1. Introduction
Forest resources and Non-Timber Forest Products (NTFP) species, in particular, face multiple anthropogenic pressures that include direct threats, due to their exploitation, and indirect ones, through habitat threats [
1,
2]. Such pressures include overexploitation, changing land use, bushfires, grazing and invasive species [
2,
3,
4], which can negatively impact natural populations at both the species and community scales [
5,
6]. Such impacts may be irreversible for some species [
7]. In this context, it is important to analyse the current status of NTFP populations in relation to anthropogenic pressures to understand and potentially mitigate the impacts.
Synergistically with human disturbance, environmental factors (such as climate, soil conditions and slope) influence the abundance and observed variability in the functional characteristics of tree species in natural habitats [
8,
9]. Plant responses to diverse environmental stresses often manifest in their morphological and physiological characteristics [
10], which determine their growth, survival and reproduction. Thus, variations in environmental factors typically lead to variations in NTFP population density, vigour and structure. Therefore, understanding ecological correlates and stresses, along with anthropogenic disturbances, is also an important step in the planning of management actions for important NTFPs. This can help to predict the potential for recovery of species populations if pressures are mitigated or removed.
In the face of these different pressures, a comprehensive knowledge of species autecology is needed to understand population characteristics and dynamics due to the different pressures. This is necessary for guiding sustainable conservation strategies [
11,
12]. In terms of species autecology, population structure (density and size class distribution) and plant morphology are a direct reflection of their health and performance [
11]. Thus, the knowledge of population structure, along with plant species’ morphologies, is an essential tool in understanding their population ecology and trends in relation to pressures [
8].
Mimusops andongensis Hiern and
Mimusops kummel Bruce ex A. DC are two NTFP tree species naturally occurring in many African countries, including Benin, where they occur mainly in semi-deciduous and riparian forests, respectively. Both species are used for energy, construction and manufacturing, as well as for alimentary and medicinal purposes [
13,
14,
15,
16,
17]. Additionally,
M.
kummel has been identified as one of the important sources of pollen for honey production in different African countries, although among the least abundant species [
18,
19]. Thus,
M.
andongensis and
M.
kummel are two important NTFP species that deserve more study [
13,
20]. Mirroring many countries worldwide, forest resources in Benin face various pressures, including overharvesting, deforestation for cultivation, uncontrolled bushfires, invasive species, etc. [
21].
M.
andongensis and
M.
kummel are subject to such pressures [
15] and therefore understanding variation in their population structure and morphology in the context of anthropogenic and ecological stresses will inform conservation options.
Thus, this study assessed (i) the population structure and morphology of M. andongensis and M. kummel within different bioclimatic zones in Benin, (ii) how ecological factors influence structural and morphological characteristics of both species, and (iii) how anthropogenic disturbances on the species and their habitats shape their population structure.
4. Discussion
Population structure and morphology were different between
M.
andongensis and
M.
kummel. The density of trees with dbh ≥ 5cm, regeneration stems, and mean height of trees were higher for
M.
andongensis than
M.
kummel. The first explanation for this difference may be attributed to them being two different species that may have specific functional traits and thus different performance in response to ecological conditions and competition [
31,
32]. However, habitat is an important factor which might also account for the difference between the two species [
33]. For instance, the two species occur under different climatic conditions with
M.
andongensis in the most humid part of the country and
M.
kummel in the sub-humid and semi-arid parts [
22]. We also found a significant difference between the forests based on soil characteristics, which may explain the differences between the two species. Indeed, soils in Lama Forest reserve have the highest clay (47% with 33% of sand and 20% of silt), organic carbon content and CEC in comparison to other forests, indicating that this forest is more fertile and more suitable for growth and development [
34,
35]. Fertile soils reflect a higher capacity to hold nutrients due to higher exchangeable base cations and higher organic matter content with a healthy pH [
34]. However, other factors, including soil structure, climate and parent material, as well as physiochemical and biological properties, can influence soil fertility [
36]. Also, tree morphology and the abundance of a species are a result of its performance in response to changes in ecological conditions through time [
10].
Considering the structural characteristics, however, we found no significant difference (except for regeneration density) between Lama Forest reserve and some of the other forests, while there were significant differences between forests in the same climatic zone and under similar soil conditions. There was also no significant relationship between
M.
kummel regeneration densities and soil characteristics. Indeed, regeneration individuals were lacking in many forests. Therefore, the difference, here, between
M.
andongensis and
M.
kummel may be due to other factors. Such differences between the two species’ characteristics might be mainly imputed to the difference in the levels of human pressures that the species, as well as their habitats faced [
37]. For instance,
M.
andongensis was found in Lama Forest reserve, one of the most important protected areas and probably the most preserved forest in Benin, while
M.
kummel occurs in riparian forests, in and out of protected areas that are exploited by local people. Furthermore, mean diameter was higher for
M.
kummel than
M.
andongensis, a result of an abundance of small-diameter trees in Lama Forest reserve relative to the riparian forests, where people (and animals) using the forest may impede natural regeneration or recruitment to larger size classes.
The relationships between soil characteristics and regeneration of
M.
andongensis were mostly negative or non-significant, highlighting the probable existence of other factors that influence its regeneration density [
38]. Competition with invasive species and seed predation are some factors that affect the regeneration of the species in Lama Forest reserve [
4]. Similarly, there was no significant influence of soil conditions on seedling and sapling abundance for
M.
kummel (in fact, regeneration lacks in several forests) and this could be due to predation of fruits and seeds [
39]. Fire is also often an important factor influencing species regeneration [
40] and this could be the case for both species in Benin. In fact, it has been shown that riparian forests in Benin face several human pressures such as fire, farm encroachment and timber extraction [
41]. Lama Forest reserve sometimes experiences accidental fires [
42], which might impact
M.
andongensis seeds. Although data indicate that the soil in Lama Forest reserve is more fertile and suitable for growth, the mean values for leaf morphological characteristics were higher for
M.
kummel than
M.
andongensis. The mean value in only one forest where
M.
kummel occurs was significantly lower than value in
M.
andongensis. Moreover, correlations between the morphological characteristics of both species and soil properties were weak. This suggests functional characteristics to be more responsible for variation in leaf morphology between the species.
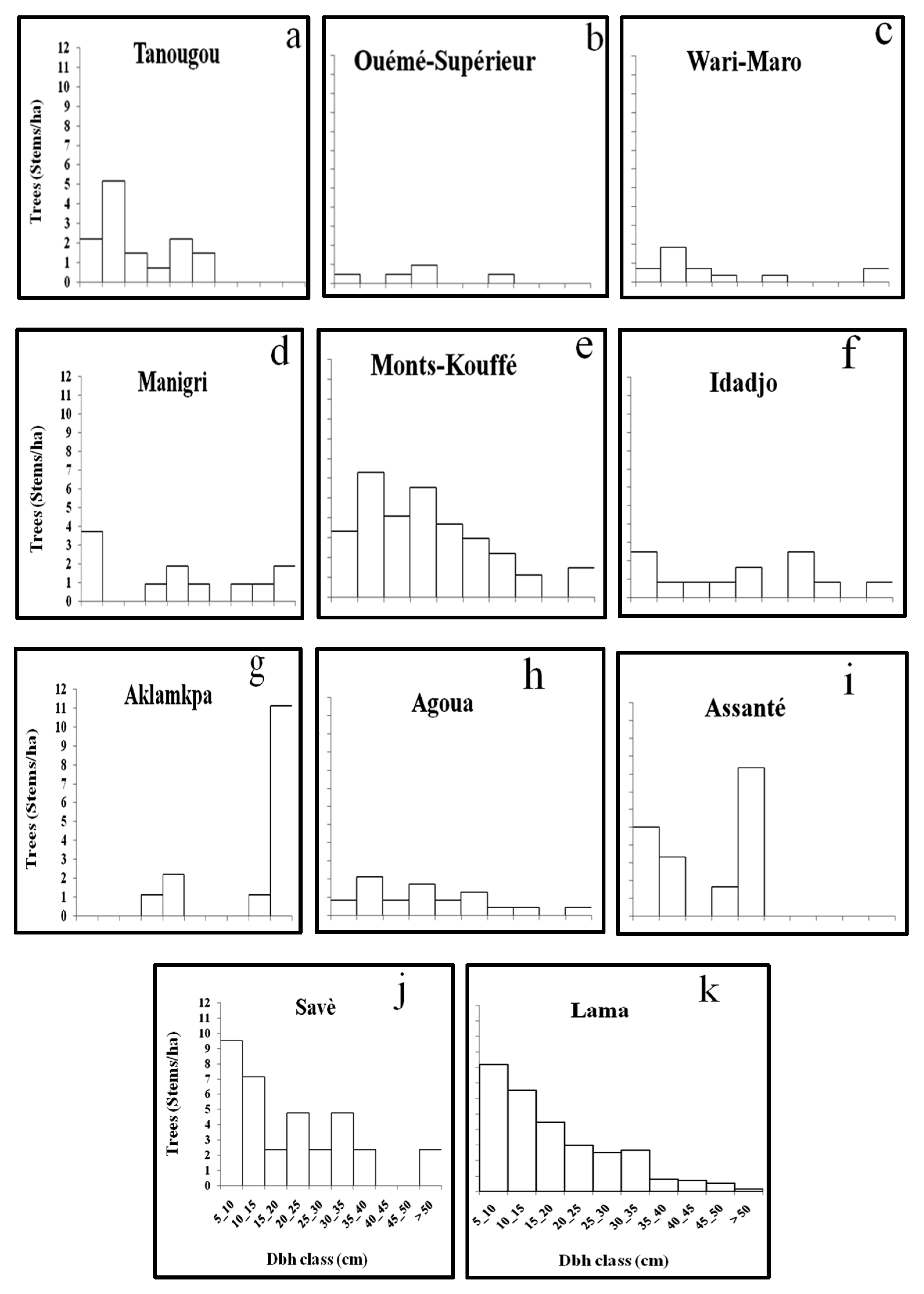
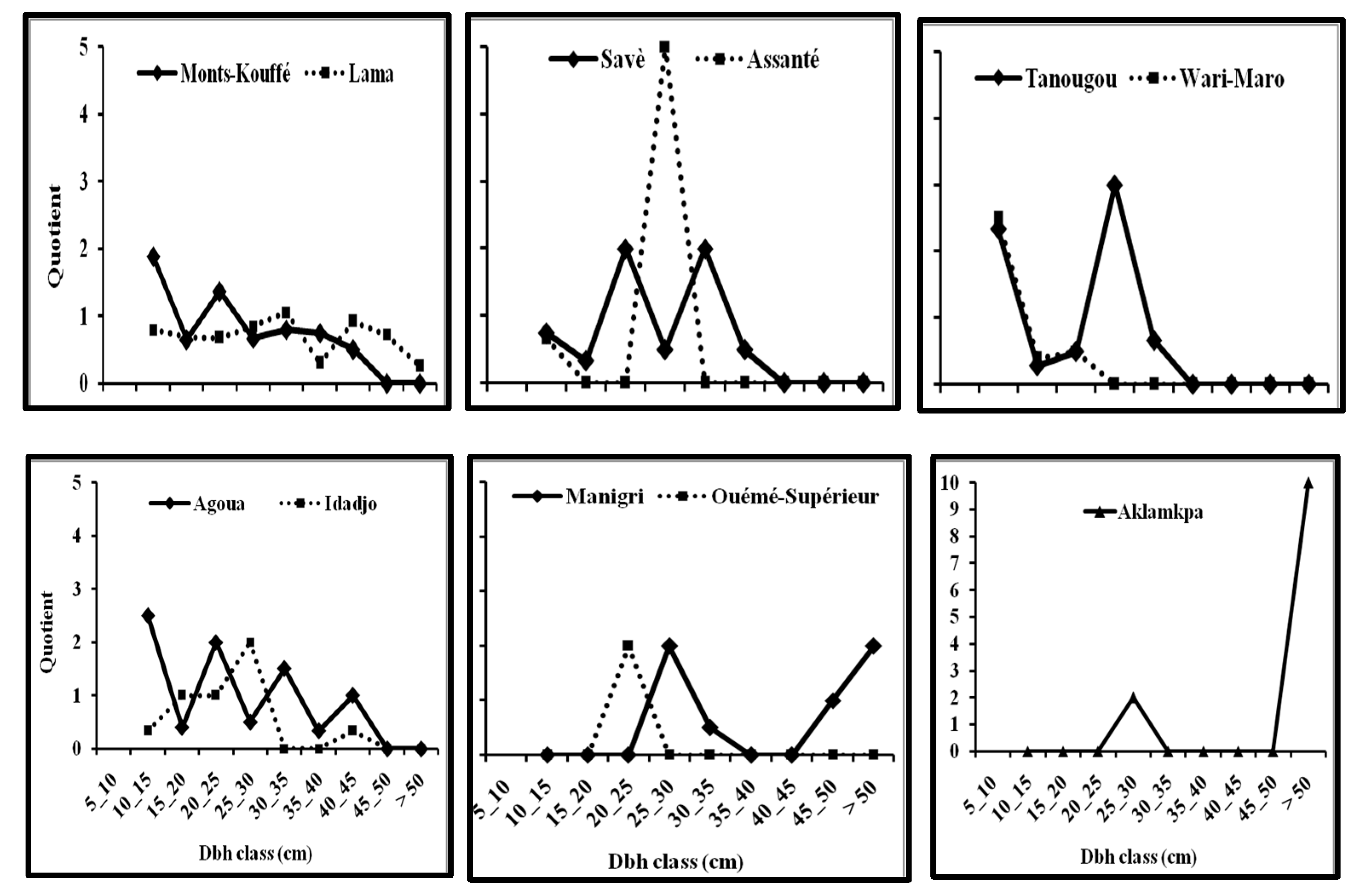
All diameter classes were represented in Lama Forest reserve, while in other forests there were missing classes. Added to this, the permutation and Simpson indices, as well as the quotients determined between successive diameter classes, indicated the population of
M.
andongensis to be more stable than the populations of
M.
kummel [
27,
43]. This may be due to differences in the levels of anthropogenic disturbance between Lama Forest reserve and the other forests. There were missing diameter classes in the populations of
M.
kummel and this might be a result of the impact of anthropogenic pressures followed by failures in regeneration and recruitment. However, populations of both species showed good proportions of trees with dbh 5–15 cm (>30 %) except two (Ouémé Supérieur and Aklamkpa), suggesting good recruitment. However, tree size is rarely a direct function of age, especially for long-lived species, and trees with dbh 5–15 cm might be older than they seemed [
44,
45]. This is confirmed in the case of
Mimusops species, for which trees with dbh 10 cm and 6 cm for
M.
andongensis and
M.
kummel respectively, exhibited flowers [
46], indicating the reproductive maturity of relatively small trees. This means that populations of
M.
kummel, within most forests, have sufficient reproductive trees but lack regeneration and recruits. For many long-lived species, populations can be maintained with a low or episodic recruitment [
47,
48]. However, this should be considered with caution especially in the context of climate change, which may threaten the species [
48]. We found no or very low regeneration in many forests and climate change could alter the hydrologic regime, potentially causing the death of adult trees, particularly in less humid parts of the range [
49]. Therefore, it is important to assess the mortality process of the species and factors likely to influence it.
Our results show that the population in the most protected forest in the country is more stable than populations in the other forests, with no missing diameter classes. This suggests the full protection of Lama Forest reserve to be beneficial for the preservation of
M.
andongensis populations, although the long-term benefits of protection might be mitigated by competition with invasive species, especially on regenerative stems [
15,
50]. The advantages of protecting forests for species conservation are well known [
51] and this management strategy has been applied for different vegetation types, with other forms of harvesting regulations [
52]. However, the full restriction of access to forests by local people (like in the case of Lama Forest reserve) might result in the loss of local knowledge/cultural heritage and therefore increase local people’s vulnerability during difficult times [
53]. Therefore, exploitation methods likely to limit negative impacts on the remaining populations of both species should be considered for sustainable management and conservation of both species and the biodiversity in general.
A limitation of this study is the short period of data collection. Indeed, only long-term investigation of population structure, in relation to the different ecological conditions and anthropogenic disturbances, will allow for observation of important changes for an effective understanding of the dynamics there between. This is also useful for prioritizing informed efficient management and conservation strategies [
54]. Thus, the species’ populations should be monitored for long-term data collection. For that, repeat monitoring in the installed plots is recommended at intervals of one or two years so that the necessary changes may be detected [
55]. Furthermore, the stands with regeneration failure should be studied further and prioritized for restoration purposes.
5. Conclusions
This study showed a significant difference between the population structures and morphologies of M. andongensis and M. kummel in Benin. Significant difference was also found between bioclimatic zones, while there was no significant difference between forests in different bioclimatic zones. Although the difference between the two species may be attributed to their specific functional characteristics, the results from this study also highlighted the influence of climate. Nonetheless, the two species occur under different soil conditions and this might contribute to the differences observed between them. However, we found no significant difference between forests with different soil characteristics, while there was significant difference between some forests in the same climatic zone and under similar soil conditions. This suggests plant-scale analysis to be the most sensitive, mainly because of the interference of anthropogenic pressures, which influence the observed effects of climate and soil. Thus, further investigation on the growth and demographic dynamics of both species, in relation to the different threats, could be useful to inform sustainable management options.
The relationships revealed between both species’ regeneration densities and soil characteristics were mostly negative or non-significant. Regeneration and recruits were also lacking in many forests, for M. kummel. This suggests the existence of other factors (e.g., fruit/seed predation, fire) that may affect regeneration in both species. Success in the regeneration and recruitment of plant species ensures population survival. Thus, for conservation purposes, further research would be welcomed to elucidate seed germination and development dynamics of regeneration as well as the influencing factors. Several soil properties (e.g., individual nutrient concentrations) were not considered in the analyses and it would be useful to have such information to assess how soil properties influence abundance of seedlings and saplings.
According to our results, the protection of forests is the key to sustaining species populations. However, the full protection of forests is at odds with the needs of local people for NTFPs, especially in the current context of climate change, which is likely to worsen rural peoples’ vulnerability in times of hardship. Also, the full protection of forests might lead to the loss of local knowledge on different useful species that are important for domestication purposes, and the cultural heritages and identities of many people. Therefore, exploitation strategies that are likely to limit the negative impacts on existing populations, of not only the target species but other species in demand, should be encouraged. Also, assessing the assisted germination or vegetative propagation of useful species should be investigated for species’ reintroduction.









{kind=link}
{kind=link}
{kind=link}
{kind=link}