
Seasonal Variations in Carbon, Nitrogen, and Phosphorus Stoichiometry of a Robinia pseudoacacia Plantation on the Loess Hilly Region, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Data Collection
2.4. Statistical Analysis
3. Results
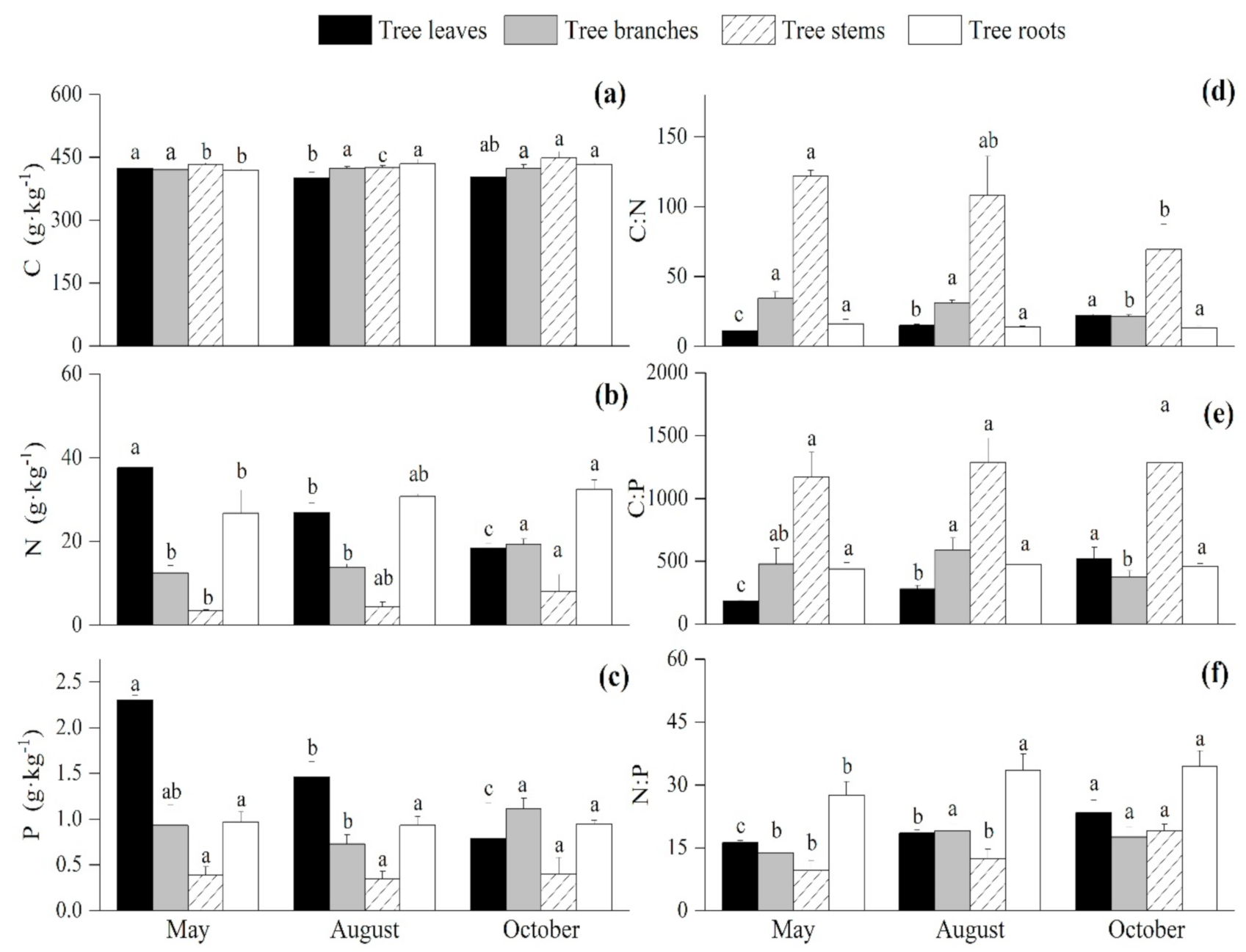
3.1. Seasonal Variations of the C:N:P Stoichiometry in the Arbor Layer
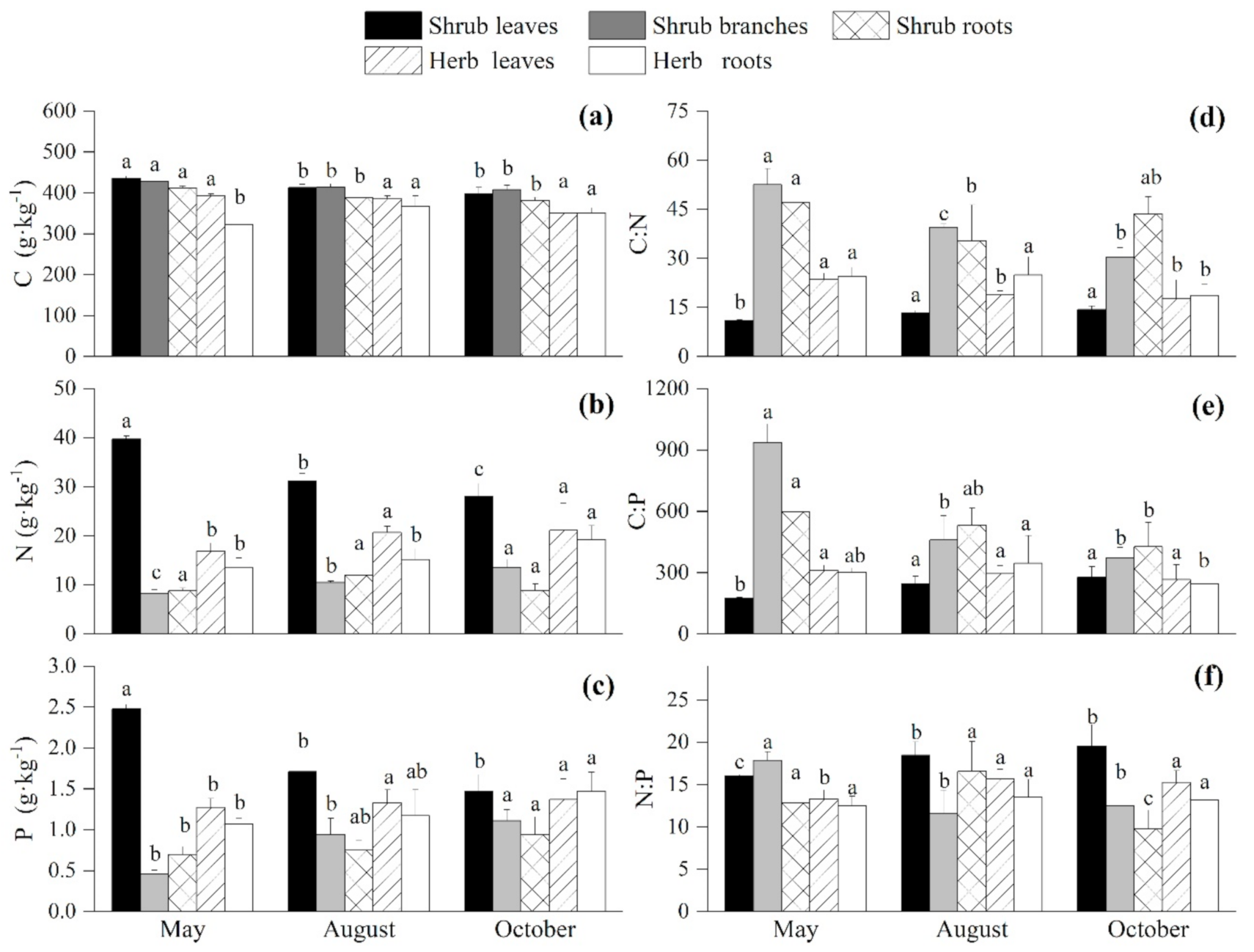
3.2. Seasonal Variations of the C:N:P Stoichiometry in the Undergrowth Layer
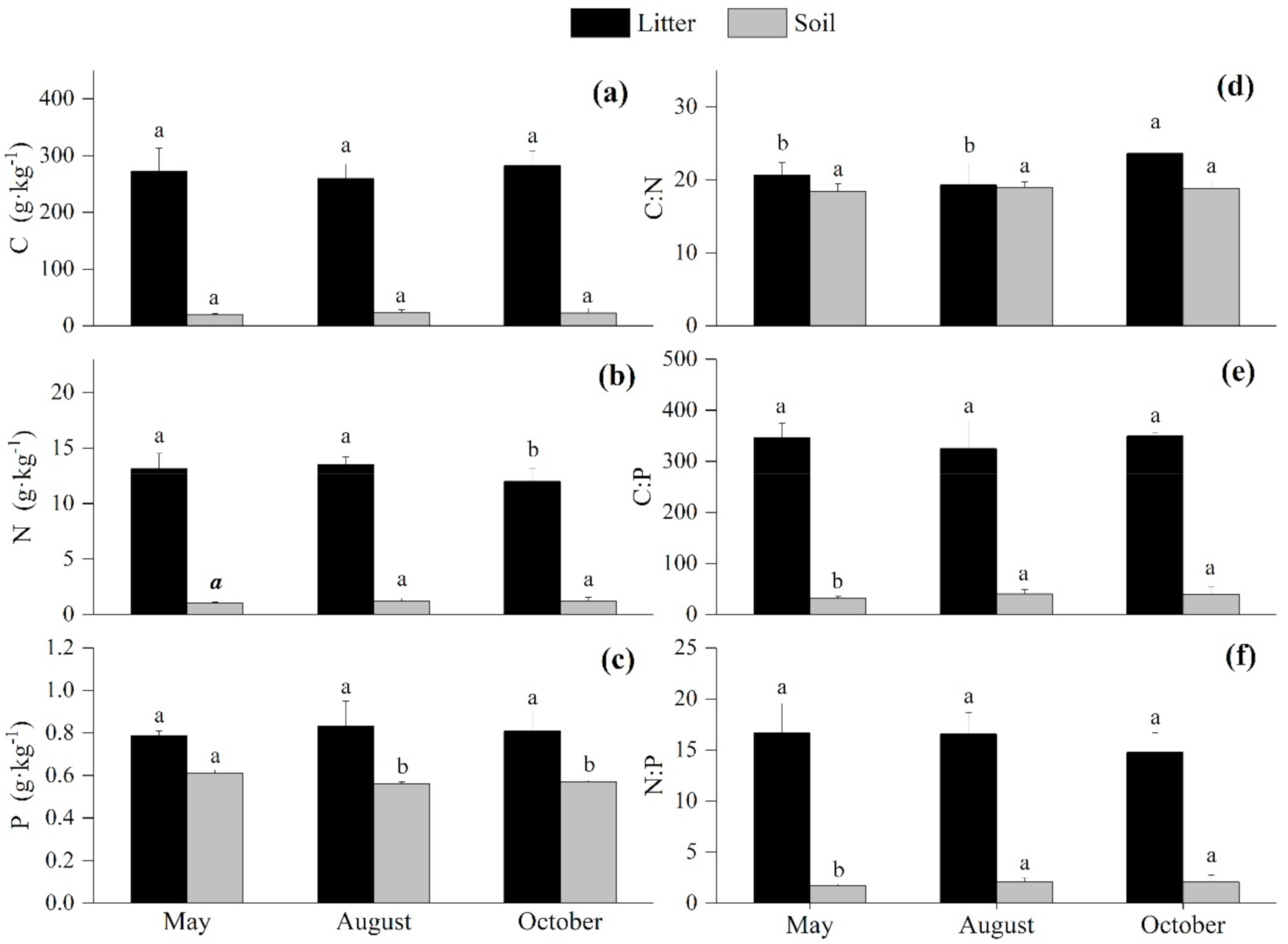
3.3. Seasonal Variations of the C:N:P Stoichiometry in the Litter and Soil
3.4. Correlations of the C:N:P Stoichiometry Among the Plants, Litter, and Soil
4. Discussion
4.1. Seasonal Variations of the C, N, and P Contents and Ratios of the Plants
4.2. Seasonal Variation of the C:N:P Stoichiometry in the Litter and Soil
4.3. Correlation Analysis of Plants, Litter and Soil C, N, P Stoichiometry
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elser, J.J.; Hamilton, A. Stoichiometry and the new biology: The future is now. PLoS Biol. 2007, e181. [Google Scholar] [CrossRef]
- Han, W.X.; Fang, J.Y.; Guo, D.L.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 358–377. [Google Scholar] [CrossRef]
- Klausmeier, C.A.; Litchman, E.; Daufresne, T.; Levin, S.A. Optimal nitrogen-to-phosphorus stoichiometry of phytoplankton. Nature 2004, 429, 171–174. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. The role of plants in the effects of global change on nutrient availability and stoichiometry in the Plant-Soil System. Plant Physiol. 2012, 160, 1741–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Liu, B.; Gao, X.P.; Li, X.Y.; Li, C.D. Nitrogen and phosphorus addition differentially affect plant ecological stoichiometry in desert grassland. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- McGroddy, M.E.; Daufresne, T.; Hedin, L.O. Scaling of C:N:P stoichiometry in forests worldwide: Implications of terrestrial redfield-type ratios. Ecology 2004, 85, 2390–2401. [Google Scholar] [CrossRef]
- Maas-Hebner, K.G.; Emmingham, W.H.; Larson, D.J.; Chan, S.S. Establishment and growth of native hardwood and conifer seedlings underplanted in thinned Douglas-fir stands. For. Ecol. Manag. 2005, 208, 331–345. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.R.; An, S.S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. Catena 2018, 166, 328–338. [Google Scholar] [CrossRef]
- Berg, B. Litter decomposition and organic matter turnover in northern forest soils. For. Ecol. Manag. 2000, 133, 13–22. [Google Scholar] [CrossRef]
- Bai, X.J.; Wang, B.R.; An, S.S.; Zeng, Q.C.; Zhang, H.X. Response of forest species to C:N:P in the plant-litter-soil system and stoichiometric homeostasis of plant tissues during afforestation on the Loess Plateau, China. Catena 2019, 183, 104186. [Google Scholar] [CrossRef]
- Manzoni, S.; Jackson, R.B.; Trofymow, J.A.; Porporato, A. The global stoichiometry of litters nitrogen mineralization. Science 2008, 321, 684–686. [Google Scholar] [CrossRef]
- Gao, D.X.; Zhang, W.; Ren, C.J.; Dai, Y.Y.; Qiao, W.J.; Chen, Z.X.; Yang, G.H.; Han, X.H. Ecological stoichiometry characteristics of soil and leaves during the recovery process of typical vegetation on the Loess Plateau. Acta Ecol. Sin. 2019, 39, 3622–3630. (In Chinese) [Google Scholar]
- Fan, H.B.; Wu, J.P.; Liu, W.F.; Yuan, Y.H.; Hu, L.; Cai, Q.K. Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant Soil 2015, 392, 127–138. [Google Scholar] [CrossRef]
- Führer, E. Forest functions, ecosystem stability and management. For. Ecol. Manag. 2000, 132, 29–38. [Google Scholar] [CrossRef]
- Loreau, M.; De Mazancourt, C.; Duffy, E. Biodiversity and ecosystem stability: A synthesis of underlying mechanisms. Ecol. Lett. 2013, 16, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Lü, X.; Reed, S.; Hou, S.L.; Hu, Y.Y.; Wei, H.W.; Lü, F.M.; Cui, Q.; Han, X.G. Temporal variability of foliar nutrients: Responses to nitrogen deposition and prescribed fire in a temperate steppe. Biogeochemistry 2017, 133, 295–305. [Google Scholar] [CrossRef]
- Zhao, Q.Q.; Bai, J.H.; Liu, Q.; Lu, Q.Q.; Gao, Z.Q.; Wang, J.J. Spatial and seasonal variations of soil carbon and nitrogen content and stock in a Tidal Salt Marsh with Tamarix chinensis, China. Wetlands 2016, 36, 145–152. [Google Scholar] [CrossRef]
- Ågren, G.I. Stoichiometry and nutrition of plant growth in natural communities. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 153–170. [Google Scholar] [CrossRef]
- Yang, Y.H.; Luo, Y.Q. Carbon:nitrogen stoichiometry in forest ecosystems during stand development. Glob. Ecol. Biogeogr. 2011, 20, 354–361. [Google Scholar] [CrossRef]
- Wright, I.J.; Westoby, M. Nutrient concentration, resorption and lifespan: Leaf traits of Australian sclerophyll species. Funct. Ecol. 2003, 17, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Grigal, D.F.; Ohmann, L.F.; Brander, R.B. Seasonal dynamics of tall shrubs in northeastern Minnesota: Biomass and nutrient element changes. For. Sci. 1976, 195–208. [Google Scholar] [CrossRef]
- Kerkhoff, A.J.; Enquist, B.J.; Elser, J.J.; Fagan, W.F. Plant allometry, stoichiometry and the temperature-dependence of primary productivity. Glob. Ecol. Biogeogr. 2005, 14, 585–598. [Google Scholar] [CrossRef]
- Liu, F.D.; Liu, Y.H.; Wang, G.M.; Song, Y.; Liu, Q.; Li, D.S.; Mao, P.L.; Zhang, H. Seasonal Variations of C:N:P stoichiometry and their trade-offs in different organs of Suaeda salsa in Coastal Wetland of Yellow River Delta, China. PLoS ONE 2015, 10, e138169. [Google Scholar] [CrossRef]
- Jing, L.; Li, J.P.; Zhang, Y.; Ma, H.Y.; Li, J. Study on dry soil layers under different land-use systerms in the Loess Plateau. Pratacult. Sci. 2018, 35, 1829–1835. (In Chinese) [Google Scholar]
- Wu, D.Y.; Jiao, J.Y.; Yu, W.J.; Xu, H.Y.; Chen, Y.X. Respone characteristics of Locust growth, understory vegetation and soil moisture on planting density in northern Shaanxi. Acta Bot. Boreali-Occident. Sin. 2017, 37, 346–355. (In Chinese) [Google Scholar]
- Wu, Z.B.; Yang, X.M. Artificial Robinia peseudoacacia growth and soil moisture variation in Loessial Hilly-Gully region. Res. Soil Water Conserv. 2004, 11, 117–120. (In Chinese) [Google Scholar]
- Cao, Y.; Chen, Y.M. Coupling of plant and soil C:N:P stoichiometry in black locust (Robinia pseudoacacia) plantations on the Loess Plateau, China. Trees 2017, 31, 1559–1570. [Google Scholar] [CrossRef]
- Du, M.; Feng, H.; Zhang, L.; Pei, S.; Tang, X. Variations in carbon, nitrogen and phosphorus stoichiometry during a growth season within a Platycladus orientalis plantation. Pol. J. Environ. Stud. 2020, 29. [Google Scholar] [CrossRef]
- Yeomans, J.C.; Bremner, J.M. A rapid and precise method for routine determination of organic carbon in soil. Commun. Soil Sci. Plant Anal. 1988, 19, 1467–1476. [Google Scholar] [CrossRef]
- Zhang, G.Q.; Zhang, P.; Peng, S.Z.; Chen, Y.M.; Cao, Y. The coupling of leaf, litter, and soil nutrients in warm temperate forests in northwestern China. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, J.A.; Allen, S.E. A wet oxidation procedure suitable for the determination of nitrogen and mineral nutrients in biological material. Commun. Soil Sci. Plant Anal. 2009, 6, 1–11. [Google Scholar] [CrossRef]
- Thomas, S.C.; Martin, A.R. Carbon content of tree tissues: A synthesis. Forests 2012, 3, 332–352. [Google Scholar] [CrossRef] [Green Version]
- Abbas, M.; Ebeling, A.; Oelmann, Y.; Ptacnik, R.; Roscher, C.; Weigelt, A.; Weisse, W.W.; Wilcke, W.; Hillebrand, H. Biodiversity effects on plant stoichiometry. PLoS ONE 2013, 8, e58179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.L.; Crabbe, M.J.C.; Xu, F.L.; Wang, W.L.; Niu, R.L.; Gao, X.; Zhang, P.; Chen, H.K. Seasonal Variations in carbon, nitrogen and phosphorus concentrations and C:N:P stoichiometry in the leaves of differently aged Larix principis-rupprechtii Mayr. Plantations. Forests 2017, 8, 373. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Wu, J.G.; ClarK, C.M.; Pan, Q.M.; Zhang, L.X.; Chen, S.P.; Wang, Q.B.; Han, X.Q. Grazing alters ecosystem functioning and C:N:P stoichiometry of grasslands along a regional precipitation gradient. J. Appl. Ecol. 2012, 49, 1204–1215. [Google Scholar] [CrossRef]
- Yang, J.J.; Zhang, X.R.; MA, L.S.; Chen, Y.A.; Dang, T.H.; An, S.S. Ecological stoichiometric relationships between components of Robinia pseudoacacia forest in loess plateau. Acta Pedol. Sin. 2014, 51, 133–142. (In Chinese) [Google Scholar]
- Ye, Y.S.; Liang, X.Q.; Chen, Y.X.; Li, L.; Ji, Y.J.; Zhu, C.Y. Carbon, Nitrogen and phosphorus accumulation and partitioning, and C:N:P stoichiometry in late-season rice under different water and nitrogen managements. PLoS ONE 2014, 9, e101776. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.N.; Lu, J.Y.; Yang, H.M.; Zhang, X.; Luo, C.L.; Zhao, Y.X. Resorption of nitrogen, phosphorus and potassium from leaves of lucerne stands of different ages. Plant Soil. 2014, 383, 301–312. [Google Scholar] [CrossRef]
- Sardans, J.; Rivas-Ubach, A.; Peuelas, J. Factors affecting nutrient concentration and stoichiometry of forest trees in Catalonia (NE Spain). For. Ecol. Manag. 2011, 262, 2024–2034. [Google Scholar] [CrossRef]
- Yu, Q.; Wu, H.H.; He, N.P.; Lu, X.T.; Wang, Z.P.; Elser, J.J. Testing the Growth Rate Hypothesis in vascular plants with above- and below-ground biomass. PLoS ONE 2012, 7, e32162. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Zhuang, W.W.; Li, J. Dynamic analysis of stoichiometric ratios of C, N, and P during different growth periods in four herbaceous plants from Gurbantünggüt Desert. Plant Sci. J. 2019, 37, 356–366. (In Chinese) [Google Scholar]
- Santa Regina, I.; Rico, M.; Gallego, M.H.A. Seasonal variation in nutrient concentration in leaves and branches of Quercus pyrenaica. J. Veg. Sci. 1997, 8, 651–654. [Google Scholar] [CrossRef]
- Townsend, A.R.; Cleveland, C.C.; Asner, G.P.; Bustamante, M.M. Controls over foliar N:P ratios in tropical rain forests. Ecology 2007, 88, 107–118. [Google Scholar] [CrossRef]
- Aerts, R. Nutrient resorption from senescing leaves of perennials: Are there general patterns. J. Ecol. 1996, 84, 597–608. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, J.; Biswas, A.; Cao, J.; Qi, X. Seasonal dynamics of leaf stoichiometry of phragmites australis: A case study from Yangguan Wetland, Dunhuang, China. Plants 2020, 9, 1323. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.; Fagan, W.F.; Markow, T.A.; Cotner, J.B.; Harrison, J.F.; Hobbie, S.E.; Odell, G.M.; Weider, L.W. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.N.; Gao, P.; Li, C.; Wang, R.J.; Niu, X.; Wang, B. Ecological stoichiometry characteristics of the leaf-litter-soil continuum of Quercus acutissima Carr. and Pinus densiflora Sieb. in Northern China. Environ. Earth Sci. 2019, 78. [Google Scholar] [CrossRef]
- Chen, C.; Wang, G.J.; Zhao, Y.; Zhou, G.X.; Li, L.; Gao, J.Q. Seasonal dynamics and allometric growth relationships of C, N and P stoichiometry in the organs of Cunninghamia lanceolata from Huitong. Acta Ecol. Sin. 2016, 36, 7614–7623. [Google Scholar] [CrossRef] [Green Version]
- Forja, J.M.; Blasco, J.; Gómez-Parra, A. Spatial and seasonal variation of in situ benthic fluxes in the bay of Cadiz (South-west Spain). Estuar. Coast. Shelf Sci. 1994, 39, 127–141. [Google Scholar] [CrossRef]
- Waive, J.; Larsson, U. Seasonal changes in Baltic Sea seston stoichiometry: The influence of diazotrophic cyanobacteria. Mar. Ecol. Prog. 2010, 13–25. [Google Scholar] [CrossRef]
- Li, H.L.; Crabbe, M.J.C.; Xu, F.L.; Wang, W.L.; Ma, L.H.; Niu, R.L.; Gao, X.; Li, X.X.; Zhang, P.; Ma, X.; et al. Seasonal variations in carbon, nitrogen and phosphorus concentrations and C:N:P stoichiometry in different organs of a Larix principis-rupprechtii Mayr. plantation in the Qinling Mountains, China. PLoS ONE 2017, 12, e185163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grace, J. Toward models of resource allocation by plants. Plant Resour. Alloc. 1997, 279–291. [Google Scholar] [CrossRef]
- Vitousek, P. Nutrient cycling and nutrient use efficiency. Am. Nat. 1982, 119, 553–572. [Google Scholar] [CrossRef]
- Koerselman, W.; Meuleman, A.F.M. The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- Tian, H.Q.; Chen, G.S.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Hatton, P.J.; Castanha, C.; Torn, M.S.; Bird, J.A. Litter type control on soil C and N stabilization dynamics in a temperate forest. Glob. Chang. Biol. 2015, 1358–1367. [Google Scholar] [CrossRef]
- Kang, H.; Freeman, C. Soil enzyme analysis for leaf litter decomposition in Global Wetlands. Commun. Soil Sci. Plant Anal. 2009, 40, 3323–3334. [Google Scholar] [CrossRef]
- Ren, S.J.; Cao, M.K.; Tao, B.; Li, K.R. The effects of nitrogen limitation on terrestrial ecosystem carbon cycle: A review. Prog. Geogr. 2006, 25, 58–67. (In Chinese) [Google Scholar]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio”for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Yang, Y.H.; Fang, J.Y.; Guo, D.L.; Ji, C.J.; Ma, W.H. Vertical patterns of soil carbon, nitrogen and carbon: Nitrogen stoichiometry in Tibetan grasslands. Biogeosci. Discuss. 2010, 7, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Neufeldt, H.; Da Silva, J.E.D.; Ayarza, M.A.; Zech, W. Land-use effects on phosphorus fractions in Cerrado oxisols. Biol. Fertil. Soils 2000, 31, 30–37. [Google Scholar] [CrossRef]
- Post, W.M.; Pastor, J.; Zinke, P.J.; Stangenberger, A.G. Global patterns of soil nitrogen storage. Nature 1985, 317, 613–616. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Vitousek, P.M. Nutrient limitation of decomposition in Hawaiian forests. Ecology 2000, 81, 1867–1877. [Google Scholar] [CrossRef]
- Wang, Z.; Zheng, F. Impact of vegetation succession on leaf-litter-soil C:N:P stoichiometry and their intrinsic relationship in the Ziwuling Area of China’s Loess Plateau. J. For. Res. 2020. [Google Scholar] [CrossRef]
- Walker, T.W.; Adams, A.F.R. Studies on soil organic matter. Soil Sci. 1959, 85, 307–318. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | C | N | P | C:N | C:P | N:P | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AV/g·kg−1 | CV/% | AV/g·kg−1 | CV/% | AV/g·kg−1 | CV/% | AV | CV/% | AV | CV/% | AV | CV/% | |
| TL | 409.02 ± 19.82 | 4.85 | 27.61 ± 8.15 | 29.53 | 1.52 ± 0.64 | 42.15 | 16.08 ± 4.64 | 28.87 | 326.97 ± 153.01 | 46.8 | 19.44 ± 3.45 | 17.74 |
| TB | 419.47 ± 8.37 | 1.99 | 15.18 ± 3.28 | 21.64 | 0.93 ± 0.22 | 23.86 | 28.9 ± 6.22 | 21.54 | 481.16 ± 127.44 | 26.49 | 16.75 ± 2.97 | 17.72 |
| TS | 435.13 ± 10.05 | 3.13 | 5.28 ± 3.10 | 58.8 | 0.38 ± 0.12 | 31.78 | 99.79 ± 34.82 | 34.89 | 1248.53 ± 350.31 | 28.06 | 13.71 ± 4.54 | 33.14 |
| TR | 428.32 ± 13.64 | 2.35 | 30.01 ± 4.11 | 13.71 | 0.95 ± 0.09 | 9.13 | 14.55 ± 2.14 | 14.73 | 455.62 ± 47.28 | 10.38 | 31.79 ± 4.72 | 14.83 |
| SL | 415.7 ± 18.57 | 4.47 | 32.97 ± 5.31 | 16.1 | 1.88 ± 0.47 | 24.97 | 12.85 ± 1.61 | 12.53 | 233.2 ± 55.74 | 23.9 | 18.01 ± 2.95 | 16.39 |
| SB | 416.68 ± 12.07 | 2.9 | 10.76 ± 2.46 | 22.83 | 0.84 ± 0.31 | 37.11 | 40.77 ± 9.80 | 24.05 | 589.99 ± 267.43 | 45.33 | 14 ± 3.62 | 25.84 |
| SR | 393.69 ± 15.82 | 4.02 | 9.87 ± 2.53 | 25.67 | 0.8 ± 0.18 | 22.9 | 41.99 ± 8.66 | 20.64 | 519.23 ± 116.37 | 22.41 | 13.07 ± 4.91 | 37.53 |
| HL | 376.67 ± 31.24 | 8.29 | 19.57 ± 3.83 | 19.55 | 1.32 ± 0.18 | 13.68 | 19.96 ± 4.24 | 21.25 | 290.74 ± 51.68 | 17.77 | 14.75 ± 1.56 | 10.59 |
| HR | 347.16 ± 26.82 | 7.73 | 15.97 ± 3.36 | 21.05 | 1.24 ± 0.28 | 22.76 | 22.64 ± 4.91 | 21.68 | 297.98 ± 91.36 | 30.66 | 13.04 ± 1.47 | 11.27 |
| LI | 270.96 ± 50.37 | 18.59 | 12.86 ± 2.15 | 16.76 | 0.81 ± 0.09 | 10.63 | 21.19 ± 2.72 | 12.84 | 340 ± 79.50 | 23.38 | 15.99 ± 2.74 | 17.14 |
| SO | 21.31 ± 5.95 | 27.9 | 1.13 ± 0.27 | 23.57 | 0.58 ± 0.02 | 3.89 | 18.72 ± 0.87 | 4.63 | 36.64 ± 10.59 | 28.91 | 1.94 ± 0.48 | 24.72 |
| Plants | Components | C | N | P |
|---|---|---|---|---|
| Tree | Leaves–Branches | 0.404 * | −0.847 ** | −0.587 ** |
| Leaves-Stems | 0.532 ** | −0.613 ** | −0.183 | |
| Leaves-Roots | −0.352 | −0.597 ** | −0.189 | |
| Branches-Stems | −0.298 | 0.724 ** | 0.602 ** | |
| Branches-Roots | −0.142 | 0.557 ** | 0.458 * | |
| Stems-Roots | 0.030 | 0.085 | 0.421 * | |
| Shrub | Leaves-Branch | 0.893 ** | −0.728 ** | −0.835 ** |
| Leaves-Root | 0.761 ** | −0.118 | −0.294 | |
| Branches-Roots | 0.680 ** | 0.142 | 0.439 * | |
| Herb | Leaves-Roots | −0.103 | 0.684 ** | 0.717 ** |
| Component | C | N | P | C:N | C:P | N:P |
|---|---|---|---|---|---|---|
| TL-SL | 0.613 ** | 0.919 ** | 0.928 ** | 0.759 ** | 0.806 ** | 0.794 ** |
| TL-AH | −0.299 | −0.494 ** | −0.24 | −0.582 ** | −0.539 ** | 0.279 |
| TL-LI | 0.44 * | 0.492 ** | −0.092 | 0.542 ** | 0.066 | −0.173 |
| TL-SO | −0.523 ** | −0.312 | 0.785 ** | 0.221 | 0.182 | 0.082 |
| SL-AH | 0.143 | −0.345 | −0.014 | −0.213 | −0.241 | 0.202 |
| SL-LI | 0.241 | 0.21 | −0.044 | 0.547 ** | 0.367 | 0.208 |
| SL-SO | −0.586 ** | −0.232 | 0.858 ** | 0.05 | −0.126 | −0.263 |
| AH-LI | −0.334 | −0.116 | 0.629 ** | 0.074 | 0.377 | −0.031 |
| AH-SO | 0.193 | 0.771 ** | −0.15 | −0.318 | −0.368 | 0.572 ** |
| LI-SO | −0.547 ** | −0.554 ** | −0.073 | 0.09 | −0.404 * | −0.512 ** |
| Item | Area | C/(g·kg−1) | N/(g·kg−1) | P/(g·kg−1) | C:N | C:P | N:P | Date Source |
|---|---|---|---|---|---|---|---|---|
| Tree leaves | The Loess Plateau | 409.02 ± 19.8 | 27.61 ± 8.1 | 1.52 ± 0.6 | 16.08 ± 4.4 | 326.97 ± 86 | 19.44 ± 3.4 | This study |
| China | - | 20.2 ± 8.4 | 1.46 ± 1.0 | - | - | 13.83 ± 9.3 | Han | |
| Global | 464.0 ± 3.2 | 20.6 ± 1.2 | 1.99 ± 0.1 | 22.5 ± 1.9 | 232.0 ± 72.4 | 12.7 ± 10.1 | Elser | |
| Global | 461.3 ± 72.2 | 20.1 ± 8.7 | 1.77 ± 1.1 | 23.8 ± 17.3 | 300.9 ± 236.8 | 13.8 ± 9.5 | Reich and Oleksyn | |
| Litter | The Loess Plateau | 270.96 ± 50.3 | 12.86 ± 2.1 | 0.81 ± 0.1 | 21.19 ± 2.7 | 340.0 ± 79.5 | 15.99 ± 2.7 | This study |
| China | 508.16 ± 28.2 | 10.85 ± 1.9 | 0.46 ± 0.2 | 54.1 ± 16.4 | 1355.74 ± 80.1 | 25.47 ± 2.4 | Wang | |
| Global | 467.7 ± 2.7 | 10.0 ± 0.1 | 0.7 ± 0.0 | 52.9 ± 1.3 | 1183.0 ± 70 | 19.2 ± 0.5 | Yuan | |
| Global | - | 10.93 ± 6.5 | 0.85 ± 0.7 | - | - | 18.32 ± 11.2 | Kang | |
| Soil | The Loess Plateau | 21.31 ± 5.9 | 1.13 ± 0.3 | 0.58 ± 0.0 | 18.72 ± 0.8 | 36.64 ± 10.5 | 1.94 ± 0.4 | This study |
| China | 11.12 ± 1.6 | 1.07 ± 0.1 | 2.05 ± 0.7 | 11.9 ± 0.2 | 60.0 ± 1.1 | 5.1 ± 0.1 | Tian | |
| Global | - | - | - | 14.3 ± 0.5 | 186.0 ± 12.9 | 13.1 ± 0.8 | Cleveland |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, C.; Qiao, Y.; Cao, Y.; Chen, Y.; Wu, X.; Xue, W. Seasonal Variations in Carbon, Nitrogen, and Phosphorus Stoichiometry of a Robinia pseudoacacia Plantation on the Loess Hilly Region, China. Forests 2021, 12, 214. https://doi.org/10.3390/f12020214
Dong C, Qiao Y, Cao Y, Chen Y, Wu X, Xue W. Seasonal Variations in Carbon, Nitrogen, and Phosphorus Stoichiometry of a Robinia pseudoacacia Plantation on the Loess Hilly Region, China. Forests. 2021; 12(2):214. https://doi.org/10.3390/f12020214
Chicago/Turabian StyleDong, Congguo, Yuning Qiao, Yang Cao, Yunming Chen, Xu Wu, and Wenyan Xue. 2021. "Seasonal Variations in Carbon, Nitrogen, and Phosphorus Stoichiometry of a Robinia pseudoacacia Plantation on the Loess Hilly Region, China" Forests 12, no. 2: 214. https://doi.org/10.3390/f12020214
APA StyleDong, C., Qiao, Y., Cao, Y., Chen, Y., Wu, X., & Xue, W. (2021). Seasonal Variations in Carbon, Nitrogen, and Phosphorus Stoichiometry of a Robinia pseudoacacia Plantation on the Loess Hilly Region, China. Forests, 12(2), 214. https://doi.org/10.3390/f12020214