Spain. Balt. For. 2017, 23, 68–73. [Google Scholar]
- Kowalski, T.; Kraj, W.; Bednarz, B.; Rossa, R. The association of Boeremia lilacis with necrotic lesions on shoots and leaf petioles and its pathogenicity towards Fraxinus excelsior. Eur. J. Plant Pathol. 2019, 154, 961–974. [Google Scholar] [CrossRef] [Green Version]
- Wicklow, D.T. Interference competition. In The Fungal Community: Its Organization and Role in the Ecosystem; Carroll, G.C., Wicklow, D.T., Eds.; Marcel Dekker: New York, NY, USA, 1992; pp. 265–274. [Google Scholar]
- Zhang, L.; Andersen, K.H.; Dieckmann, U.; Brännström, Å. Four types of interference competition and their impacts on the ecology and evolution of size-structured populations and communities. J. Theor. Biol. 2015, 380, 280–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalski, T.; Czekaj, A. Symptomy chorobowe i grzyby na zamierających jesionach (Fraxinus excelsior L.) w drzewostanach Nadleśnictwa Staszów. Disease symptoms and fungi on dying ash trees (Fraxinus excelsior L.) in Staszów Forest District stands. Leśne Pr. Badaw. 2010, 71, 357–368. [Google Scholar]
- Lacap, D.C.; Hyde, K.D.; Liew, E.C.Y. An evaluation of the fungal “morphotype” concept based on ribosomal DNA sequences. Fungal Divers. 2003, 12, 53–66. [Google Scholar]
- Kowalski, T.; Bartnik, C. Morphologial variation in colonies of Chalara fraxinea isolated from ash (Fraxinus excelsior L.) stems with symptoms of dieback and effects of temperature on colony growth and structure. Acta Agrobot. 2010, 63, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Kirisits, T.; Dämpfle, L.; Kräutler, K. Hymenoscyphus albidus is not associated with an anamorphic stage and displays slower growth than Hymenoscyphus pseudoalbidus on agar media. For. Pathol. 2013, 43, 386–389. [Google Scholar] [CrossRef]
- Kirk, P.; Cannon, P.; Minter, D.; Stalpers, J. Ainthworth and Bisby’s Dictionary of the Fungi, 10th ed.; Kirk, P., Cannon, P., Minter, D., Stalpers, J., Eds.; CABI: Wallingford, UK, 2008; ISBN 978-0-85199-826-8. [Google Scholar]
- Baral, H.-O.; Bemmann, M. Hymenoscyphus fraxineus vs. Hymenoscyphus albidus—A comparative light microscopic study on the causal agent of European ash dieback and related foliicolous, stroma-forming species. Mycology 2014, 5, 228–290. [Google Scholar] [CrossRef] [Green Version]
- Koske, R.E. Typhula erythropus: II. Sclerotial germination and basidiocarp production. Mycologia 1975, 67, 128–146. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, F.; Hsiang, T. Fertile sporophore production of Typhula phacorrhiza in the field is related to temperatures near freezing. Can. J. Microbiol. 2006, 52, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Kraj, W.; Kowalski, T. Genetic variability of Hymenoscyphus pseudoalbidus on ash leaf rachises in leaf litter of forest stands in Poland. J. Phytopathol. 2014, 162, 218–227. [Google Scholar] [CrossRef]
- Index Fungorum—Search Page. Available online: http://www.indexfungorum.org/names/Names.asp (accessed on 29 August 2021).
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, J.; Platas, G.; Paulus, B.; Bills, G.F. High-throughput culturing of fungi from plant litter by a dilution-to-extinction technique. FEMS Microbiol. Ecol. 2007, 60, 521–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BIOEDIT: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/ NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Lahlali, R.; Hijri, M. Screening, identification and evaluation of potential biocontrol fungal endophytes against Rhizoctonia solani AG3 on potato plants. FEMS Microbiol. Lett. 2010, 311, 152–159. [Google Scholar] [CrossRef]
- Martínez-Álvarez, P.; Fernández-González, R.A.; Sanz-Ros, A.V.; Pando, V.; Diez, J.J. Two fungal endophytes reduce the severity of pitch canker disease in Pinus radiata seedlings. Biol. Control 2016, 94, 1–10. [Google Scholar] [CrossRef]
- Bilański, P.; Kowalski, T. Fungal endophytes in Fraxinus excelsior petioles and their in vitro antagonistic potential against the ash dieback pathogen Hymenoscyphus fraxineus. Microbiol. Res. 2021. accepted. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication, 1st ed.; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Camargo, J.A. Must dominance increase with the number of subordinate species in competitive interactions? J. Theor. Biol. 1993, 161, 537–542. [Google Scholar] [CrossRef]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open source software for exploring and manipulating networks. In Proceedings of the International AAAI Conference on Weblogs and Social Media, San Jose, CA, USA, 17–20 May 2009; pp. 361–362. [Google Scholar]
- STATISTICA (Data Analysis Software System); StatSoft Inc.: Tulsa, OK, USA, 2014.
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core: Vienna, Austria, 2018. [Google Scholar]
- Osono, T. Ecology of ligninolytic fungi associated with leaf litter decomposition. Ecol. Res. 2007, 22, 955–974. [Google Scholar] [CrossRef]
- Kubartová, A.; Ranger, J.; Berthelin, J.; Beguiristain, T. Diversity and decomposing ability of saprophytic fungi from temperate forest litter. Microb. Ecol. 2009, 58, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Frankland, J.C. Fungal succession—Unravelling the unpredictable. Mycol. Res. 1998, 102, 1–15. [Google Scholar] [CrossRef]
- Paulus, B.; Gadek, P.; Hyde, K. Successional patterns of microfungi in fallen leaves of Ficus pleurocarpa (Moraceae) in an Australian tropical rain forest. Biotropica 2006, 38, 051207072004001. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Rajeshkumar, K.C.; Hawksworth, D.L.; Madrid, H.; Kirk, P.M.; Braun, U.; Singh, R.V.; Crous, P.W.; Kukwa, M.; et al. Notes for genera: Ascomycota. Fungal Divers. 2017, 86, 1–594. [Google Scholar] [CrossRef] [Green Version]
- Jahn, H.T. Pilze die an Holz Wachsen, 1st ed.; Verlag Busse: Herford, Germany, 1979; ISBN 978-3871208539. [Google Scholar]
- Petrini, L.; Petrini, O. Xylariaceous fungi as endophytes. Sydowia 1985, 38, 216–234. [Google Scholar]
- Granmo, A.; Læssøe, T.; Schumacher, T. The genus Nemania s.l. (Xylariaceae) in Norden. Sommerfeltia 1999, 27, 1. [Google Scholar] [CrossRef]
- Kowalski, T.; Kraj, W.; Bednarz, B. Fungi on stems and twigs in initial and advanced stages of dieback of European ash (Fraxinus excelsior) in Poland. Eur. J. For. Res. 2016, 135, 565–579. [Google Scholar] [CrossRef] [Green Version]
- Linaldeddu, B.T.; Bottecchia, F.; Bregant, C.; Maddau, L.; Montecchio, L. Diplodia fraxini and Diplodia subglobosa: The main species associated with cankers and dieback of Fraxinus excelsior in north-eastern Italy. Forests 2020, 11, 883. [Google Scholar] [CrossRef]
- Schlegel, M.; Queloz, V.; Sieber, T.N. The Endophytic Mycobiome of European Ash and Sycamore Maple Leaves—Geographic Patterns, Host Specificity and Influence of Ash Dieback. Front. Microbiol. 2018, 9, 2345. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Bussaban, B.; Paulus, B.; Crous, P.W.; Lee, S.; Mckenzie, E.H.C.; Photita, W.; Lumyong, S. Diversity of saprobic microfungi. Biodivers. Conserv. 2007, 16, 7–35. [Google Scholar] [CrossRef]
- Promputtha, I.; Lumyong, S.; Dhanasekaran, V.; McKenzie, E.H.C.; Hyde, K.D.; Jeewon, R. A phylogenetic evaluation of whether endophytes become saprotrophs at host senescence. Microb. Ecol. 2007, 53, 579–590. [Google Scholar] [CrossRef]
- De Silva, N.I.; Phillips, A.J.L.; Liu, J.-K.; Lumyong, S.; Hyde, K.D. Phylogeny and morphology of Lasiodiplodia species associated with Magnolia forest plants. Sci. Rep. 2019, 9, 14355. [Google Scholar] [CrossRef]
- Osono, T. Role of phyllosphere fungi of forest trees in the development of decomposer fungal communities and decomposition processes of leaf litter. Can. J. Microbiol. 2006, 52, 701–716. [Google Scholar] [CrossRef]
- Sivanesan, A. The Bitunicate Ascomycetes and Their Anamorphs, 1st ed.; Lubrecht & Cramer: Schaan, Liechtenstein, 1984; ISBN 978-3-76821-329-5. [Google Scholar]
- Schubert, K.; Rischel, A.; Braun, D.U. A monograph of Fusicladium s.lat. (Hyphomycetes). Schlechtendalia 2013, 9, 1–132. [Google Scholar]
- Agan, A.; Drenkhan, R.; Adamson, K.; Tedersoo, L.; Solheim, H.; Børja, I.; Matsiakh, I.; Timmermann, V.; Nagy, N.E.; Hietala, A.M. The relationship between fungal diversity and invasibility of a foliar niche—The case of ash dieback. J. Fungi 2020, 6, 150. [Google Scholar] [CrossRef] [PubMed]
- Bakys, R.; Vasaitis, R.; Barklund, P.; Ihrmark, K.; Stenlid, J. Investigations concerning the role of Chalara fraxinea in declining Fraxinus excelsior. Plant Pathol. 2009, 58, 284–292. [Google Scholar] [CrossRef]
- Carpenter, S.E. Monograph of Crocicreas (Ascomycetes, Helotiales, Leotiaceae. Mem. N. Y. Bot. Gard. 1981, 33, 1–290. [Google Scholar]
- Baral, H.-O.; Krieglsteriner, G.J. Bausteine zu einer Askomyzeten-Flora der Bundesrepublik Deutschland: In Süddeutschland gefundene inoperculate Discomyzeten mit taxonomische, okologischen, chorologischen Hinweisen und einer Farbtafel. Beih. Z. Mykol. 1985, 6, 1–160. [Google Scholar]
- Minter, D.W. Hypoderma rubi. In CMI Descriptions of Pathogenic Fungi and Bacteria; Commonwealth Mycological Institute: Kew, UK, 1984; pp. 1–2. [Google Scholar]
- Thomas, P.A. Biological Flora of the British Isles: Fraxinus excelsior. J. Ecol. 2016, 104, 1158–1209. [Google Scholar] [CrossRef]
- Pastirčák, M.; Pastirčáková, K. Hypoderma rubi on two new hosts in Slovakia. Mycotaxon 2017, 132, 849–855. [Google Scholar] [CrossRef]
- Hütter, R. Untersuchungen über die Gattung Pyrenopeziza Fuck. J. Phytopathol. 1958, 33, 1–54. [Google Scholar] [CrossRef]
- Pastirčáková, K.; Ivanová, H.; Pastirčák, M. Species diversity of fungi on damaged branches and leaves of ashes (Fraxinus spp.) in different types of stands in Slovakia. Cent. Eur. For. J. 2018, 64, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Pastircáková, K.; Adamcíková, K.; Barta, M.; Pažitnỳ, J.; Hot’ka, P.; Sarvašová, I.; Horáková, M.K. Host range of Hymenoscyphus fraxineus in Slovak Arboreta. Forests 2020, 11, 596. [Google Scholar] [CrossRef]
- Hietala, A.M.; Børja, I.; Solheim, H.; Nagy, N.E.; Timmermann, V. Propagule pressure build-up by the invasive Hymenoscyphus fraxineus following its introduction to an ash forest inhabited by the native Hymenoscyphus albidus. Front. Plant Sci. 2018, 9, 1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieber, T.N. Endophytic fungi in forest trees: Are they mutualists? Fungal Biol. Rev. 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Kowalski, T. Zur Pilzflora toter Kiefernnadeln. Z. Mykol. 1988, 54, 159–173. [Google Scholar]
- Boerema, G.H.; Gruyter, J.; Noordeloos, M.E.; Hamers, M.E.C. Phoma Identification Manual. Differentiation of Specific and Infra-Specific Taxa in Culture; Boerema, G.H., de Gruyter, J., Noordeloos, M.E., Hamers, M.E.C., Eds.; CABI: Wallingford, UK, 2004; ISBN 978-0-85199-743-8. [Google Scholar]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Phookamsak, R.; Wanasinghe, D.N.; Hongsanan, S.; Phukhamsakda, C.; Huang, S.-K.; Tennakoon, D.S.; Norphanphoun, C.; Camporesi, E.; Bulgakov, T.S.; Promputtha, I.; et al. Towards a natural classification of Ophiobolus and ophiobolus-like taxa; introducing three novel genera Ophiobolopsis, Paraophiobolus and Pseudoophiobolus in Phaeosphaeriaceae (Pleosporales). Fungal Divers. 2017, 87, 299–339. [Google Scholar] [CrossRef]
- Hohmeyer, H. Ein Schlüssel zu den europaischen Arten der Gattung Peziza L. Z. Mykol. 1986, 52, 161–188. [Google Scholar]
- Remsberg, R.E. Studies in the genus Typhula. Mycologia 1940, 32, 52–96. [Google Scholar] [CrossRef]
- Wu, C.; Hsiang, T. Pathogenicity and formulation of Typhula phacorrhiza, a biocontrol agent of gray snow mold. Plant Dis. 1998, 82, 1003–1006. [Google Scholar] [CrossRef]
- Zabel, R.; Morrell, J. Wood Microbiology: Decay and Its Prevention, 1st ed.; Academic Press: San Diego, CA, USA, 1992; ISBN 978-0-32313-946-5. [Google Scholar]
- Worrall, J.J.; Anagnost, S.E.; Zabel, R.A. Comparison of wood decay among diverse lignicolous fungi. Mycologia 1997, 89, 199–219. [Google Scholar] [CrossRef]
- Boddy, L. Fungal community ecology and wood decomposition process in angiosperms: From standing tree to complete decay of coarse woody debris. Ecol. Bull. 2001, 49, 43–56. [Google Scholar]
- Dix, N.J.; Webster, J. Fungal Ecology, 1st ed.; Springer: Dordrecht, The Netherlands, 1995; ISBN 978-94-010-4299-4. [Google Scholar]
- Mašínová, T.; Bahnmann, B.D.; Větrovský, T.; Tomšovský, M.; Merunková, K.; Baldrian, P. Drivers of yeast community composition in the litter and soil of a temperate forest. FEMS Microbiol. Ecol. 2017, 93, fiw223. [Google Scholar] [CrossRef] [PubMed]
- Richter, C.; Yurkov, A.M.; Boekhout, T.; Stadler, M. Diversity of Tilletiopsis-Like Fungi in Exobasidiomycetes (Ustilaginomycotina) and Description of Six Novel Species. Front. Microbiol. 2019, 10, 2544. [Google Scholar] [CrossRef] [PubMed]
- Grad, B.; Kowalski, T.; Kraj, W. Studies on secondary metabolite produced by Chalara fraxinea and its phytotoxic influence on Fraxinus excelsior. Phytopathologia 2009, 54, 61–69. [Google Scholar]
- Andersson, P.F.; Bengtsson, S.; Stenlid, J.; Broberg, A. B-norsteroids from Hymenoscyphus pseudoalbidus. Molecules 2012, 17, 7769–7781. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.L.H.; Zaspel, I.; Schuemann, M.; Stephanowitz, H.; Krause, E. Rapid In-Vitro and In-Vivo Detection of Chalara fraxinea by Means of Mass Spectrometric Techniques. Am. J. Plant Sci. 2013, 4, 444–453. [Google Scholar] [CrossRef] [Green Version]
- Andersson, P.F.; Johansson, S.B.K.; Stenlid, J.; Broberg, A. Isolation, identification and necrotic activity of viridiol from Chalara fraxinea, the fungus responsible for dieback of ash. For. Pathol. 2010, 40, 43–46. [Google Scholar] [CrossRef]
- Citron, C.A.; Junker, C.; Schulz, B.; Dickschat, J.S. A volatile lactone of Hymenoscyphus pseudoalbidus, pathogen of European ash dieback, inhibits host germination. Angew. Chem. Int. Ed. 2014, 53, 4346–4349. [Google Scholar] [CrossRef]
- Kowalski, T.; Holdenrieder, O. Pathogenicity of Chalara fraxinea. For. Pathol. 2009, 39, 1–7. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef] [Green Version]
- Frankland, J.C. Mechanisms in fungal succession. In The Fungal Community: Its Organization and Role in the Ecosystem; Carroll, G.C., Wicklow, D.T., Eds.; Marcel Dekker: New York, NY, USA, 1992; pp. 383–401. ISBN 978-0824786052. [Google Scholar]
- Becker, R.; Ulrich, K.; Behrendt, U.; Kube, M.; Ulrich, A. Analyzing ash leaf-colonizing fungal communities for their biological control of Hymenoscyphus fraxineus. Front. Microbiol. 2020, 11, 590944. [Google Scholar] [CrossRef]
- Timpe, D. Introduction to the theory of science. Philos. Hist. 1981, 14, 37–39. [Google Scholar] [CrossRef]
- Mahajan, C.; Basotra, N.; Singh, S.; Di Falco, M.; Tsang, A.; Chadha, B.S. Malbranchea cinnamomea: A thermophilic fungal source of catalytically efficient lignocellulolytic glycosyl hydrolases and metal dependent enzymes. Bioresour. Technol. 2016, 200, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, L.M.O.; Vici, A.C.; Pinheiro, M.P.; Heinen, P.R.; de Oliveira, A.H.C.; Ward, R.J.; Prade, R.A.; Buckeridge, M.S.; de Lourdes Teixeira de Moraes Polizeli, M. A highly glucose tolerant ß-glucosidase from Malbranchea pulchella (MpBg3) enables cellulose saccharification. Sci. Rep. 2020, 10, 6998. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Sato, G.; Toriyabe, A.; Kudo, H.; Hashimoto, A.; Matsumura, M.; Harada, Y.; Kurihara, Y.; et al. Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 2015, 82, 75–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, M.; Yawata, K.; Orino, Y.; Ueda, S.; Isogai, Y.; Taguchi, G.; Shimosaka, M.; Hashimoto, S. Agrobacterium tumefaciens-mediated transformation of antifungal lipopeptide producing fungus Coleophoma empetri F-11899. Curr. Genet. 2009, 55, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.E.; Finlay, R.; Ward, J.S. Antifungal compounds produced by Epicoccum purpurascens against soil-borne plant pathogenic fungi. Soil Biol. Biochem. 1987, 19, 657–664. [Google Scholar] [CrossRef]
- Bozoudi, D.; Tsaltas, D. The multiple and versatile roles of Aureobasidium pullulans in the vitivinicultural sector. Fermentation 2018, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- Asgari, B.; Zare, R.; Gams, W. Coniochaeta ershadii, a new species from Iran, and a key to well-documented Coniochaeta species. Nov. Hedwig. 2007, 84, 175–187. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.J.; Cadelis, M.M.; Kim, S.H.; Swift, S.; Copp, B.R.; Villas-Boas, S.G. Epipyrone a, a broad-spectrum antifungal compound produced by Epicoccum nigrum ICMP 19927. Molecules 2020, 25, 5997. [Google Scholar] [CrossRef]
- Zhu, J.; Xiong, P.; Li, Z.; Li, J.; Lin, L.; Fu, X.; Huang, Y.; Xiong, Y.; Li, C. Antifungal sesquiterpenes with post-harvest anthracnose control effect on bananas from the fungus Fusarium lateritium. Nat. Prod. Res. 2021, 15, 1–8. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Stark, R.E.; Casadevall, A. Fungal Melanin: What do we know about structure? Front. Microbiol. 2015, 6, 1463. [Google Scholar] [CrossRef] [Green Version]
- Pukalski, J.; Marcol, N.; Wolan, N.; Płonka, P.M.; Ryszka, P.; Kowalski, T.; Latowski, D. Detection of a pheomelanin-like pigment by EPR spectroscopy in the mycelium of Plenodomus biglobosus. Acta Biochim. Pol. 2020, 67, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Barnett, H.L.; Lilly, V.G. A destructive mycoparasite, Gliocladium roseum. Mycologia 1962, 54, 72–77. [Google Scholar] [CrossRef]
- Pachenari, A.; Dix, N.J. Production of toxins and wall degrading enzymes by Gliocladium roseum. Trans. Br. Mycol. Soc. 1980, 74, 561–566. [Google Scholar] [CrossRef]
- Bailey, B.A.; Bae, H.; Strem, M.D.; Crozier, J.; Thomas, S.E.; Samuels, G.J.; Vinyard, B.T.; Holmes, K.A. Antibiosis, mycoparasitism, and colonization success for endophytic Trichoderma isolates with biological control potential in Theobroma cacao. Biol. Control 2008, 46, 24–35. [Google Scholar] [CrossRef]
- John, R.P.; Tyagi, R.D.; Prévost, D.; Brar, S.K.; Pouleur, S.; Surampalli, R.Y. Mycoparasitic Trichoderma viride as a biocontrol agent against Fusarium oxysporum f. sp. adzuki and Pythium arrhenomanes and as a growth promoter of soybean. Crop Prot. 2010, 29, 1452–1459. [Google Scholar] [CrossRef]
- Boddy, L. Interspecific combative interactions between wood-decaying basidiomycetes. FEMS Microbiol. Ecol. 2000, 31, 185–194. [Google Scholar] [CrossRef]
- Rayner, A.D.M.; Boddy, L. Fungal Decomposition of Wood: Its Biology and Ecology, 1st ed.; Wiley: Chichester, UK; New York, NY, USA, 1988; ISBN 0471103101. [Google Scholar]
- Sutton, B.C.; Gibson, I.A. Pezizella oenotherae (conidial state Hainesia lythri). In CMI Descriptions of Pathogenic Fungi and Bacteria; Commonwealth Mycological Institute: Kew, UK, 1977; pp. 1–2. [Google Scholar]
- Debode, J.; Van Hemelrijck, W.; Heungens, K.; Maes, M.; Creemers, P. First report of Pilidium concavum causing tan-brown rot on strawberry fruit in Belgium. Plant Dis. 2011, 95, 1029. [Google Scholar] [CrossRef]
- Wunsch, M.J.; Bergstrom, G.C. Genetic and morphological evidence that Phoma sclerotioides, causal agent of brown root rot of alfalfa, is composed of a species complex. Phytopathology 2011, 101, 594–610. [Google Scholar] [CrossRef] [Green Version]
- Neubauer, C.; Heitmann, B.; Vogel, C. Morphology, vegetative compatibility and pathogenicity of Verticillium dahliae isolates from woody ornamentals in Germany. J. Plant Dis. Prot. 2009, 116, 109–114. [Google Scholar] [CrossRef]
- Pautasso, M.; Aas, G.; Queloz, V.; Holdenrieder, O. European ash (Fraxinus excelsior) dieback—A conservation biology challenge. Biol. Conserv. 2013, 158, 37–49. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Research Plots | Brody | Dynów | Miechów | Myślenice | Ojców |
|---|---|---|---|---|---|
| Coordinates | 49°51′34″ N | 49°47′17″ N | 50°13′02″ N | 49°56′45″ N | 50°11′59″ N |
| 19°40′32″ E | 22°15′20″ E | 20°02′09″ E | 19°52′48″ E | 19°48′43″ E | |
| Altitude [m a.s.l.] | 369 | 417 | 281 | 356 | 446 |
| Fraxinus excelsior: | |||||
| age | 54 | 60 | 27 | 82 | 63 |
| Share (%) | 30 | 10 | 10 | 50 | 80 |
| Other tree species present 1 | Ag, Ap, Cb, Qr, Tc | Bp, Cb, Fs, Ps, Qr | Ap, Fs, Ld, Pt, Qr | Ap, Ag, Ld, Qr, Tc | Ap, Bp, Fs, Ps, Qr |
| Habitat type | Fresh upland broadleaved forest | Fresh upland broadleaved forest | Fresh upland broadleaved forest | Fresh upland broadleaved forest | Fresh upland broadleaved forest |
| Sampling date | 10 May 2017 | 16 May 2017 | 08 June 2017 | 23 May 2017 | 30 May 2017 |
| Number of petioles (petiole sections) used in fungal isolations | 60 (720) | 60 (720) | 60 (720) | 60 (720) | 60 (720) |
| Number of petioles (petiole sections) yielding colonies of fungi | 60 (419) | 60 (582) | 60 (465) | 60 (487) | 60 (548) |
| Taxon | Phylum 1 | Order | Number of Colonized Ash Petioles in Study Plots 2 | |||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | Total Number (%) | |||
| Agaricomycetes sp. | B | - | 4 | 5 | 9 (3.0) | |||
| Alternaria sp. 1 | A | Pleosporales | 11 a 3 | 16 a | 9 a | 20 a | 11 a | 67 (22.3) 4 |
| Alternaria sp. 2 | A | Pleosporales | 1 | 1 (0.3) | ||||
| Alternaria sp. 3 | A | Pleosporales | 2 | 7 | 9 (3.0) | |||
| Alternaria sp. 4 | A | Pleosporales | 1 | 1 (0.3) | ||||
| Athelia sp. | B | Atheliales | 1 | 1 (0.3) | ||||
| Aureobasidium pullulans | A | Dothideales | 3 | 2 | 1 | 6 (2.0) | ||
| Boeremia exigua | A | Pleosporales | 1 | 1 | 3 | 2 | 7 (2.3) | |
| Boeremia sp. | A | Pleosporales | 4 bc | 13 ab | 26 a | 17 a | 2 c | 62 (20.7) 4 |
| Chaetomium globosum | A | Sordariales | 2 | 2 (0.7) | ||||
| Chromelosporium sp. | A | Pezizales | 2 | 1 | 1 | 1 | 5 (1.7) | |
| Cladosporium sp. 1 | A | Capnodiales | 2 | 2 | 1 | 5 (1.7) | ||
| Cladosporium sp. 2 | A | Capnodiales | 3 | 1 | 6 | 1 | 1 | 12 (4.0) 4 |
| Clonostachys rosea | A | Hypocreales | 2 | 2 | 1 | 5 (1.7) | ||
| Colletotrichum acutatum | A | Glomerellales | 4 | 2 | 4 | 4 | 14 (4.7) 4 | |
| Coniochaeta angustispora | A | Coniochaetales | 1 | 1 (0.3) | ||||
| Coniochaeta sp. 1 | A | Coniochaetales | 1 | 2 | 3 | 2 | 8 (2.7) | |
| Coniochaeta sp. 2 | A | Coniochaetales | 2 | 2 (0.7) | ||||
| Coniochaeta sp. 3 | A | Coniochaetales | 6 | 2 | 2 | 3 | 13 (4.3) 4 | |
| Coniothyrium cf. dispersellum | A | Pleosporales | 7 | 7 (2.3) | ||||
| Coprinellus disseminatus | B | Agaricales | 1 | 2 | 3 (1.0) | |||
| Coprinellus micaceus | B | Agaricales | 1 | 1 (0.3) | ||||
| Cyathicula coronata | A | Helotiales | 1 | 1 | 1 | 2 | 5 (1.7) | |
| Cyathicula fraxinophila | A | Helotiales | 1 | 2 | 3 | 2 | 1 | 9 (3.0) |
| Cyathicula sp. 2 | A | Helotiales | 1 | 1 | 1 | 3 (1.0) | ||
| Cytospora pruinosa | A | Diaporthales | 2 | 1 | 2 | 5 (1.7) | ||
| Cytospora sp. | A | Diaporthales | 2 | 1 | 3 (1.0) | |||
| Dactylaria sp. | A | Rhytismatales | 1 | 1 (0.3) | ||||
| Desmazierella acicola | A | Pezizales | 7 | 2 | 1 | 1 | 3 | 14 (4.7) 4 |
| Diaporthe eres | A | Diaporthales | 2 | 1 | 1 | 16 | 4 | 24 (8.0) 4 |
| Discohainesia oenotherae | A | Chaetomellales | 2 | 2 (0.7) | ||||
| Epicoccum nigrum | A | Pleosporales | 8 ab | 1 b | 9 ab | 20 a | 4 b | 42 (14.0) 4 |
| Eupenicillium sp. | A | Eurotiales | 1 | 1 (0.3) | ||||
| Fellozyma sp. | B | Incertae sedis | 1 | 1 (0.3) | ||||
| Fusariella sp. | A | Incertae sedis | 1 | 1 | 1 | 3 (1.0) | ||
| Fusarium lateritium | A | Hypocreales | 10 a | 4 a | 2 a | 12 a | 5 a | 33 (11.0) 4 |
| Fusarium sp. | A | Hypocreales | 2 b | 14 a | 3 ab | 11 ab | 5 ab | 35 (11.7) 4 |
| Graphilbum sp. | A | Ophiostomatales | 3 | 1 | 4 (1.3) | |||
| Gyrothrix sp. | A | Incertae sedis | 1 | 1 | 2 (0.7) | |||
| Helotiales sp. 1 | A | Helotiales | 26 a | 0 c | 3 bc | 6 bc | 14 ab | 49 (16.3) 4 |
| Herpotrichia sp. | A | Pleosporales | 2 | 2 (0.7) | ||||
| Hymenoscyphus caudatus | A | Helotiales | 1 | 5 | 1 | 1 | 8 (3.0) | |
| Hymenoscyphus fraxineus | A | Helotiales | 14 b | 37 a | 34 a | 15 b | 10 b | 110 (36.7) 4 |
| Hymenoscyphus scutula | A | Helotiales | 2 | 1 | 2 | 1 | 6 (2.0) | |
| Hymenoscyphus sp. | A | Helotiales | 2 | 1 | 1 | 4 (1.3) | ||
| Hypholoma fasciculare | B | Agaricales | 1 | 1 (0.3) | ||||
| Hypoderma rubi | A | Rhytismatales | 2 | 5 | 1 | 9 | 6 | 23 (7.7) 4 |
| Lanzia sp. | A | Helotiales | 2 | 1 | 3 (1.0) | |||
| Lasiosphaeriaceae sp. | A | Sordariales | 2 | 3 | 5 (1.7) | |||
| Lemonniera sp. | A | Helotiales | 2 | 2 (0.7) | ||||
| Leptosphaeria conoidea | A | Pleosporales | 1 | 1 (0.3) | ||||
| Leptosphaeria rubefaciens | A | Pleosporales | 4 | 1 | 1 | 6 (2.0) | ||
| Leptosphaeria urticae | A | Pleosporales | 1 | 1 | 1 | 3 (1.0) | ||
| Leptosphaeria sp. 1 | A | Pleosporales | 1 | 1 (0.3) | ||||
| Leptosphaeria sp. 2 | A | Pleosporales | 1 | 1 | 1 | 1 | 4 (1.3) | |
| Leptospora rubella | A | Incertae sedis | 3 | 1 | 4 (1.3) | |||
| Leptospora sp. | A | Incertae sedis | 1 | 1 (0.3) | ||||
| Lophiostoma corticola | A | Pleosporales | 1 | 3 | 4 (1.3) | |||
| Malbranchea sp. | A | Onygenales | 1 | 1 (0.3) | ||||
| Microsphaeropsis sp. | A | Pleosporales | 12 | 4 | 3 | 19 (6.3) 4 | ||
| Mortierella sp. | M | Mortierellales | 2 | 2 (0.7) | ||||
| Mycena citrinomarginata | B | Agaricales | 1 | 1 | 2 (0.7) | |||
| Mycena sp. | B | Agaricales | 1 | 1 | 2 (0.7) | |||
| Mycoarthris sp. 1 | A | Rhytismatales | 1 | 5 | 6 | 12 (4.0) 4 | ||
| Mycoarthris sp. 2 | A | Rhytismatales | 2 | 2 (0.7) | ||||
| Nemania diffusa | A | Xylariales | 0 b | 12 a | 16 a | 0 b | 5 ab | 33 (11.0) 4 |
| Nemania serpens | A | Xylariales | 38 b | 13 c | 25 bc | 34 b | 54 a | 164 (54.7) 4 |
| Nemania sp. | A | Xylariales | 1 | 3 | 4 (1.3) | |||
| Neofabraea kienholzii | A | Helotiales | 2 | 3 | 5 (1.7) | |||
| Neonectria sp. 1 | A | Hypocreales | 4 | 4 (1.3) | ||||
| Neonectria sp. 2 | A | Hypocreales | 2 | 6 | 1 | 9 (3.0) | ||
| Paracucurbitaria corni | A | Pleosporales | 2 | 2 | 4 (1.3) | |||
| Paraophiobolus arundinis | A | Pleosporales | 1 | 1 (0.3) | ||||
| Paraphaeosphaeria michotii | A | Pleosporales | 1 | 1 (0.3) | ||||
| Paraphaeosphaeria neglecta | A | Pleosporales | 9 | 4 | 13 (4.3) 4 | |||
| Peniophora incarnata | B | Russulales | 6 | 1 | 3 | 3 | 2 | 15 (5.0) 4 |
| Periconia sp. 1 | A | Pleosporales | 1 | 1 | 2 | 4 (1.3) | ||
| Periconia sp. 2 | A | Pleosporales | 1 | 1 | 2 (0.7) | |||
| Peziza ninguis | A | Pezizales | 5 | 1 | 6 (2.0) | |||
| Peziza varia | A | Pezizales | 8 | 7 | 5 | 3 | 6 | 29 (9.7) 4 |
| Pezizomycetes sp. | A | - | 1 | 1 (0.3) | ||||
| Phacidium lacerum | A | Phacidiales | 1 | 1 | 2 (0.7) | |||
| Phlyctema vagabunda | A | Helotiales | 2 | 1 | 3 (1.0) | |||
| Phoma sp. 1 | A | Pleosporales | 3 | 3 (1.0) | ||||
| Phoma sp. 2 | A | Pleosporales | 2 | 2 | 4 (1.3) | |||
| Phoma sp. 3 | A | Pleosporales | 1 | 17 | 2 | 3 | 23 (7.7) 4 | |
| Phoma sp. 4 | A | Pleosporales | 1 | 2 | 3 | 5 | 11 (3.7) 4 | |
| Phoma sp. 5 | A | Pleosporales | 5 | 6 | 2 | 13 (4.3) 4 | ||
| Plectosphaerella cucumerina | A | Glomerellales | 1 | 3 | 4 (1.3) | |||
| Plenodomus sp. 1 | A | Pleosporales | 1 | 1 | 2 (0.7) | |||
| Pleospora sp. | A | Pleosporales | 1 | 3 | 4 (1.3) | |||
| Psathyrella piluliformis | B | Agaricales | 1 | 1 (0.3) | ||||
| Pseudocoleophoma polygonicola | A | Pleosporales | 1 | 1 | 1 | 3 (1.0) | ||
| Pseudoophiobolus italicus | A | Pleosporales | 4 | 4 (1.3) | ||||
| Pseudoplectania nigrella | A | Pezizales | 2 | 1 | 3 (1.0) | |||
| Pyrenochaeta sp. | A | Incertae sedis | 1 | 1 | 2 (0.7) | |||
| Pyrenochaetopsis leptospora | A | Pleosporales | 2 | 2 (0.7) | ||||
| Pyrenopeziza petiolaris | A | Helotiales | 1 | 10 | 2 | 6 | 3 | 22 (7.3) 4 |
| Pyrenopeziza sp. | A | Helotiales | 1 | 1 | 2 (0.7) | |||
| Rhexocercosporidium sp. | A | Helotiales | 3 | 4 | 5 | 4 | 5 | 21 (7.0) 4 |
| Rosellinia abscondita | A | Xylariales | 3 | 2 | 1 | 1 | 7 (2.3) | |
| Rosellinia corticium | A | Xylariales | 1 | 1 | 2 (0.7) | |||
| Rosellinia nectrioides | A | Xylariales | 1 | 1 | 2 (0.7) | |||
| Sistotrema sp. | B | Cantharellales | 1 | 1 (0.3) | ||||
| Tetracladium sp. | A | Rhytismatales | 2 | 1 | 3 (1.0) | |||
| Tilletiopsis washingtonensis | B | Entylomatales | 2 | 2 (0.7) | ||||
| Trametes versicolor | B | Polyporales | 2 | 2 (0.7) | ||||
| Trichoderma viride | A | Hypocreales | 1 | 3 | 4 (1.3) | |||
| Truncatella sp. | A | Amphisphaeriales | 4 | 4 (1.3) | ||||
| Typhula sp. 1 | B | Agaricales | 2 | 1 | 1 | 4 (1.3) | ||
| Typhula sp. 2 | B | Agaricales | 3 a | 10 a | 3 a | 8 a | 9 a | 33 (11.0) 4 |
| Umbelopsis sp. | M | Umbelopsidales | 1 | 1 | 1 | 3 (1.0) | ||
| Vargamyces aquaticus | A | Incertae sedis | 1 | 1 (0.3) | ||||
| Venturia fraxini | A | Venturiales | 9 a | 4 a | 11 a | 7 a | 10 a | 41 (13.7) 4 |
| Verticillium dahliae | A | Glomerellales | 1 | 1 (0.3) | ||||
| Xylaria polymorpha | A | Xylariales | 6 | 1 | 1 | 8 (2.7) | ||
| Xylaria sp. 1 | A | Xylariales | 1 | 3 | 4 (1.3) | |||
| Number of petioles analyzed | - | - | 60 | 60 | 60 | 60 | 60 | 300 |
| Number of colonies obtained | - | - | 446 | 696 | 519 | 576 | 595 | 2832 |
| Species richness (S) | - | - | 53 | 70 | 58 | 59 | 58 | 117 |
| Sum of frequency of taxa | - | - | 218 | 272 | 249 | 274 | 238 | 1251 |
| Simpson’s index of diversity (SID, SID = 1 − D) | - | - | 0.94 | 0.96 | 0.94 | 0.95 | 0.93 | 0.96 |
| Shannon index of diversity (H′) | - | - | 3.31 | 3.67 | 3.36 | 3.49 | 3.40 | 3.86 |
| Taxon | Order 1 | Phylum 2 | Taxa No. | Number of Petioles Colonized in Study Plots 3 | Total Number (%) | Hf 6 | Accession | BLASTn | Identities % | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |||||||||
| Amniculicola sp. | P | A | FeF402 | 2 | 2 (0.3) | MZ492963 | MK353143 | 92.42 | |||||
| Anthostoma turgidum | X | A | FeF425 | 2 | 2 (0.3) | MZ493125 | KC774617 | 99.84 | |||||
| Apiognomonia hystrix | D | A | FeF401 | 1 | 1 (0.2) | MZ493111 | KX776442 | 100 | |||||
| Boeremia exigua | P | A | FeF403 | 2 | 6 | 8 (1.3) | 1 | MZ493112 | MN540289 | 100 | |||
| Calycina sp. | H | A | FeF404 | 1 | 1 | 2 (0.3) | MZ493113 | JF908571 | 100 | ||||
| Cyathicula coronata | H | A | FeF405 | 1 | 16 | 7 | 3 | 27 (4.5) 5 | 16 | MZ492976 | MH858141 | 100 | |
| Cyathicula fraxinophila | H | A | FeF406 | 21 a 4 | 11 a | 18 a | 12 a | 20 a | 82 (13.7) 5 | 31 | MZ492980 | MK584998 | 99.09 |
| Cyathicula sp. 1 | H | A | FeF407 | 3 | 3 (0.5) | MZ492983 | MK584998 | 98.7 | |||||
| Cyathicula sp. 2 | H | A | FeF408 | 2 | 1 | 2 | 5 (0.8) | MZ492978 | MK584998 | 98.7 | |||
| Cyathicula sp. 3 | H | A | FeF409 | 6 | 1 | 6 | 2 | 15 (2.5) | 5 | MZ493114 | MK584998 | 98.35 | |
| Cyathicula sp. 4 | H | A | FeF445 | 1 | 2 | 3 (0.5) | MZ492982 | MK584998 | 99.09 | ||||
| Diaporthe sp. 1 | D | A | FeF446 | 1 | 1 (0.2) | 1 | MZ493057 | EU552122 | 99.67 | ||||
| Diplodia fraxini | B | A | FeF410 | 1 | 1 | 2 (0.3) | 2 | MZ493115 | MT587347 | 100 | |||
| Helotiales sp. 1 | H | A | FeF413 | 2 | 1 | 4 | 7 (1.2) | MZ492996 | GU174285 | 98.97 | |||
| Helotiales sp. 2 | H | A | FeF414 | 1 | 1 (0.2) | MZ492997 | MH578458 | 98.41 | |||||
| Helotiales sp. 3 | H | A | FeF415 | 2 | 2 | 1 | 2 | 7 (1.2) | 1 | MZ493124 | KY430513 | 99.44 | |
| Hymenoscyphus caudatus | H | A | FeF416 | 3 | 4 | 5 | 4 | 5 | 21 (3.5) 5 | 7 | MZ492985 | KM114539 | 99.38 |
| Hymenoscyphus fraxineus | H | A | FeF417 | 30 bc | 59 ab | 66 a | 21 c | 26 c | 202 (33.7) 5 | MZ492992 | LLCC00000000 | 100 | |
| Hymenoscyphus scutula | H | A | FeF418 | 8 | 2 | 4 | 6 | 7 | 27 (4.5) | 12 | MZ492989 | MK674606 | 99.83 |
| Hymenoscyphus sp. | H | A | FeF419 | 1 | 1 | 2 | 4 (0.7) | MZ492987 | MK674606 | 97.03 | |||
| Hypoderma rubi | R | A | FeF420 | 8 | 4 | 3 | 13 | 4 | 32 (5.3) 5 | 13 | MZ493025 | GU138741 | 99.64 |
| Lasionectria sp. | Hy | A | FeF421 | 1 | 1 (0.2) | MZ493040 | KY607542 | 96.47 | |||||
| Leptosphaeria sclerotioides | P | A | FeF422 | 7 | 14 | 2 | 23 (3.8) 5 | 5 | MZ492959 | FJ179155 | 100 | ||
| Leptosphaeria sp. 2 | P | A | FeF423 | 29 ab | 17 ab | 17 ab | 38 a | 11 b | 112 (18.7) 5 | 39 | MZ492957 | KC965764 | 95.33 |
| Leptospora rubella | Is | A | FeF424 | 2 | 2 | 1 | 1 | 6 (1.0) | 5 | MZ493116 | MH857603 | 99.83 | |
| Lophiostoma corticola | P | A | FeF426 | 2 | 1 | 1 | 4 (0.7) | 2 | MZ493117 | KU712227 | 100 | ||
| Marasmius epiphyllus | A | B | FeF427 | 1 | 1 (0.2) | MZ493079 | FN293008 | 99.66 | |||||
| Neonectria sp. 2 | Hy | A | FeF411 | 2 | 1 | 2 | 5 (0.8) | 1 | MZ493037 | JF735313 | 99.82 | ||
| Paraophiobolus arundinis | P | A | FeF428 | 2 | 2 | 5 | 9 (1.5) | 6 | MZ492953 | MG520945 | 100 | ||
| Phacidium lacerum | Ph | A | FeF429 | 2 | 1 | 1 | 4 (0.7) | 3 | MZ493005 | KU942438 | 100 | ||
| Plenodomus sp. 2 | P | A | FeF430 | 1 | 1 (0.2) | MZ492947 | KU973715 | 100 | |||||
| Pleosporales sp. | P | A | FeF431 | 1 | 1 (0.2) | MZ492961 | KC965786 | 96.51 | |||||
| Pseudocoleophoma polygonicola | P | A | FeF432 | 1 | 1 (0.2) | MZ492973 | NR_154274 | 99.62 | |||||
| Pyrenochaeta sp. | Is | A | FeF433 | 11 | 2 | 4 | 2 | 19 (3.2) | 2 | MZ492948 | MT236853 | 98.98 | |
| Pyrenopeziza petiolaris | H | A | FeF434 | 7 | 10 | 3 | 11 | 31 (5.2) 5 | 12 | MZ493015 | MH857804 | 99.29 | |
| Trichopeziza sp. 1 | H | A | FeF435 | 8 | 1 | 1 | 1 | 11 (1.8) | 1 | MZ493009 | AB481285 | 97.93 | |
| Trichopeziza sp. 2 | H | A | FeF436 | 1 | 1 | 1 | 3 (0.5) | MZ493008 | JN033398 | 99.23 | |||
| Typhula erythropus | A | B | FeF439 | 1 | 1 | 1 | 3 (0.5) | 2 | MZ493118 | MT232359 | 100 | ||
| Typhula sp. 1 | A | B | FeF437 | 1 | 3 | 1 | 6 | 4 | 15 (2.5) | 3 | MZ493119 | KR673716 | 99.86 |
| Typhula sp. 2 | A | B | FeF438 | 10 | 2 | 15 | 10 | 37 (6.7) 5 | 16 | MZ493081 | NR_132792 | 88.65 | |
| Venturia fraxini | V | A | FeF440 | 26 a | 24 a | 27 a | 27 a | 29 a | 133 (22.2) 5 | 53 | MZ493120 | EU035457 | 99.83 |
| Xylaria sp. 1 | X | A | FeF441 | 2 | 2 (0.3) | MZ493122 | MF774330 | 99.83 | |||||
| Xylaria sp. 2 | X | A | FeF442 | 1 | 1 | 2 (0.3) | 1 | MZ493121 | MF774330 | 99.48 | |||
| Xylaria sp. 3 | X | A | FeF443 | 1 | 2 | 3 (0.3) | 1 | MZ493059 | MH578518 | 97.86 | |||
| Xylaria sp. 4 | X | A | FeF444 | 1 | 1 (0.2) | MZ493123 | MH578518 | 97.1 | |||||
| Number of petioles analyzed | - | - | - | 120 | 120 | 120 | 120 | 120 | 600 | - | - | - | - |
| Species richness (S) | - | - | - | 25 | 24 | 17 | 30 | 31 | 45 | - | - | - | - |
| Sum of frequency of taxa | - | - | - | 180 | 171 | 171 | 194 | 166 | 882 | 241 | - | - | - |
| Simpson’s index of diversity (SID, SID=1-D) | - | - | - | 0.9 | 0.84 | 0.79 | 0.91 | 0.91 | 0.89 | - | - | - | - |
| Shannon index of diversity (H′) | - | - | - | 2.61 | 2.36 | 1.99 | 2.72 | 2.82 | 2.73 | - | - | - | - |
| Interaction Types | Hymenoscyphus fraxineus (Strain) Number (%) | ||
|---|---|---|---|
| Hf 1 2 | Hf 2 2 | Total 3 | |
| A (mutual direct contact of mycelia) | 73 (44.5) a | 69 (42.1) a | 142 (43.3) a |
| B (inhibition zone) 1 | 75 (45.7) a | 79 (48.2) a | 154 (46.9) a |
| C (overgrowth of H. fraxineus colony by a saprotroph) | 16 (9.8) a | 16 (9.8) a | 32 (9.8) b |
| D (overgrowth of saprotroph’s colony by H. fraxineus) | 0 | 0 | 0 c |
| Number of pathogen/co-partner combinations in dual cultures | 164 | 164 | 328 |
| 1 inhibition zone width | |||
| Bs (up to 3 mm) | 29 a | 31 a | 60 (18.3) a |
| Bm (4–5 mm) | 25 a | 26 a | 51 (15.5) ab |
| Bw (6–8 mm) | 10 a | 12 a | 22 (6.7) b |
| Bv (>8 mm) | 11 a | 10 a | 21 (6.4) b |
| Reduction Rate | Hymenoscyphus fraxineus | Fungal Co-Partners | ||||
|---|---|---|---|---|---|---|
| Hf 1 1 | Hf 2 1 | Total Number (%) 2 | Hf 1 1 | Hf 2 1 | Total Number (%) 2 | |
| a (<25%) | 5 a | 5 a | 10 (3.0) b | 37 a | 46 a | 83 (25.3) b |
| b (26–50%) | 68 a | 57 a | 125 (38.1) a | 104 a | 97 a | 201 (61.3) a |
| c (51–75%) | 85 a | 93 a | 178 (54.3) a | 5 a | 6 a | 11 (3.4) c |
| d (>75%) | 6 a | 9 a | 15 (4.6) b | 0 | 0 | 0 c |
| f (0%) | 0 | 0 | 0 (0.0) b | 18 a | 15 a | 33 (10.0) bc |
| Total | 164 | 164 | 328 (100.0) | 164 | 164 | 328 (100.0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalski, T.; Bilański, P. Fungi Detected in the Previous Year’s Leaf Petioles of Fraxinus excelsior and Their Antagonistic Potential against Hymenoscyphus fraxineus. Forests 2021, 12, 1412. https://doi.org/10.3390/f12101412
Kowalski T, Bilański P. Fungi Detected in the Previous Year’s Leaf Petioles of Fraxinus excelsior and Their Antagonistic Potential against Hymenoscyphus fraxineus. Forests. 2021; 12(10):1412. https://doi.org/10.3390/f12101412
Chicago/Turabian StyleKowalski, Tadeusz, and Piotr Bilański. 2021. "Fungi Detected in the Previous Year’s Leaf Petioles of Fraxinus excelsior and Their Antagonistic Potential against Hymenoscyphus fraxineus" Forests 12, no. 10: 1412. https://doi.org/10.3390/f12101412
APA StyleKowalski, T., & Bilański, P. (2021). Fungi Detected in the Previous Year’s Leaf Petioles of Fraxinus excelsior and Their Antagonistic Potential against Hymenoscyphus fraxineus. Forests, 12(10), 1412. https://doi.org/10.3390/f12101412