
Ecological Gradient Analysis and Environmental Interpretation of Cyclocarya paliurus Communities


Abstract
:1. Introduction
2. Materials and Methods
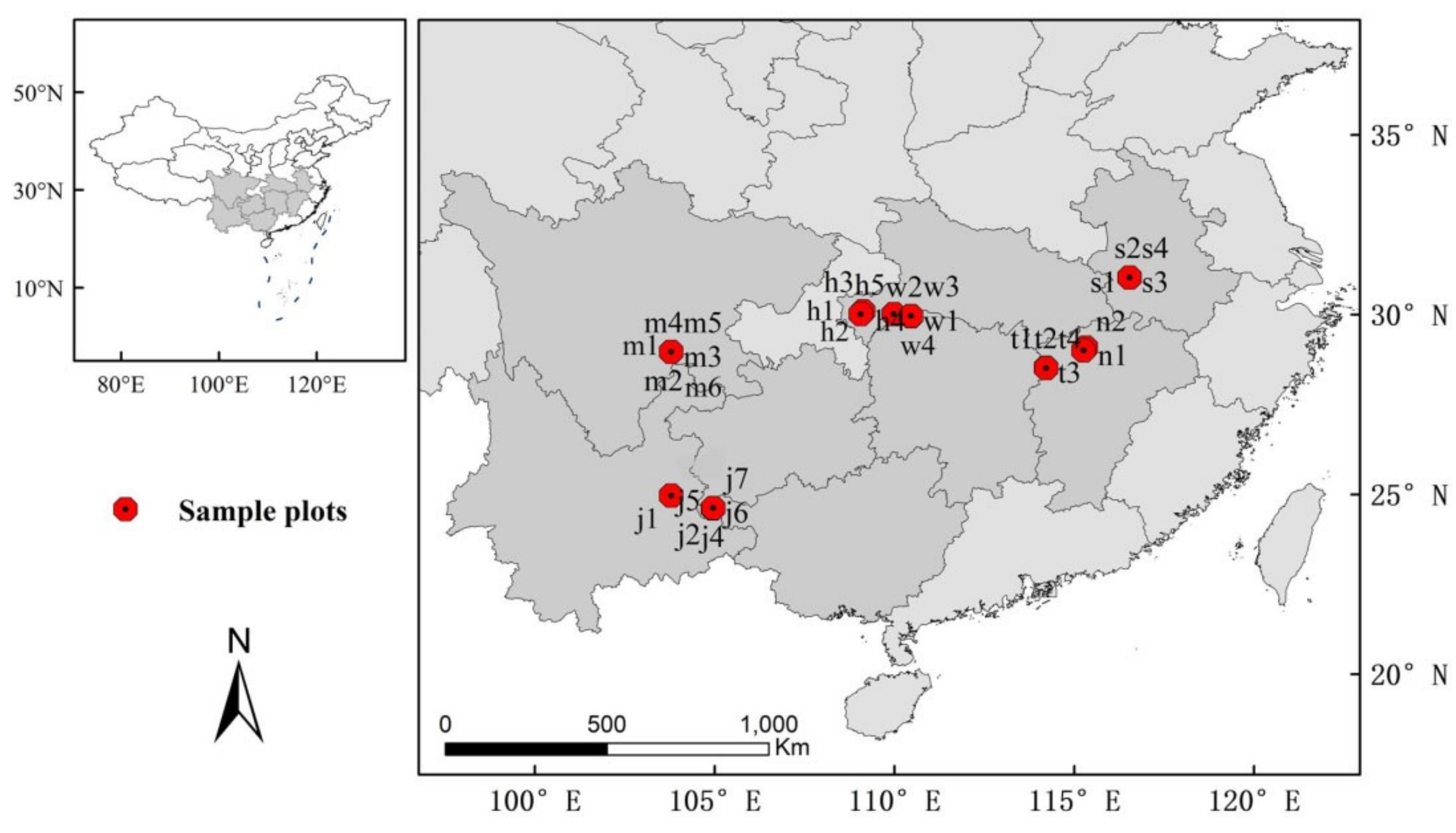
2.1. Data Collection
2.2. Data Analysis
- Species richness:
- Simpson diversity:
- Shannon–Wiener diversity:
- Pielou uniformity:
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Manchester, S.R.; Chen, Z.D.; Lu, A.M.; Uemura, K. Eastern Asian endemic seed plant genera and their paleogeographic history throughout the Northern Hemisphere. J. Syst. Evol. 2009, 47, 1–42. [Google Scholar] [CrossRef]
- Li, X.C.; Fu, X.X.; Shang, X.L.; Yang, W.X.; Fang, S.Z. Natural population structure and genetic differentiation for heterodicogamous plant: Cyclocarya paliurus, (Batal.) Iljinskaja (Juglandaceae). Tree. Genet. Genomes. 2017, 13, 80–89. [Google Scholar] [CrossRef]
- Fang, S.Z.; Wang, J.Y.; Wei, Z.Y.; Zhu, Z.X. Methods to break seed dormancy in Cyclocarya paliurus (Batal) Iljinskaja. Sci. Hortic-Amsterdam. 2006, 110, 305–309. [Google Scholar] [CrossRef]
- Kakar, M.U.; Naveed, M.; Saeed, M.; Zhao, S.C.; Rasheed, M.; Firdoos, S.; Manzoor, R.; Deng, Y.L.; Dai, R.J. A review on structure, extraction, and biological activities of polysaccharides isolated from Cyclocarya paliurus (Batalin) Iljinskaja. Int. J. Biol. Macromol. 2020, 156, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.H.; Dong, C.J.; Nie, S.P.; Li, F.; Wang, Z.J.; Shen, M.Y.; Xie, M.Y. Extraction, chemical composition and antioxidant activity of favonoids from Cyclocarya paliurus (Batal.) Iljinskaja leaves. Food Chem. 2015, 186, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.; Byrd, J.; Gao, Y.; Stubblefeld, J.; Park, H.; Dunlap, N. Isolation and structural clarifcation of triterpenes from Cyclocarya paliurus: Cyclocaric acid A and B. Planta. Med. 2014, 80, 139–143. [Google Scholar] [CrossRef]
- Liu, Y.; Cao, Y.N.; Fang, S.Z.; Wang, T.L.; Yin, Z.Q.; Shang, X.L.; Yang, W.X.; Fu, X.X. Antidiabetic effect of Cyclocarya paliurus leaves depends on the contents of antihyperglycemic flavonoids and antihyperlipidemic triterpenoids. Molecules 2018, 23, 1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, B.; Cao, Y.N.; Fang, S.Z.; Shang, X.L.; Yang, W.X.; Qian, C.Y. Variation and stability of growth and leaf flavonoid content in Cyclocarya paliurus across environments. Ind. Crop Prod. 2015, 76, 386–393. [Google Scholar] [CrossRef]
- Deng, B.; Fang, S.; Shang, X.; Fu, X.; Yang, W. Influence of genotypes and environmental factors on leaf triterpenoid content and growth of Cyclocarya paliurus. J. For. Res. 2018, 4, 1–10. [Google Scholar] [CrossRef]
- Fang, S.Z.; Yang, W.X.; Chu, X.L.; Shang, X.L.; She, C.Q.; Fu, X.X. Provenance and temporal variations in selected flavonoids in leaves of Cyclocarya paliurus. Food Chem. 2011, 124, 1382–1386. [Google Scholar] [CrossRef]
- Cao, Y.N.; Fang, S.Z.; Yin, Z.Q.; Fu, X.X.; Shang, X.L.; Yang, W.X.; Yang, H.M. Chemical fingerprint and multicomponent quantitative analysis for the quality evaluation of Cyclocarya paliurus leaves by HPLC-Q-TQF-MS. Molecules 2017, 22, 1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, B.; Fang, S.; Shang, X.; Fu, X.; Li, Y. Influence of provenance and shade on biomass production and triterpenoid accumulation in Cyclocarya paliurus. Agroforest Syst. 2017, 1, 1–10. [Google Scholar] [CrossRef]
- Liu, Y.; Qian, C.Y.; Ding, S.H.; Shang, X.L.; Yang, W.X.; Fang, S.Z. Effect of light regime and provenance on leaf characteristics, growth and flavonoid accumulation in Cyclocarya paliurus (Batal) Iljinskaja coppices. Bot. Stud. 2016, 57, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, D.; Xu, X.; Zhou, Z.X.; He, Y.H.; Wang, C.Z.; Shi, F.K.; Gao, L.L. Niche dynamics of main populations of old-tree communities in Jiuhua Mountain Scenic Area of Anhui Province. Chinese J. Ecol. 2019, 38, 1292–1304. [Google Scholar] [CrossRef]
- Chen, Y.F.; Yu, Z.P.; Chen, L.; Song, S.L.; Wen, R.Q.; Song, Q.N.; Yang, Q.P. Population dynamics and niche interpretation of Torreya grandis community in Guanshan National Nature Reserve of Jiangxi Province. South China For. Sci. 2019, 47, 7–11. [Google Scholar] [CrossRef]
- Xiong, X.G.; Xiong, G.; Xie, Z.Q. The Regeneration of Tree Species in the Mixed Evergreen-deciduous Broad-leaved Forests in the Shennongjia Mountains, Hubei Province. Acta Ecol. Sinica 2002, 22, 2001–2005. [Google Scholar]
- Shao, F.L.; Yu, X.X.; Zheng, J.K.; Wang, H.N. Relationships between dominant arbor species distribution and environmental factors of shelter forests in the Beijing mountain area. Acta Ecol. Sinica 2012, 32, 6092–6099. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.W.; Shang, X.L.; Ding, H.F.; Cao, Y.N.; Fang, S.Z. Natural variations in flavonoids and triterpenoids of Cyclocarya paliurus leaves. J. For. Res. 2020. [Google Scholar] [CrossRef]
- Liu, J.; Yin, M.L.; Zhang, W.L.; Tsang, D.C.W.; Wei, X.D.; Zhou, Y.T.; Xiao, T.F.; Wang, J.; Dong, X.J.; Sun, Y.B.; et al. Response of microbial communities and interactions to thallium in contaminated sediments near a pyrite mining area. Environ. Pollut. 2019, 248, 916–928. [Google Scholar] [CrossRef]
- Khatiwada, J.R.; Zhao, T.; Chen, Y.H.; Wang, B.; Xie, F.; Cannatella, D.C.; Jiang, J.P. Amphibian community structure along elevation gradients in eastern Nepal Himalaya. BMC Ecol. 2019, 19. [Google Scholar] [CrossRef] [Green Version]
- Fattahi, B.; Arzani, K.; Souri, M.K.; Barzegar, M. Effects of cadmium and lead on seed germination, morphological traits, and essential oil composition of sweet basil (Ocimum basilicum L.). Ind. Cr. Prod. 2019, 138. [Google Scholar] [CrossRef]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B. Vegan: Community Ecology Package; R Foundation for Statistical Computing: Vienna, Austria, 2007. [Google Scholar]
- Sun, C.; Jia, L.; Xi, B.; Wang, L.; Weng, X. Natural variation in fatty acid composition of Sapindus spp. Seed Oils. Ind. Cr. Prod. 2017, 102, 97–104. [Google Scholar] [CrossRef]
- He, Y.L.; He, Y.; Gong, L.L.; Fang, M.F.; Li, Z.H. Population genetic structure and interspecific differentiation between Acer davidii Franchi. and A-morrisonense Hayata (Aceraceae) based on SSR markers. Biochem. Syst. Ecol. 2017, 71, 42–49. [Google Scholar] [CrossRef]
- Jin, Z.X. A study of deciduous broad-l eaved forest feature in the Tiantai Mountain of Zhejiang. Guihaia 2000, 20, 107–116. [Google Scholar] [CrossRef]
- Kneitel, J.M.; Chase, J.M. Trade-offs in community ecology: Linking spatial scales and species coexistence. Ecol Lett. 2004, 7, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.H.; Jin, Y.X.; Zhao, Z.E.; Wu, J.Q.; Huang, H.D. The structure and dynamics of the rareplant communities in subtropical mountian of China. Acta Ecolog. Sinica 2000, 20, 800–807. [Google Scholar] [CrossRef]
- Wong, D.M.; Guo, R.; Zhang, H.W.; Cheng, Z.F.; Tian, Y.; Yu, J.J.; Wang, Y.X.; Liu, J.L. Species composition and 425 community structure of deciduous broad-leaved forests in National Nature Reserves of Zhejiang Province. J. Zhejiang Univ. 2014, 41, 600–610. [Google Scholar]
- Wang, Z.G.; Zhang, Z.X.; Wang, W.G.; Chu, J. Preliminary analysis of forest community structure of Yaoluoping National Nature Reserve in Yuexi County, Anhui Province, China. Chinese J. Plant Ecol. 2016, 40, 615–619. [Google Scholar] [CrossRef] [Green Version]
- Ren, L.; Luo, Y.H.; Wang, L.; Meng, Y.Y.; Liang, J.J.; Luo, L.J. Community Structure of Forest at Different Elevations in Cenwanglaoshan Natural Reserve. Guangxi For. Sci. 2018, 47, 139–144. [Google Scholar] [CrossRef]
- Yi, X.G.; Ding, H.; Fang, Y.M.; Ye, Y.Q.; Chen, S.F.; Wang, X.; Cong, R.; Zhang, K.W.; Li, Y.; Wang, X.R. Species diversity of forest communities at different altitudes based on fixed plot in Huangshan Mountains. J. Nanjing For. Univ. 2018, 42, 149–155. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, S.F.; Chen, Y.T.; Duan, H.P. Response of Architecture and Functions of Roots in Three Kinds of Sweet Gums Under Drought Stress. Soils 2013, 45, 1119–1126. [Google Scholar] [CrossRef]
- Qin, J.; Tang, X.H.; Yang, X.M. Effects of soil physical and chemical properties on different forest types of Pinus massoniana. Ecol. Environ. Sci. 2013, 22, 598–604. [Google Scholar]
- Fan, Y.; Lou, Y.K.; Ku, W.P.; Dai, Q.L.; Wang, Z.Y.; Zhao, M.S.; Yu, S.Q. Age structure and spatial point pattern of Phoebe sheareri population in Mount Tianmu. J. Zhejiang A F Univ. 2020, 37, 1027–1035. [Google Scholar] [CrossRef]
- Wu, X.Y.; Zhou, Z.H.; Wu, Y.Q.; Yao, F.P.; Ni, R.X.; Wang, P.; Yao, J. Structure Characteristics and Species Association of Dendropanax dentiger in Communities. J Zhejiang For. Sci. Technol. 2020, 40, 24–29. [Google Scholar] [CrossRef]
- Jia, Y.; Yang, J.; He, Y.L.; He, Y.; Niu, C.; Gong, L.L.; Li, Z.H. Characterization of the whole chloroplast genome sequence of Acer davidii Franch (Aceraceae). Conserv. Genet. Resour. 2016, 8, 141–143. [Google Scholar] [CrossRef]
- Zhang, L.; Pan, J.R.; Zhu, C. Determination of the geographical origin of Chinese teas based on stable carbon and nitrogen isotope ratios. J Zhejiang Univ. 2012, 13, 824–830. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.Q.; Li, H.; Hoang, T.X.; Ruan, X.; Zhang, Y.; Arkorful, E.; Chen, X.; Sun, K.; Li, X.H. Genetic Diversity And Relationship Of Dongting Biluochun Tea Germplasm In Suzhou Revealed By Ssr Markers. Pak J Bot. 2019, 51, 895–902. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Li, Y.; Fang, S.Z. MaxEnt model-based identification of potential Cyclocarya paliurus cultivation regions. J. Nanjing For. Univ. 2017, 41, 25–29. [Google Scholar] [CrossRef]
- Jin, Z.X. Species Diversity of Yukeng Forest Communities in Xianju of Zhejiang. J. Wuhan Bot. Res. 2001, 19, 225–232. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Plots | Lat | Lon | Slo | Asp | Alt | Tem | LTem | HTem | Prec | pH | SW1 | SW2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (°N) | (°E) | (°) | (mm) | (°C) | (°C) | (°C) | (mm) | (g/cm3) | (g/cm3) | |||
| h1 h2 h3 h4 h5 | 109.13–109.98 | 30.1–30.02 | 12–48 | 1–5 | 1106–1283 | 12.99 | –5.28 | 28.5 | 2100 | 4.54–5.7 | 1.04–1.62 | 0.97–1.48 |
| j1 j2 j4 j5 j6 j7 | 104.95–104.96 | 24.61–24.63 | 10–40 | 2–5 | 1539–1829 | 18.91 | 3.54 | 33 | 1326 | 3.83–4.49 | 0.55–0.85 | 0.54–0.9 |
| m1 m2 m3 m4 m5 m6 | 103.79 | 28.97 | 19–32 | 1–8 | 1192–1230 | 15.85 | 1.38 | 29.71 | 1589 | 3.18–3.39 | 0.57–0.73 | 0.68–0.89 |
| n1 n2 | 115.26–115.32 | 29.01–29.09 | 22 | 8 | 771–780 | 16.75 | –1.83 | 33.1 | 1862 | 4.59–4.64 | 0.92–0.97 | 0.96–1 |
| s1 s2 s3 s4 | 116.54 | 31.04 | 24–45 | 3–7 | 764–831 | 14.45 | –8.12 | 31.32 | 1603 | 4.88–5.28 | 0.99–1.11 | 1.07–1.18 |
| t1 t2 t3 t4 | 114.21–114.22 | 28.52–28.53 | 38–50 | 5–7 | 790–860 | 16.82 | –1.85 | 32.7 | 1487 | 4.55–4.69 | 0.76–0.83 | 0.85–0.97 |
| w1 w2 w3 w4 | 110.43–110.46 | 29.86–29.97 | 18–38 | 2–6 | 1340–1536 | 13.12 | –5.49 | 28.65 | 1704 | 5.29–5.92 | 0.85–1.04 | 1.06–1.19 |
| Tree Species | IV | Sample Plots | No of Sample Plots | Crossing Regions |
|---|---|---|---|---|
| Cyclocarya paliurus (Batal.) Iljinsk. | 7.42 | all | 31 | 7 |
| Cunninghamia lanceolata (Lamb.) Hook. | 0.45 | h3, h5, t1, t2, t3 | 5 | 2 |
| Liquidambar formosana Hance | 0.42 | j1, j5, j6, s3, n1, h3, h4 | 7 | 4 |
| Ilex ficoidea Hemsl. | 0.42 | m3, m4, m5, m6, t3 | 5 | 2 |
| Cinnamomum wilsonii Gamble | 0.4 | h3, h5, j5, m1, m2, m3, m4, m5, m6 | 9 | 3 |
| Litsea elongata (Wall. ex Nees) Benth. et Hook. f. | 0.38 | h3, h4, n1, n2, t1, t4 | 6 | 3 |
| Taxus chinensis (Pilger) Rehd. | 0.36 | t1, t2, t3, t4 | 4 | 1 |
| Ilex centrochinensis S. Y. Hu | 0.33 | m1, m2, m3, m4, m5, m6 | 6 | 1 |
| Cryptomeria fortunei Hooibrenk ex Otto et Dietr. | 0.32 | h1, h5 | 2 | 1 |
| Acer davidii Franch. | 0.32 | h1, h4, h5, j1, j5, j7, m1, m3, m5, n2, s2 | 11 | 5 |
| Dendrobenthamia capitata (Wall.) Hutch. | 0.31 | w1, w2, w3, w4 | 4 | 1 |
| Lithocarpus glaber (Thunb.) Nakai | 0.3 | j1, j2, j4, j5 | 4 | 1 |
| Castanopsis fargesii Franch. | 0.3 | m2, m3, m4, m6, j5, j7 | 6 | 2 |
| Bothrocaryum controversum (Hemsl.) Pojark | 0.28 | w1 w2 w3 h1, h2, n1 | 6 | 3 |
| Swida macrophylla (Wall.) Sojak | 0.28 | w2 w4 h2, h4, | 4 | 2 |
| Lithocarpus harlandii (Seem.) Rehd. et Wils. | 0.27 | h1, h2, h3, h4, h5, m3, m4, m5, m6 | 9 | 2 |
| Lindera communis Hemsl. | 0.27 | h1, j2, s3, s4 | 4 | 3 |
| Platycarya strobilacea Sieb. Et Zucc. | 0.26 | n2, t1, h3, h4, h5 | 5 | 3 |
| Clerodendrum japonicum (Thunb.) Sweet | 0.24 | j2, j4, h2 | 3 | 2 |
| Diplospora dubia (Lindl.) Masam. | 0.23 | j4, m1, m2, m4, m5, w3, | 6 | 3 |
| Castanea henryi (Skan) Rehd. et Wils. | 0.23 | h2, h5, | 2 | 1 |
| Acer palmatum Thunb. | 0.23 | w1 w2 w3 w4 t1, t2, | 6 | 2 |
| Acer elegantulum Fang et P. L. Chiu | 0.23 | h1, h3, j1, j2, j7, m4, m5, m6, w3 | 9 | 4 |
| Camellia cuspidata (Kochs) Wright ex Gard. | 0.23 | j1, j2, j4, m5 | 4 | 2 |
| Pinus massoniana Lamb. | 0.22 | h5, n2, s4 | 3 | 3 |
| Dendropanax dentiger (Harms) Merr. | 0.21 | h3, h4, m3, m5, m6, t1, t3, t4 | 8 | 3 |
| Tapiscia sinensis Oliv. | 0.2 | s1, s2, s3 | 3 | 1 |
| Eurya japonica Thunb. | 0.19 | h3, h5, m2, m4, m5, m6, n1, n2 | 8 | 3 |
| Carpinus viminea Lindley | 0.19 | n2, s1, w1, w3, w4 | 5 | 3 |
| Picrasma quassioides (D. Don) Benn. | 0.18 | s4, t2, t4, w1, w3 | 5 | 3 |
| Sum of the top 30 trees | 15.51 | |||
| Sum of all 190 tree species | 22.96 |
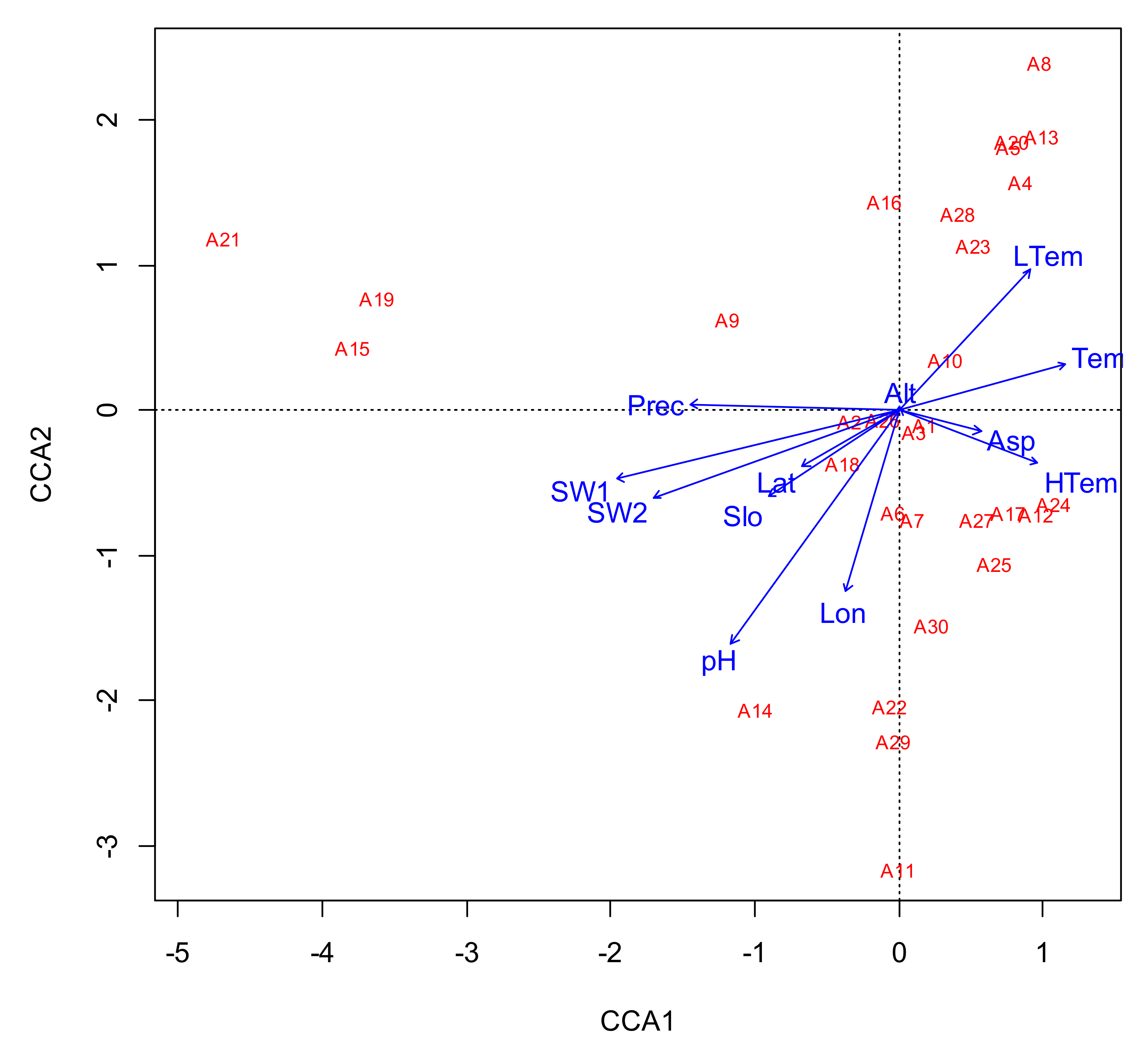
| CCA1 | CCA2 | CCA3 | CCA4 | Total | |
|---|---|---|---|---|---|
| Eigenvalue | 0.500 | 0.407 | 0.381 | 0.303 | |
| Proportion Explained | 0.117 | 0.095 | 0.089 | 0.071 | |
| Cumulative Proportion | 0.117 | 0.212 | 0.300 | 0.371 | 0.612 |
| Longitude | −0.158 | −0.601 | −0.090 | 0.499 | |
| Latitude | −0.293 | −0.184 | 0.353 | 0.403 | |
| Slope | −0.393 | −0.283 | −0.217 | 0.147 | |
| Aspect | 0.247 | −0.071 | −0.023 | 0.301 | |
| Altitude | 0.000 | 0.010 | −0.105 | −0.717 | |
| Annual mean temperature | 0.503 | 0.153 | −0.401 | −0.147 | |
| Annual lowest temperature | 0.397 | 0.464 | −0.137 | −0.318 | |
| Annual highest temperature | 0.419 | −0.175 | −0.496 | 0.119 | |
| Precipitaion | −0.626 | 0.021 | 0.087 | 0.328 | |
| pH | −0.506 | −0.771 | −0.060 | 0.197 | |
| 0–20 cm soil bulk density | −0.849 | −0.228 | −0.048 | 0.285 | |
| 20–40 cm soil bulk density | −0.740 | −0.287 | 0.127 | 0.400 | |
| Model Pr(>F) | 0.001 *** | ||||
| Sample | Ri | Shanon | Pielou | Simpson | RAb | Rc | Rf | IV |
|---|---|---|---|---|---|---|---|---|
| h1 h2 h3 h4 h5 | 11–32 | 2.04–2.90 | 0.77–0.87 | 0.82–0.90 | 0.09–0.28 | 0.32–0.44 | 0.06 | 0.17–0.24 |
| j1 j2 j4 j5 j6 j7 | 7–23 | 1.76–2.56 | 0.76–0.91 | 0.78–0.90 | 0.06–0.32 | 0.17–0.62 | 0.06 | 0.11–0.31 |
| m1 m2 m3 m4 m5 m6 | 10–23 | 1.82–2.81 | 0.74–0.90 | 0.77–0.93 | 0.06–0.12 | 0.09–0.74 | 0.06 | 0.08–0.31 |
| n1 n2 | 17–18 | 2.34–2.38 | 0.82–0.83 | 0.84–0.87 | 0.23–0.24 | 0.25–0.47 | 0.06 | 0.18–0.26 |
| s1 s2 s3 s4 | 6–14 | 1.36–2.09 | 0.73–0.79 | 0.66–0.79 | 0.29–0.4 | 0.54–0.69 | 0.06 | 0.30–0.36 |
| t1 t2 t3 t4 | 7–16 | 1.69–1.96 | 0.71–0.89 | 0.48–0.80 | 0.24–0.39 | 0.43–0.80 | 0.06 | 0.26–0.37 |
| w1 w2 w3 w4 | 21–36 | 2.392–3.148 | 0.79–0.88 | 0.84–0.93 | 0.07–0.23 | 0.19–0.49 | 0.06 | 0.11–0.26 |
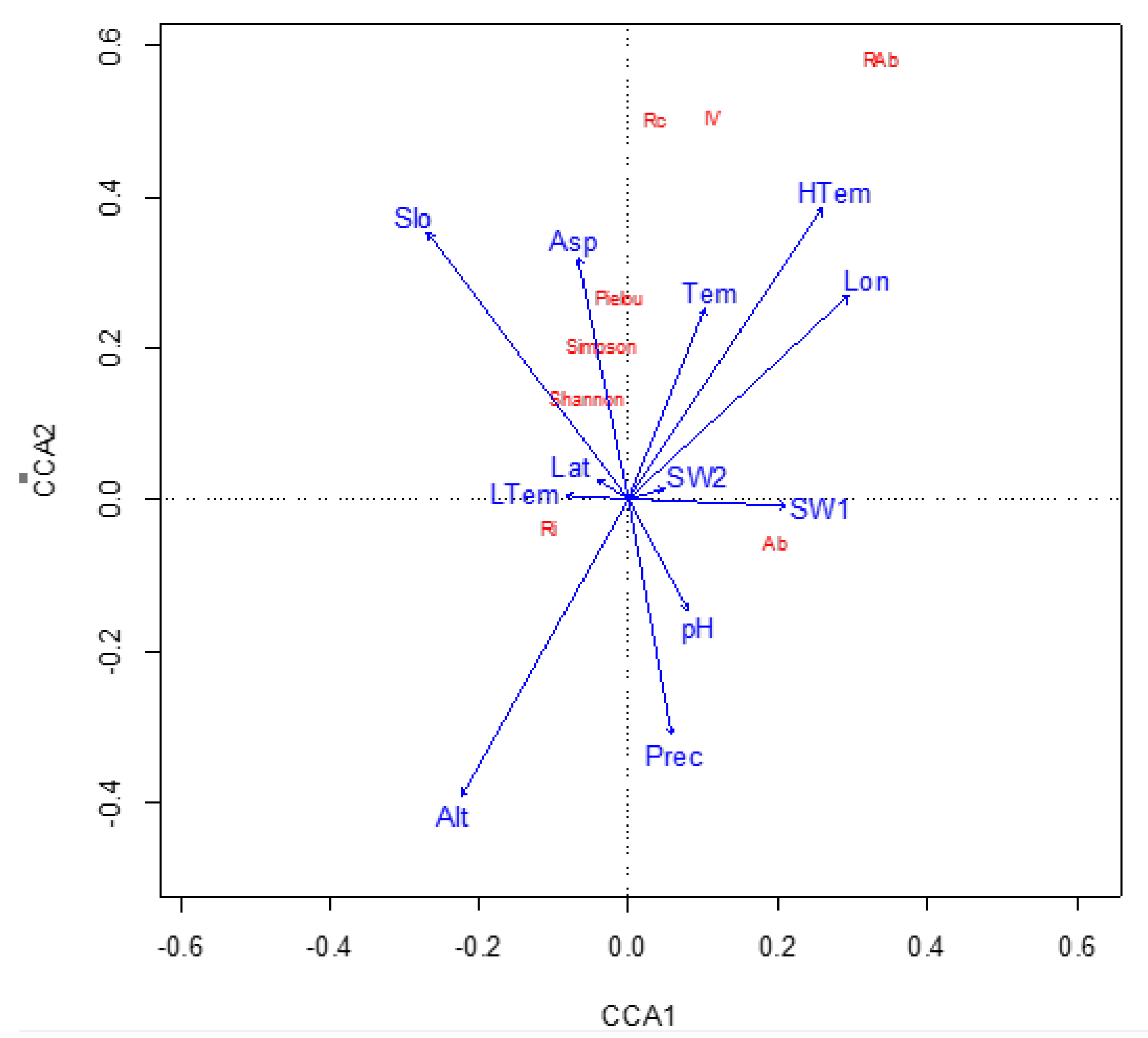
| CCA1 | CCA2 | CCA3 | CCA4 | Total | |
|---|---|---|---|---|---|
| Eigenvalue | 0.019 | 0.014 | 0.001 | 0.000 | |
| Proportion Explained | 0.356 | 0.269 | 0.020 | 0.007 | |
| Cumulative Proportion | 0.356 | 0.625 | 0.645 | 0.652 | 0.652 |
| C. paliurus relative Abundance | 0.340 | 0.585 | 0.019 | −0.204 | |
| C. paliurus relative coverage | 0.035 | 0.504 | 0.158 | 0.065 | |
| C. paliurus important value | 0.114 | 0.507 | 0.104 | −0.012 | |
| C. paliurus Abundance | 0.198 | −0.057 | 0.000 | 0.004 | |
| Tree species richness | −0.104 | −0.038 | 0.009 | −0.004 | |
| Shannon index | −0.054 | 0.137 | −0.057 | 0.008 | |
| Pielou index | −0.009 | 0.269 | −0.079 | 0.013 | |
| Simpson index | −0.036 | 0.202 | −0.071 | 0.022 | |
| Model | 0.001 *** | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Zhou, Y.; Fang, S.; Shang, X. Ecological Gradient Analysis and Environmental Interpretation of Cyclocarya paliurus Communities. Forests 2021, 12, 146. https://doi.org/10.3390/f12020146
Sun C, Zhou Y, Fang S, Shang X. Ecological Gradient Analysis and Environmental Interpretation of Cyclocarya paliurus Communities. Forests. 2021; 12(2):146. https://doi.org/10.3390/f12020146
Chicago/Turabian StyleSun, Caowen, Yongsheng Zhou, Shengzuo Fang, and Xulan Shang. 2021. "Ecological Gradient Analysis and Environmental Interpretation of Cyclocarya paliurus Communities" Forests 12, no. 2: 146. https://doi.org/10.3390/f12020146
APA StyleSun, C., Zhou, Y., Fang, S., & Shang, X. (2021). Ecological Gradient Analysis and Environmental Interpretation of Cyclocarya paliurus Communities. Forests, 12(2), 146. https://doi.org/10.3390/f12020146