
Growth and Physiological Responses of Norway Spruce (Picea abies (L.) H. Karst) Supplemented with Monochromatic Red, Blue and Far-Red Light

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
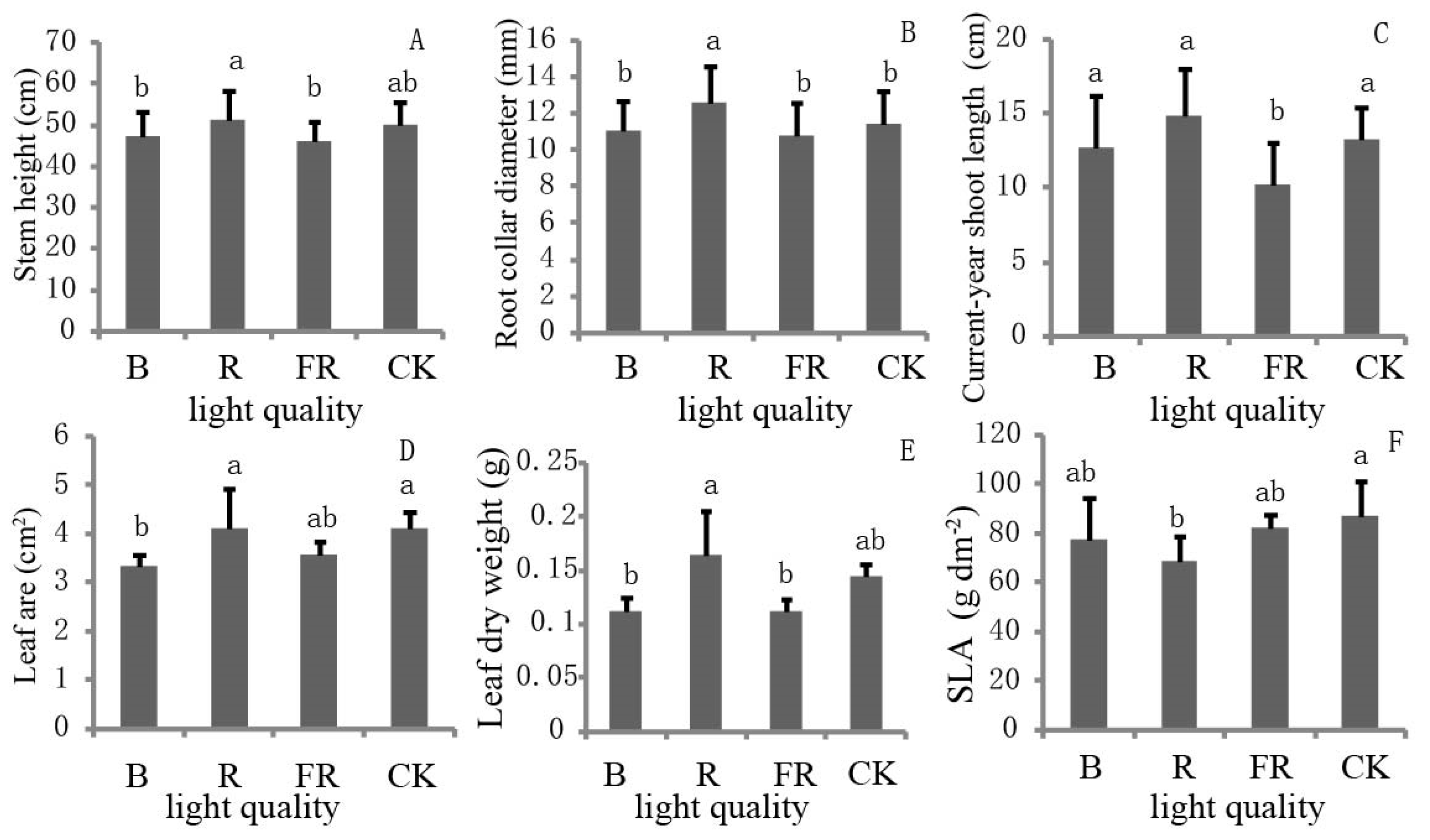
2.2. Growth and Needle Traits
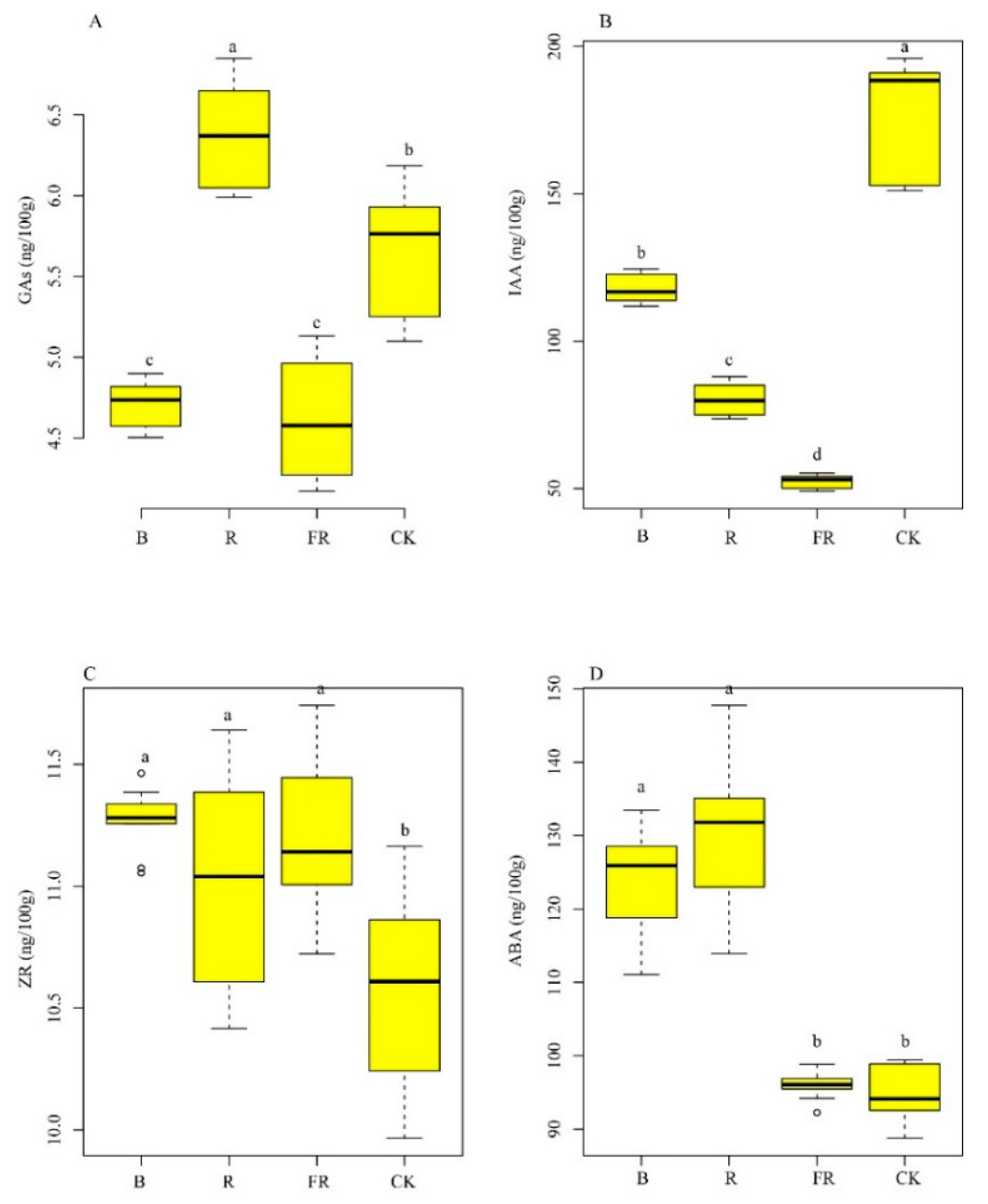
2.3. Phytohormone Level
2.4. Chlorophyll Content
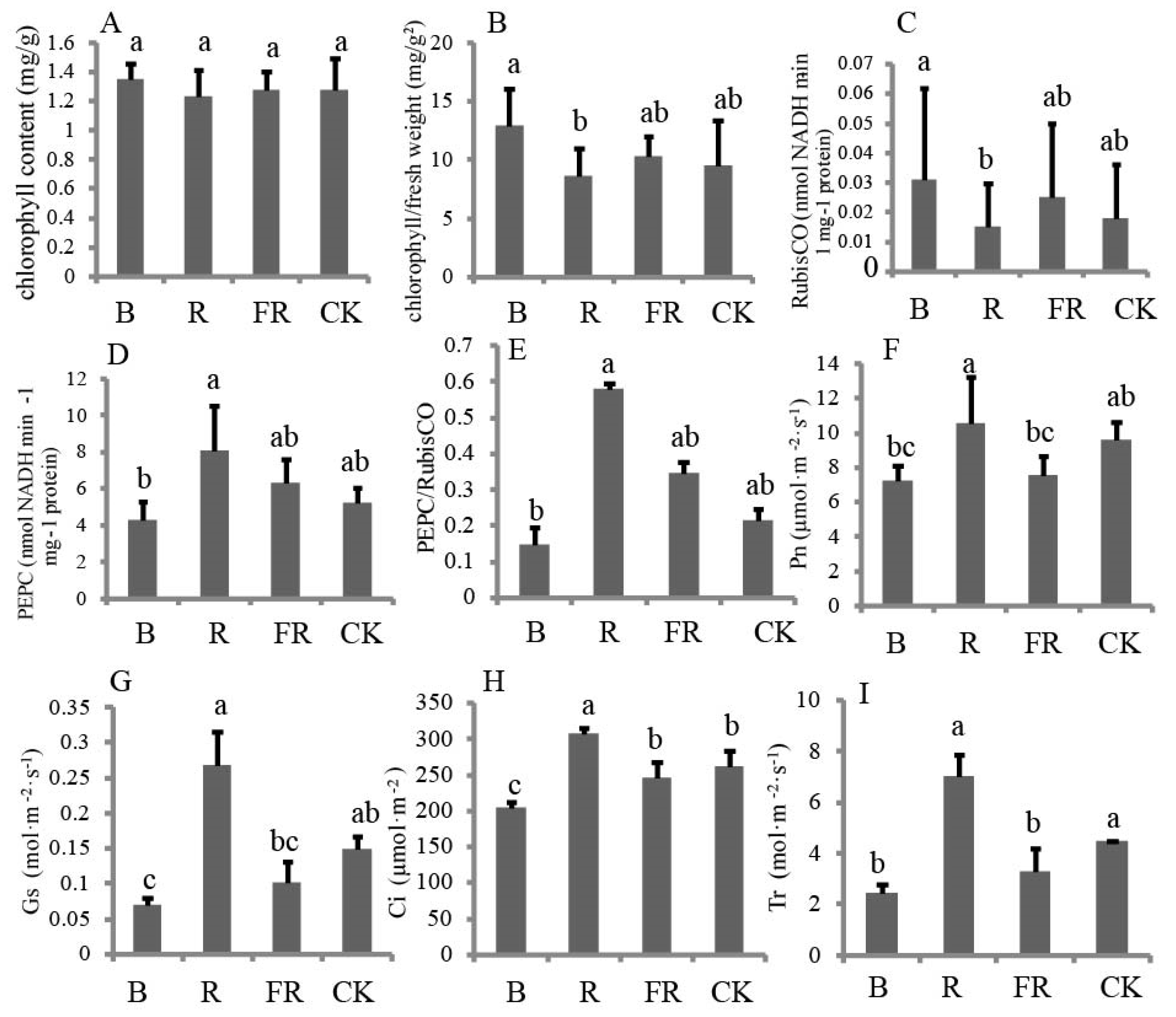
2.5. Photosynthetic Enzymes
2.6. Gas-Exchange Measurements
2.7. Chlorophyll a Fluorescence
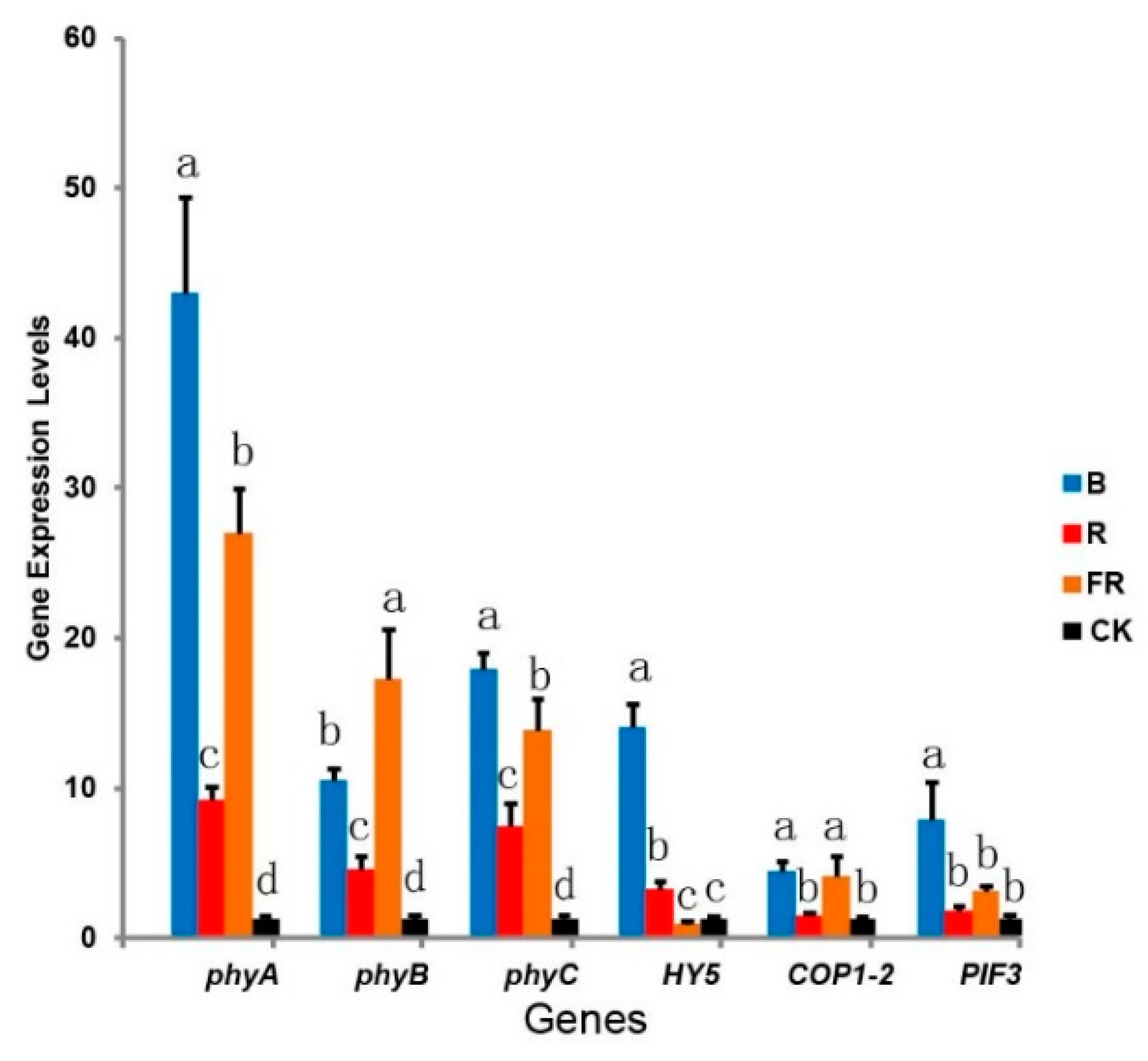
2.8. Photosynthesis-Related Gene Expression
2.9. Statistical Analysis
3. Results
3.1. Effect of Light Quality on Growth Performance
3.2. Effect of Light Quality on Phytohormone Level
3.3. Effect of Light Quality on Chlorophyll Content
3.4. Effect of Light Quality on Photosynthetic Enzyme and Gas Exchange Parameters
3.5. Effect of Light Quality on Chlorophyll Fluorescence Parameters
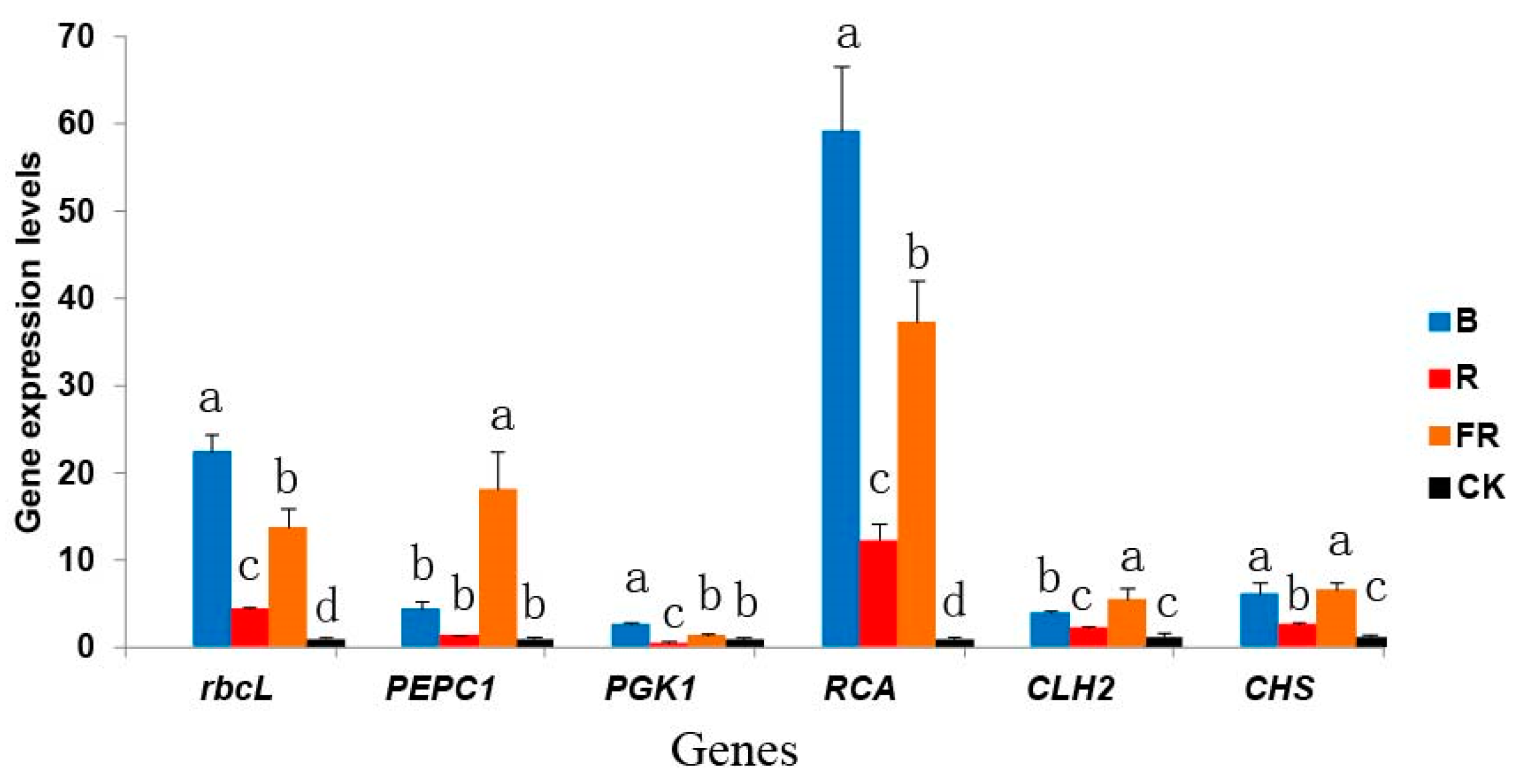
3.6. Effect of Light Quality on Photosynthesis-Related Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olsen, J.E.; Junttila, O. Far red end-of-day treatment restores wild type-like plant length in hybrid aspen overexpressing phytochrome A. Physiol. Plant 2002, 115, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Quail, P.H. Photosensory perception and signaling in plant cells: New paradigms? Curr. Opin. Cell Biol. 2002, 14, 180–188. [Google Scholar] [CrossRef]
- Kong, S.-G.; Okajima, K. Diverse photoreceptors and light responses in plants. J. Plant Res. 2016, 129, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, K.A.; Whitelam, G.C. Phytochromes and Shade-avoidance Responses in Plants. Ann. Bot. 2005, 96, 169–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, H. Phytochromes and light signal perception by plants—An emerging synthesis. Nat. Cell Biol. 2000, 407, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Runkle, E.S.; Heins, R.D. Photocontrol of flowering and extension growth in the long-day plant pansy. J. Amer. Soc. Hort. Sci. 2003, 128, 479–485. [Google Scholar] [CrossRef]
- Bae, J.-H.; Park, S.-Y.; Oh, M.-M. Supplemental irradiation with far-red light-emitting diodes improves growth and phenolic contents in Crepidiastrum denticulatum in a plant factory with artificial lighting. Hortic. Environ. Biotechnol. 2017, 58, 357–366. [Google Scholar] [CrossRef]
- Bae, J.-H.; Park, S.-Y.; Oh, M.-M. Growth and phenolic compounds of Crepidiastrum denticulatum under various blue light intensities with a fixed phytochrome photostationary state using far-red light. Hortic. Environ. Biotechnol. 2019, 60, 199–206. [Google Scholar] [CrossRef]
- Fan, X.; Zang, J.; Xu, Z.; Guo, S.; Jiao, X.; Liu, X.; Gao, Y. Effects of different light quality on growth, chlorophyll concentration and chlorophyll biosynthesis precursors of non-heading Chinese cabbage (Brassica campestris L.). Acta Physiol. Plant 2013, 35, 2721–2726. [Google Scholar] [CrossRef]
- Håbjørg, A. Effects of light quality, light intensity and night temperature on growth and development of three latitudinal populations of Betula pubescens Ehrh. Nor. Landbr. Meld 1972, 51, 1–7. [Google Scholar]
- Clapham, D.H.; Dormling, I.; Ekberg, L.; Eriksson, G.; Qamaruddin, M.; Vince-Prue, D. Latitudinal cline of requirement for far-red light for the photoperiodic control of budset and extension growth in Picea abies (Norway spruce). Physiol. Plant 1998, 102, 71–78. [Google Scholar] [CrossRef]
- Clapham, D.H.; Ekberg, I.; Eriksson, G.; Norell, L.; Vince-Prue, D. Requirement for far-red light to maintain secondary needle extension growth in northern but not southern populations of Pinus sylvestris (Scots pine). Physiol. Plant 2002, 114, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Molmann, J.A.; Junttila, O.; Johnsen, O.; Olsen, J.E. Effects of red, far-red and blue light in maintaining growth in latitudinal populations of Norway spruce (Picea abies). Plant Cell Environ. 2005, 29, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Riikonen, J.; Kettunen, N.; Gritsevich, M.; Hakala, T.; Särkkä, L.; Tahvonen, R. Growth and development of Norway spruce and Scots pine seedlings under different light spectra. Environ. Exp. Bot. 2016, 121, 112–120. [Google Scholar] [CrossRef]
- Fløistad, I.S.; Patil, G.G. Growth and Terminal Bud Formation in Picea abies Seedlings Grown with Alternating Diurnal Temperature and Different Light Qualities. Scand. J. For. Res. 2002, 17, 15–27. [Google Scholar] [CrossRef]
- Sellin, A.; Sack, L.; Õunapuu-Pikas, E.; Karusion, A. Impact of light quality on leaf and shoot hydraulic properties: A case study in silver birch (Betula pendula). Plant. Cell Environ. 2011, 34, 1079–1087. [Google Scholar] [CrossRef]
- Fernbach, E.; Mohr, H. Coaction of blue/ultraviolet-A light and light absorbed by phytochrome in controlling growth of pine (Pinus sylestris L.) seedlings. Planta 1990, 180, 212–216. [Google Scholar] [CrossRef]
- Szymańska, R.; Nowicka, B.; Gabruk, M.; Glińska, S.; Michlewska, S.; Dlużwska, J.; Sawicka, A.; Kruk, J.; Laitinen, R. Physiological and antioxidant responses of two accessions of Arabidopsis thaliana in different light and temperature conditions. Physiol. Plant 2015, 154, 194–209. [Google Scholar] [CrossRef]
- Kim, J.H.; Glick, R.E.; Melis, A. Dynamics of Photosystem Stoichiometry Adjustment by Light Quality in Chloroplasts. Plant Physiol. 1993, 102, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Krekling, T.; Appelgren, M. Light quality affects photosynthesis and leaf anatomy of birch plantlets in vitro. Plant Cell Tissue Organ Cult. (PCTOC) 1995, 41, 177–185. [Google Scholar] [CrossRef]
- Grams, T.E.E.; Thiel, S. High light-induced switch from C3-photosynthesis to Crassulacean acid metabolism is mediated by UV-A/blue light. J. Exp. Bot. 2002, 53, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Shimazaki, K.-I.; Doi, M.; Assmann, S.M.; Kinoshita, T. Light Regulation of Stomatal Movement. Annu. Rev. Plant Biol. 2007, 58, 219–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasperbauer, M.J.; Hamilton, J.L. Chloroplast Structure and Starch Grain Accumulation in Leaves That Received Different Red and Far-Red Levels during Development. Plant Physiol. 1984, 74, 967–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsinoremas, N.; Schaefer, M.; Golden, S. Blue and red light reversibly control psbA expression in the cyanobacterium Synechococcus sp. strain PCC 7942. J. Biol. Chem. 1994, 269, 16143–16147. [Google Scholar] [CrossRef]
- Allen, J.F. Thylakoid protein phosphorylation, state 1-state 2 transitions, and photosystem stoichiometry adjustment: Redox control at multiple levels of gene expression. Physiol. Plant 1995, 93, 196–205. [Google Scholar] [CrossRef]
- El Bissati, K.; Kirilovsky, D. Regulation of psbA and psaE Expression by Light Quality in Synechocystis Species PCC 6803. A Redox Control Mechanism. Plant Physiol. 2001, 125, 1988–2000. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, F.; Mao, J.-F.; Wang, J.; Zhang, S.; Li, Y. Transcriptome Analysis Reveals that Red and Blue Light Regulate Growth and Phytohormone Metabolism in Norway Spruce [Picea abies (L.) Karst.]. PLoS ONE 2015, 10, e0127896. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Yao, J.; Wang, X.; Guo, H.; Duan, D. Transcriptome Sequencing and Comparative Analysis of Saccharina japonica (Laminariales, Phaeophyceae) under Blue Light Induction. PLoS ONE 2012, 7, e39704. [Google Scholar] [CrossRef]
- Siipola, S.M.; Kotilainen, T.; Sipari, N.; Morales, L.O.; Lindfors, A.V.; Robson, T.M.; Aphalo, P.J. Epidermal UV-A absorbance and whole-leaf flavonoid composition in pea respond more to solar blue light than to solar UV radiation. Plant Cell Environ. 2015, 38, 941–952. [Google Scholar] [CrossRef]
- Lau, O.S.; Deng, X.W. Plant hormone signaling lightens up: Integrators of light and hormones. Curr. Opin. Plant Biol. 2010, 13, 571–577. [Google Scholar] [CrossRef]
- Jiao, Y.; Lau, O.S.; Deng, X.W. Light-regulated transcriptional networks in higher plants. Nat. Rev. Genet. 2007, 8, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Soy, J.; Leivar, P.; Monte, E. PIF1 promotes phytochrome-regulated growth under photoperiodic conditions in Arabidopsis together with PIF3, PIF4, and PIF5. J. Exp. Bot. 2014, 65, 2925–2936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Ji, R.; Li, H.; Zhao, T.; Liu, J.; Lin, C.; Liu, B. Constans-like 7 (COL7) is involved in phytochrome B (phyB)-mediated light-quality regulation of auxin homeostasis. Mol. Plant 2014, 7, 1429–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouril, R.; Wientjes, E.; Bultema, J.B.; Croce, R.; Boekema, E.J. High-light vs. low-light: Effect of light acclimation on photosystem II composition and organization in Arabidopsis thaliana. Biochim. et Biophys. Acta (BBA) Gen. Subj. 2013, 1827, 411–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beel, B.; Prager, K.; Spexard, M.; Sasso, S.; Weiss, D.; Müller, N.; Heinnickel, M.; Dewez, D.; Ikoma, D.; Grossman, A.R.; et al. A flavin binding Cryptochrome photoreceptor responds to both blue and red light in Chlamydomonas reinhardtii. Plant Cell 2012, 24, 2992–3008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.J.; Wang, F.-J.; Sun, X.-T.; Liu, F.-L.; Liang, Z.-R. Comparison of transcriptome under red and blue light culture of Saccharina japonica (Phaeophyceae). Planta 2013, 237, 1123–1133. [Google Scholar] [CrossRef]
- Lu, N.; Maruo, T.; Johkan, M.; Hohjo, M.; Tsukagoshi, S.; Ito, Y.; Ichimura, T.; Shinohara, Y. Effects of Supplemental Lighting with Light-Emitting Diodes (LEDs) on Tomato Yield and Quality of Single-Truss Tomato Plants Grown at High Planting Density. Environ. Control. Biol. 2012, 50, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.-M.; Ramkumar, G.; Lee, Y. Light quality influences the virulence and physiological responses of Colletotrichum acutatum causing anthracnose in pepper plants. J. Appl. Microbiol. 2013, 115, 509–516. [Google Scholar] [CrossRef]
- Razzak, A.; Ranade, S.S.; Strand, Å.; García-Gil, M.R. Differential response of Scots pine seedlings to variable intensity and ratio of red and far-red light. Plant Cell Environ. 2017, 40, 1332–1340. [Google Scholar] [CrossRef]
- Ouyang, F.; Ma, J.; Wang, J.; Kong, L.; Zhang, H.; Tigabu, M.; Wang, J. Picea species from humid continental and temperate marine climates perform better in monsoonal areas of middle latitudes of China. J. Forestry Res. 2020. [Google Scholar] [CrossRef]
- Yang, Y.-M.; Xu, C.-N.; Wang, B.-M.; Jia, J.-Z. Effects of plant growth regulators on secondary wall thickening of cotton fibres. Plant Growth Regul. 2001, 35, 233–237. [Google Scholar]
- Porra, R.J. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. In Plant Cell Monographs; Springer: Berlin/Heidelberg, Germany, 2006; Volume 20, pp. 633–640. [Google Scholar]
- Sangwan, R.S.; Singh, N.; Plaxton, W.C. Phosphoenolpyruvate Carboxylase Activity and Concentration in the Endosperm of Developing and Germinating Castor Oil Seeds. Plant Physiol. 1992, 99, 445–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.-J.; Cheng, D.-D.; Wang, W.-B.; Gao, H.-Y.; Liu, A.-X.; Li, X.-M.; Meng, Q.-W. Different enhancement of senescence induced by metabolic products of Alternaria alternata in tobacco leaves of different ages. Physiol. Plant. 2010, 138, 164–175. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute Inc. SAS/STAT User’s Guide; SAS Institute Inc.: Cary, NY, USA, 2012. [Google Scholar]
- Grassi, G.; Bagnaresi, U. Foliar morphological and physiological plasticity in Picea abies and Abies alba saplings along a natural light gradient. Tree Physiol. 2001, 21, 959–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Possart, A.; Fleck, C.; Hiltbrunner, A. Shedding (far-red) light on phytochrome mechanisms and responses in land plants. Plant Sci. 2014, 217, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Silvertown, J.; Fenner, M. Seeds: The Ecology of Regeneration in Plant Communities. J. Ecol. 1993, 81, 384. [Google Scholar] [CrossRef]
- Wang, H.; Gu, M.; Cui, J.; Shi, K.; Zhou, Y.; Yu, J. Effects of light quality on CO2 assimilation, chlorophyll-fluorescence quenching, expression of Calvin cycle genes and carbohydrate accumulation in Cucumis sativus. J. Photochem. Photobiol. B Biol. 2009, 96, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.T.; Occhialini, A.; Andralojc, P.J.; Parry, M.A.J.; Hanson, M.R. A faster Rubisco with potential to increase photosynthesis in crops. Nat. Cell Biol. 2014, 513, 547–550. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, L.; Chen, X.; Wu, X.; Xiang, X.; Zhou, J.; Xia, X.; Shi, K.; Yu, J.; Foyer, C.H.; et al. SlHY5 integrates temperature, light and hormone signaling to balance plant growth and cold tolerance. Plant Physiol. 2019, 179, 749–760. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, V.; Pelloux, J.; Podor, M.; Afif, D.; Gerant, D.; Grieu, P.; Dizengremel, P. Carbon fixation in Pinus halepensis submitted to ozone. Opposite response of ribulose-1, 5-bisphosphate carboxylase/oxygenase and phosphoenolpyruvate carboxylase. Physiol. Plantarm 1999, 105, 187–192. [Google Scholar] [CrossRef]
- Berveiller, D.; Damesin, C. Carbon assimilation by tree stems: Potential involvement of phosphoenolpyruvate carboxylase. Trees 2007, 22, 149–157. [Google Scholar] [CrossRef]
- McCree, K. The action spectrum, absorptance and quantum yield of photosynthesis in crop plants. Agric. Meteorol. 1971, 9, 191–216. [Google Scholar] [CrossRef]
- Boardman, N.K. Comparative Photosynthesis of Sun and Shade Plants. Annu. Rev. Plant Physiol. 1977, 28, 355–377. [Google Scholar] [CrossRef]
- Tsuchiya, T.; Ohta, H.; Okawa, K.; Iwamatsu, A.; Shimada, H.; Masuda, T.; Takamiya, K.-I. Cloning of chlorophyllase, the key enzyme in chlorophyll degradation: Finding of a lipase motif and the induction by methyl jasmonate. Proc. Natl. Acad. Sci. USA 1999, 96, 15362–15367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nugent, J.H.A. Oxygenic Photosynthesis. Electron Transfer in Photosystem I and Photosystem II. JBIC J. Biol. Inorg. Chem. 1996, 237, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Pakrasi, H.B. Genetic analysis of the form and function of photosystem I and photosystem II. Annu. Rev. Genet. 1995, 29, 755–776. [Google Scholar] [CrossRef]
- Strasser, B.J. Donor side capacity of Photosystem II probed by chlorophyll a fluorescence transients. Photosynth. Res. 1997, 52, 147–155. [Google Scholar] [CrossRef]
- De Ronde, J.; Cress, W.; Krüger, G.; Strasser, R.J.; Van Staden, J. Photosynthetic response of transgenic soybean plants, containing an Arabidopsis P5CR gene, during heat and drought stress. J. Plant Physiol. 2004, 161, 1211–1224. [Google Scholar] [CrossRef]
- Tóth, S.Z.; Schansker, G.; Kissimon, J.; Kovács, L.; Garab, G.; Strasser, R.J. Biophysical studies of photosystem II-related recovery processes after a heat pulse in barley seedlings (Hordeum vulgare L.). J. Plant Physiol. 2005, 162, 181–194. [Google Scholar] [CrossRef]
- Ruttink, T.; Arend, M.; Morreel, K.; Storme, V.; Rombauts, S.; Fromm, J.; Bhalerao, R.P.; Boerjan, W.; Rohde, A. A molecular timetable for apical bud formation and dormancy induction in Poplar. Plant Cell 2007, 19, 2370–2390. [Google Scholar] [CrossRef] [Green Version]
- Müller-Xing, R.; Xing, Q.; Goodrich, J. Footprints of the sun: Memory of UV and light stress in plants. Front. Plant Sci. 2014, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Son, K.-H.; Oh, M.-M. Leaf shape, growth, and antioxidant phenolic compounds of two lettuce cultivars grown under various combinations of blue and red light-emitting diodes. HortScience 2013, 48, 988–995. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Mane | Gene Id | Primer F (5’–3’) | Primer R (5’–3’) |
|---|---|---|---|
| rbcl | MA_103636g0010 | CCACTTTAGGGTTTGTTGATCTAC | CTCGGTCAGAGCAGGCATA |
| PEPC1 | MA_176143g0010 | CGGGATGATGAATTGACAGCAGATG | CCATTAGTGCCACAGGTGTAAGGTT |
| PGK1 | MA_92421g0010 | AAGCCGTGTCATCGTCGTCTACT | AGCCGAATACGAACCTGGACTTGA |
| RCA | MA_10433855g0010 | CGGAGAACTGGAGAGTGGAGATG | CACGAGGATTGTCTTGCTTGTTGT |
| CLH2 | MA_5475932g0010 | GCAGCAGCGTGAGGAGATATG | GGAAGCGATGTGTTGGAGAAGTT |
| CHS | MA_10426264g0020 | GGCATTCAGGAAGGCTCAGAGA | GGCACCTCCACCACAACCAT |
| phyA | MA_6809p0010 | GCAGCAGCGTTCCATCTGAGAC | CACCACCAACAACATCCAGCATCT |
| phyB | MA_6809g0020 | CAGCAGCCGCAGAACAGAAG | GAAGCATATCGCAGAGCACAGTT |
| phyC | MA_6809g0030 | GCATGACTCGGTTGGAATCGTGAA | GCCCTGTAATCGCCTTGAAGATGAG |
| HY5 | MA_41006g0010 | ATGCGTGAACGAGAGTGATGATGAT | GTTCTGCCTCTTCTCCTCTGACTG |
| COP1-2 | MA_10433141g0010 | TCACGCACAGAGCCGACCAT | GCTGAACCAACCGCAACATAGTT |
| PIF3 | MA_29186g0010 | ACTGGTGTTGGTCTTGGTATGG | GACTGATGTTGGCAAGCAATGTAT |
| actin | GTGCCTGCTATGTATGTTGCCATTC | GCTTCTCCTTCACATCACGGACAAT |
| Traits | Light Quality | Error | Traits | Light Quality | Error |
|---|---|---|---|---|---|
| Stem height | 90.85 ** | 34.47 | Chlorophyll | 0.01 ** | 0.04 |
| Root collar diameter | 9.19 * | 1.08 | Chlorophyll/fresh weight | 18.61 * | 8.71 |
| Current-year shoot length | 57.94 ** | 8.45 | Pn | 11.46 ** | 2.38 |
| Leaf area | 0.92 * | 0.23 | Gs | 0.02 ** | 0.09 × 10−2 |
| Leaf dry weight | 13.25 × 10−4 * | 1.32 × 10−4 | Ci | 5555.20 ** | 265.27 |
| SLA | 11.91 × 10−6 * | 4.64 × 10−6 | Tr | 13.24 ** | 0.44 |
| GAs | 6.36 ** | 0.10 | Rubisco | 1.46 × 10−4 * | 0.43 × 10−4 |
| IAA | 26523 ** | 109 | PEPC | 7.91 * | 2.34 |
| ZR | 0.93 ** | 0.11 | PEPC/Rubisco | 0.11 ** | 0.02 |
| ABA | 3030 ** | 46.93 |
| Chlorophyll a Fluorescence Parameters | Lighting 1 h | Lighting 3 h | Lighting 9 h | |||
|---|---|---|---|---|---|---|
| Light Quality | Error | Light Quality | Error | Light Quality | Error | |
| Vj | 8.06 × 10−3 * | 2.95 × 10−3 | 3.63 × 10−3 * | 1.47 × 10−3 | 9.97 × 10−3 ** | 2.37 × 10−3 |
| WK | 2.71 × 10−3 | 4.01 × 10−3 | 21.17 × 10−3 ** | 2.93 × 10−3 | 7.83 × 10−3 * | 3.61 × 10−3 |
| Fv/Fm | 0.32 × 10−3 ** | 0.11 × 10−3 | 0.48 × 10−3 ** | 0.15 × 10−3 | 0.16 × 10−3 * | 0.07 × 10−3 |
| ψo | 8.07 × 10−3 * | 2.95 × 10−3 | 3.63 × 10−3 * | 1.47 × 10−3 | 9.97 × 10−3 ** | 2.37 × 10−3 |
| φEo | 4.79 × 10−3 * | 2.07 × 10−3 | 3.02 × 10−3 * | 1.18 × 10−3 | 6.69 × 10−3 ** | 1.62 × 10−3 |
| Hour | Light Quality | Vj | WK | Fv/Fm (φPo) | ψO | φEo |
|---|---|---|---|---|---|---|
| 1 | B | 0.47 (0.05) ab | 0.41 (0.07) a | 0.8332 (0.01) b | 0.52 (0.05) ab | 0.44 (0.04) ab |
| R | 0.45 (0.03) b | 0.39 (0.05) a | 0.8350 (0.01) b | 0.57 (0.02) a | 0.47 (0.03) a | |
| FR | 0.48 (0.02) a | 0.42 (0.05) a | 0.8426 (0.01) a | 0.52 (0.02) b | 0.43 (0.02) b | |
| CK | 0.45 (0.06) b | 0.40 (0.06) a | 0.8344 (0.01) b | 0.55 (0.06) ab | 0.45 (0.04) ab | |
| 3 | B | 0.51 (0.03) a | 0.44 (0.04) a | 0.8339 (0.01) b | 0.48 (0.03) b | 0.40 (0.03) b |
| R | 0.48 (0.04) b | 0.37 (0.03) b | 0.8446 (0.01) a | 0.51 (0.03) a | 0.43 (0.03) a | |
| FR | 0.51 (0.04) a | 0.43 (0.05) a | 0.8409 (0.01) ab | 0.48 (0.04) b | 0.40 (0.03) b | |
| CK | 0.49 (0.04) ab | 0.37 (0.06) b | 0.8328 (0.01) b | 0.50 (0.03) ab | 0.41 (0.03) ab | |
| 9 | B | 0.48 (0.05) ab | 0.38 (0.05) ab | 0.8369 (0.01) ab | 0.52 (0.05) ab | 0.44 (0.04) ab |
| R | 0.45 (0.03) b | 0.36 (0.04) b | 0.8378 (0.01) ab | 0.55 (0.03) a | 0.46 (0.03) a | |
| FR | 0.51 (0.03) a | 0.42 (0.06) a | 0.8432 (0.01) a | 0.49 (0.03) b | 0.41 (0.03) b | |
| CK | 0.51 (0.05) a | 0.38 (0.07) ab | 0.8362 (0.01) b | 0.49 (0.05) b | 0.41 (0.04) b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
OuYang, F.; Ou, Y.; Zhu, T.; Ma, J.; An, S.; Zhao, J.; Wang, J.; Kong, L.; Zhang, H.; Tigabu, M. Growth and Physiological Responses of Norway Spruce (Picea abies (L.) H. Karst) Supplemented with Monochromatic Red, Blue and Far-Red Light. Forests 2021, 12, 164. https://doi.org/10.3390/f12020164
OuYang F, Ou Y, Zhu T, Ma J, An S, Zhao J, Wang J, Kong L, Zhang H, Tigabu M. Growth and Physiological Responses of Norway Spruce (Picea abies (L.) H. Karst) Supplemented with Monochromatic Red, Blue and Far-Red Light. Forests. 2021; 12(2):164. https://doi.org/10.3390/f12020164
Chicago/Turabian StyleOuYang, Fangqun, Yang Ou, Tianqin Zhu, Jianwei Ma, Sanping An, Jun Zhao, Junhui Wang, Lisheng Kong, Hanguo Zhang, and Mulualem Tigabu. 2021. "Growth and Physiological Responses of Norway Spruce (Picea abies (L.) H. Karst) Supplemented with Monochromatic Red, Blue and Far-Red Light" Forests 12, no. 2: 164. https://doi.org/10.3390/f12020164
APA StyleOuYang, F., Ou, Y., Zhu, T., Ma, J., An, S., Zhao, J., Wang, J., Kong, L., Zhang, H., & Tigabu, M. (2021). Growth and Physiological Responses of Norway Spruce (Picea abies (L.) H. Karst) Supplemented with Monochromatic Red, Blue and Far-Red Light. Forests, 12(2), 164. https://doi.org/10.3390/f12020164