
Captures of Emerald Ash Borer (Agrilus planipennis) Adults in Post-Invasion White Ash Sites with Varying Amounts of Live Phloem

Abstract
:1. Introduction
2. Methods
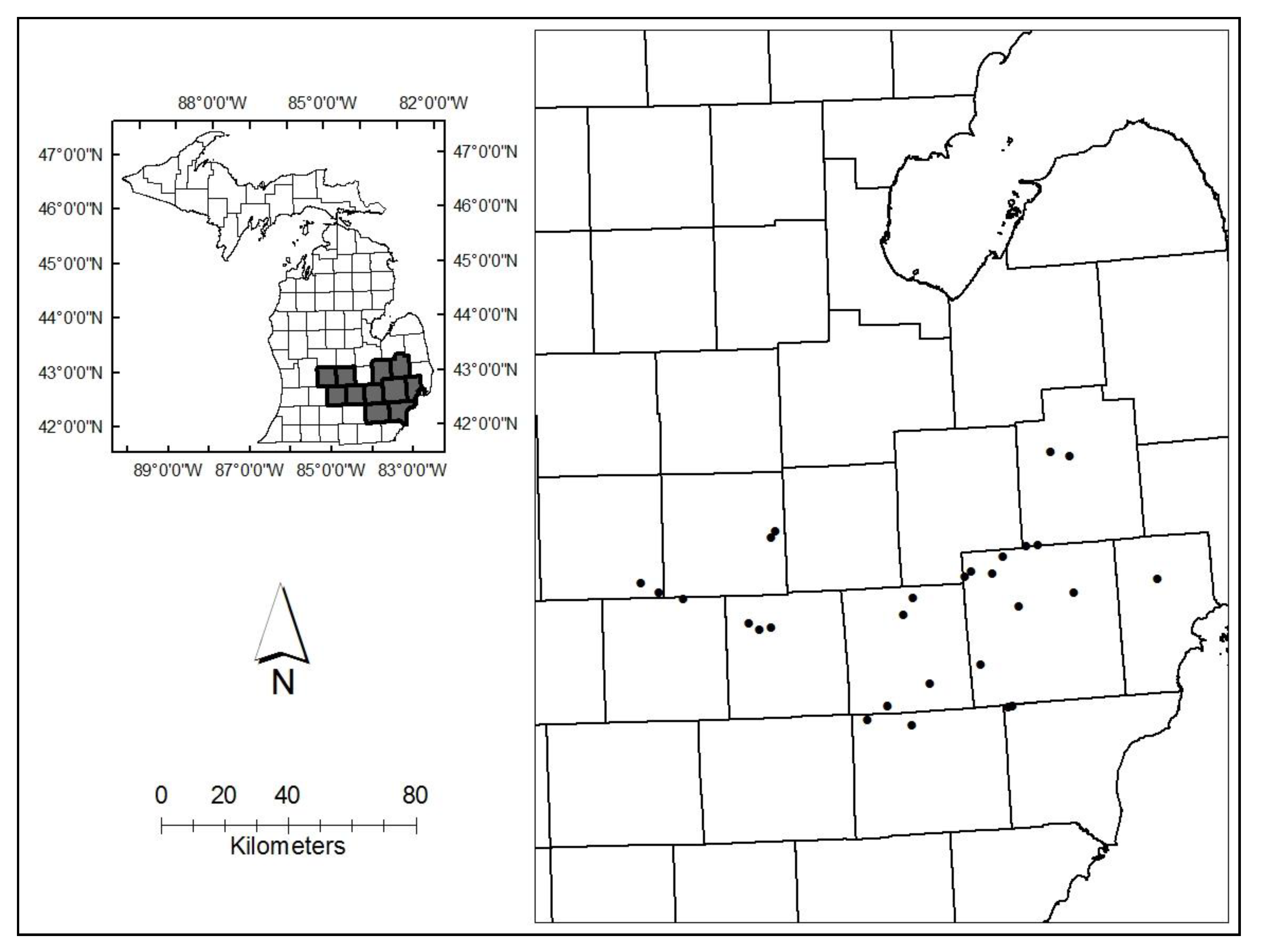
2.1. Site Selection
2.2. Fixed Radius Plots
2.3. Trapping
2.4. Statistical Analysis
3. Results
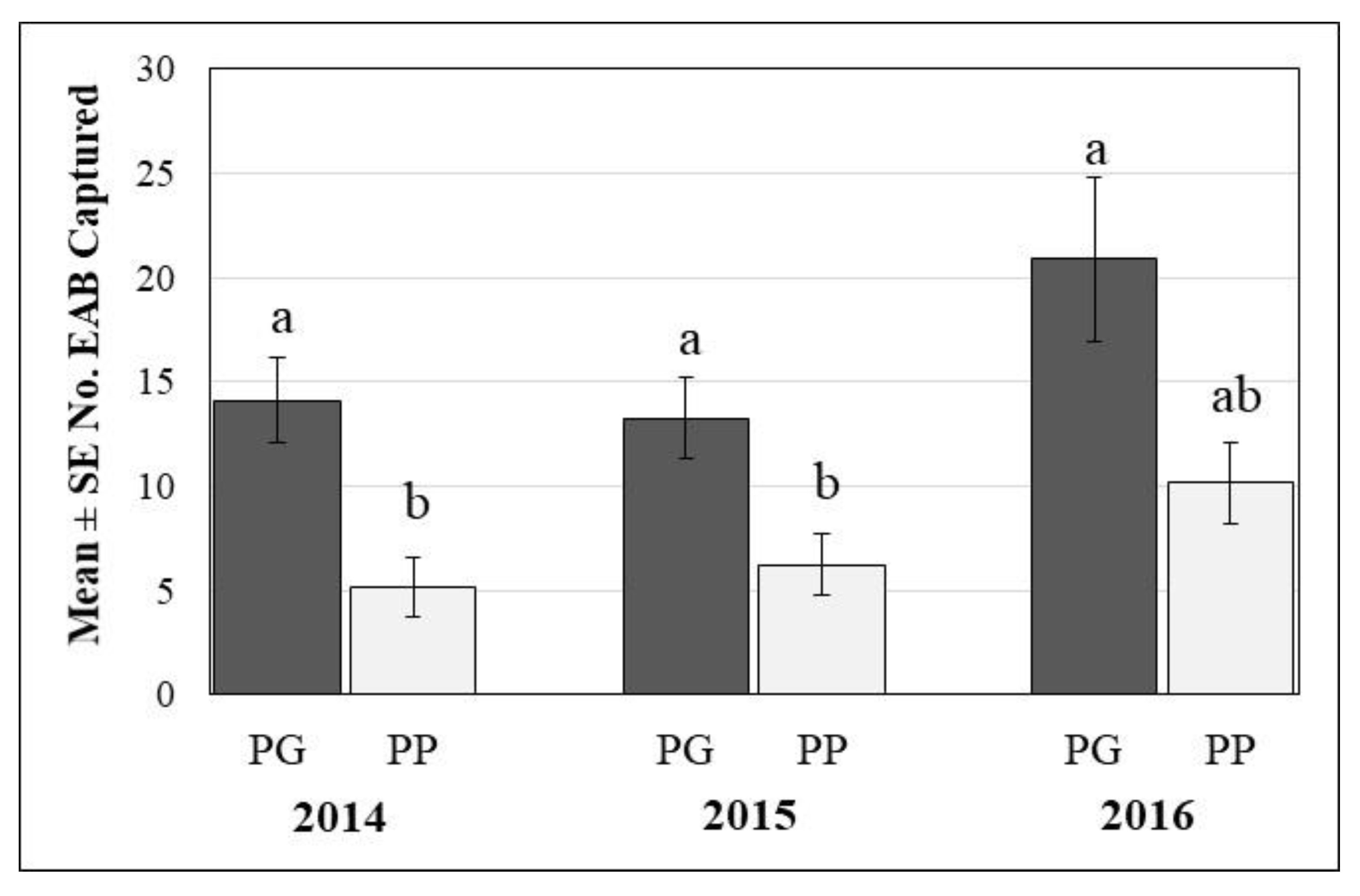
3.1. Captures of EAB on Traps
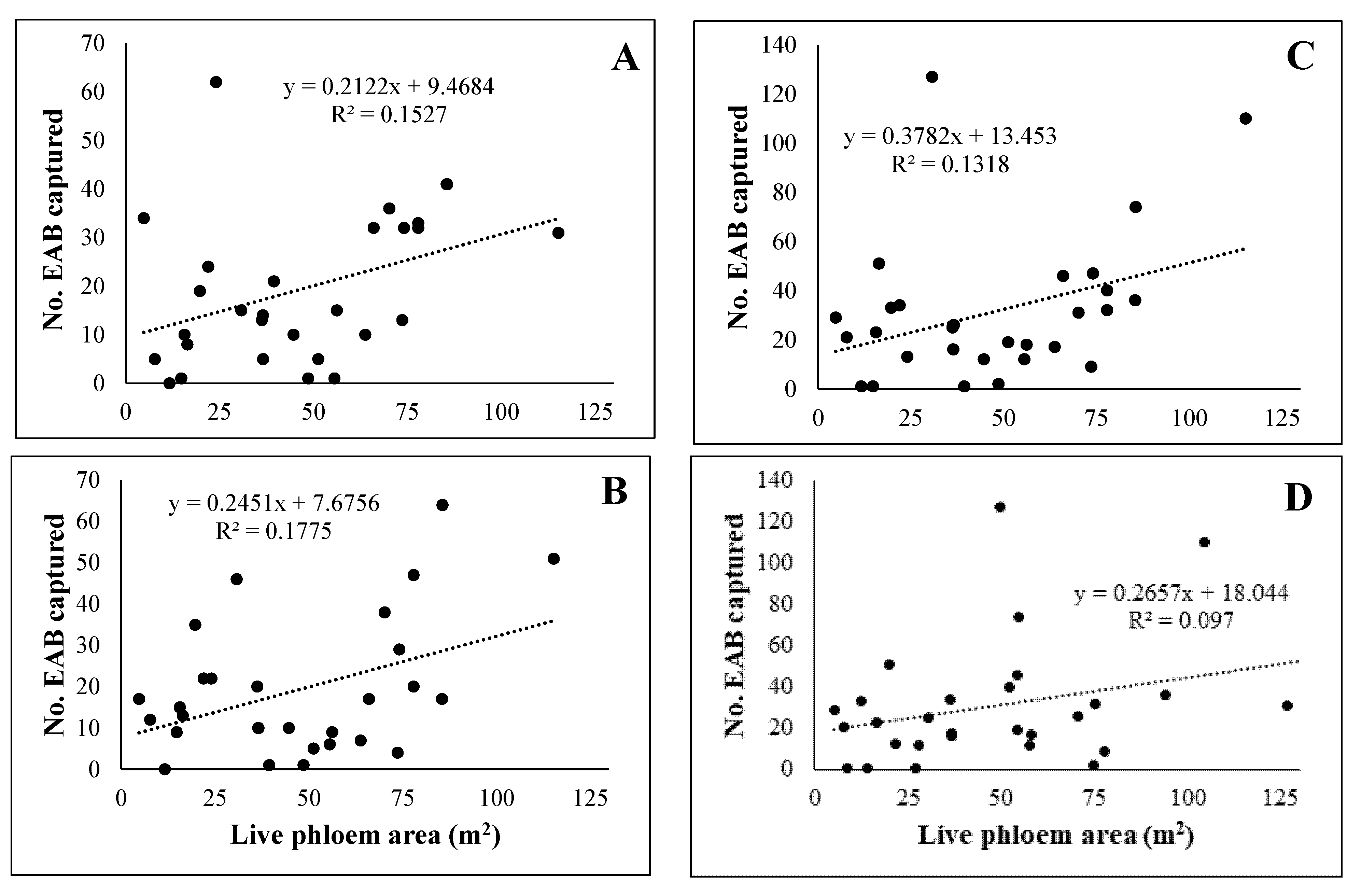
3.2. White Ash Phloem
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cappaert, D.L.; McCullough, D.G.; Poland, T.M.; Siegert, N.W. Emerald ash borer in North America: A research and regulatory challenge. Am. Entomol. 2005, 51, 152–165. [Google Scholar] [CrossRef] [Green Version]
- Poland, T.M.; McCullough, D.G. Emerald ash borer: Invasion of the urban forest and the threat to North America’s ash resource. J. For. 2006, 104, 118–124. [Google Scholar]
- Siegert, N.W.; McCullough, D.G.; Liebhold, A.M.; Telewski, F.W. Dendrochronological reconstruction of the epicentre and early spread of emerald ash borer in North America. Divers. Distrib. 2014, 20, 847–858. [Google Scholar] [CrossRef]
- Emerald Ash Borer Information Network (EAB.info). Available online: http://www.emeraldashborer.info/index.cfm (accessed on 30 October 2020).
- Aukema, J.E.; Leung, B.; Kovacs, K.; Chivers, C.; Britton, K.O.; Englin, J.; Frankel, S.J.; Haight, R.G.; Holmes, T.P.; Liebhold, A.; et al. Economic impacts of non-native forest insects in the continental United States. PLoS ONE 2011, 6, e24587. [Google Scholar] [CrossRef]
- Herms, D.A.; McCullough, D.G. Emerald ash borer invasion of North America: History, biology, ecology, impacts, and management. Ann. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCullough, D.G. Challenges, tactics and integrated management of emerald ash borer in North America. For. Int. J. For. Res. 2020, 93, 197–211. [Google Scholar] [CrossRef]
- Burr, S.J.; McCullough, D.G. Condition of green ash (Fraxinus pennsylvanica) overstory and regeneration at three stages of the emerald ash borer invasion wave. Can. J. For. Res. 2014, 44, 768–776. [Google Scholar] [CrossRef]
- Klooster, W.S.; Herms, D.A.; Knight, K.S.; Herms, C.P.; McCullough, D.G.; Smith, A.; Gandhi, K.J.K.; Cardina, J. Ash (Fraxinus spp.) mortality, regeneration, and seed bank dynamics in mixed hardwood forests following invasion by emerald ash borer (Agrilus planipennis). Biol. Invas. 2014, 16, 859–873. [Google Scholar] [CrossRef]
- Smith, A.; Herms, D.A.; Long, R.P.; Gandhi, K.J. Community composition and structure had no effect on forest susceptibility to invasion by the emerald ash borer (Coleoptera: Buprestidae). Can. Entomol. 2015, 147, 318–328. [Google Scholar] [CrossRef]
- Wang, X.Y.; Yang, Z.Q.; Gould, J.R.; Zhang, Y.N.; Liu, G.J.; Liu, E.S. The biology and ecology of the emerald ash borer Agrilus planipennis, in China. J. Insect Sci. 2010, 10, 128. [Google Scholar] [CrossRef] [Green Version]
- Anulewicz, A.C.; McCullough, D.G.; Cappaert, D.L. Emerald ash borer (Agrilus planipennis) density and canopy dieback in three North American ash species. Arboricult. Urban For. 2007, 33, 338–349. [Google Scholar]
- Tanis, S.R.; McCullough, D.G. Host resistance of five Fraxinus species to Agrilus planipennis (Coleoptera: Buprestidae) and effects of paclobutrazol and fertilization. Environ. Entomol. 2015, 44, 287–299. [Google Scholar] [CrossRef]
- Tanis, S.R.; McCullough, D.G. Differential persistence of blue ash and white ash following emerald ash borer invasion. Can. J. For. Res. 2012, 42, 1542–1550. [Google Scholar] [CrossRef]
- Robinett, M.A.; McCullough, D.G. White ash (Fraxinus americana) survival in the core of the emerald ash borer (Agrilus planipennis) invasion. Can. J. For. Res. 2019, 49, 510–520. [Google Scholar] [CrossRef]
- Mercader, R.J.; Siegert, N.W.; Liebhold, A.M.; McCullough, D.G. Estimating the effectiveness of three potential management options to slow the spread of emerald ash borer populations in localized outlier sites. Can. J. For. Res. 2011, 41, 254–264. [Google Scholar] [CrossRef]
- Mercader, R.J.; Siegert, N.W.; Liebhold, A.M.; McCullough, D.G. Simulating the influence of the spatial distribution of host trees on the spread of the emerald ash borer, Agrilus planipennis, in recently colonized sites. Popul. Biol. 2011, 53, 271–285. [Google Scholar]
- McCullough, D.G.; Siegert, N.W. Estimating potential emerald ash borer (Coleoptera: Buprestidae) populations using ash inventory data. J. Econ. Entomol. 2007, 100, 1577–1586. [Google Scholar] [CrossRef]
- Burr, S.J.; McCullough, D.G.; Poland, T.M. Density of emerald ash borer (Coleoptera: Buprestidae) adults and larvae at three stages of the invasion wave. Environ. Entomol. 2018, 47, 121–132. [Google Scholar] [CrossRef]
- McCullough, D.G.; Poland, T.M.; Tluczek, A.R.; Anulewicz, A.C.; Wieferich, J.; Siegert, N.W. Emerald ash borer (Coleoptera: Buprestidae) densities over a 6-yr period on untreated trees and trees treated with systemic insecticides at 1-, 2-, and 3-yr intervals in a central Michigan forest. J. Econ. Entomol. 2019, 112, 201–212. [Google Scholar] [CrossRef]
- Siegert, N.W.; Engelken, P.; McCullough, D.G. Changes in demography and carrying capacity of green ash and black ash ten years after emerald ash borer invasion of two ash-dominant forests. For. Ecol. Manage. 2021. submitted. [Google Scholar]
- Engelken, P.; McCullough, D.G. Riparian forest conditions along three northern Michigan rivers following emerald ash borer invasion. Can. J. For. Res. 2020, 50, 800–810. [Google Scholar] [CrossRef]
- Engelken, P.; Benbow, M.E.; McCullough, D.G. Legacy effects of emerald ash borer on riparian forest vegetation and structure. For. Ecol. Manage. 2020, 457, 117684. [Google Scholar] [CrossRef]
- Kashian, D.M. Sprouting and seed production may promote persistence of green ash in the presence of the emerald ash borer. Ecosphere 2016, 7, e01332. [Google Scholar] [CrossRef] [Green Version]
- Aubin, I.; Cardou, F.; Ryall, K.; Kreutzweiser, D.; Scarr, T. Ash regeneration capacity after emerald ash borer (EAB) outbreaks: Some early results. For. Chron. 2015, 91, 291–298. [Google Scholar] [CrossRef]
- Siegert, N.W.; McCullough, D.G.; Poland, T.M.; Heyd, R.L. Optimizing use of girdled ash trees for management of low-density emerald ash borer populations. J. Econ. Entomol. 2017, 110, 1096–1106. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.J.; Abell, K.J.; Bauer, L.S.; Gould, J.; Van Driesche, R.G. Natural enemies implicated in the regulation of an invasive pest: A life table analysis of the population dynamics of the emerald ash borer. Agric. For. Entomol. 2014, 16, 406–416. [Google Scholar] [CrossRef]
- Duan, J.J.; Bauer, L.S.; Van Driesche, R.G. Emerald ash borer biocontrol in ash saplings: The potential for early stage recovery of North American ash trees. For. Ecol. Manag. 2017, 394, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Crook, D.J.; Mastro, V.C. Chemical ecology of the emerald ash borer Agrilus planipennis. J. Chem. Ecol. 2010, 36, 101–112. [Google Scholar] [CrossRef]
- Ryall, K. Detection and sampling of emerald ash borer (Coleoptera: Buprestidae) infestations. Can. Entomol. 2015, 147, 290–299. [Google Scholar] [CrossRef]
- Poland, T.M.; Petrice, T.R.; Ciaramitaro, T.M. Trap designs, colors, and lures for emerald ash borer detection. Front. For. Glob. Chang. 2019, 2, 80. [Google Scholar] [CrossRef] [Green Version]
- Poland, T.M.; McCullough, D.G.; Anulewicz, A.C. Evaluation of double-decker traps for emerald ash borer (Coleoptera: Buprestidae). J. Econ. Entomol. 2011, 104, 517–531. [Google Scholar] [CrossRef]
- Silk, P.J.; Mayo, P.; Ryall, K.; Roscoe, L. Semiochemical and communication ecology of the emerald ash borer, Agrilus planipennis (Coleoptera: Buprestidae). Insects 2019, 10, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCullough, D.G.; Poland, T.M. Building double-decker traps for early detection of emerald ash borer. J. Vis. Exp. 2017, 128, e55252. [Google Scholar] [CrossRef] [PubMed]
- McCullough, D.G.; Siegert, N.W.; Poland, T.M.; Pierce, S.J.; Ahn, S.Z. Effects of trap type, placement and ash distribution on emerald ash borer captures in a low density site. Environ. Entomol. 2011, 40, 1239–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poland, T.M.; McCullough, D.G. Comparison of trap types and colors for capturing emerald ash borer adults at different population densities. Environ. Entomol. 2014, 43, 157–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarnoch, S.J.; Bechtold, W.A.; Stolte, K.W. Using crown condition variables as indicators of forest health. Can. J. For. Res. 2004, 34, 1057–1070. [Google Scholar] [CrossRef]
- Limback, C.K. Tree Vigor and Its Relation to Emerald Ash Borer (Agrilus planipennis Fairmarie) Adult Host Preference and Larval Development on Green and White Ash Trees. Master’s Thesis, Department of Entomology, Michigan State University, East Lansing, MI, USA, 2010; p. 98. [Google Scholar]
- Flower, C.E.; Knight, K.S.; Rebbeck, J.; Gonzalez-Meler, M.A. The relationship between the emerald ash borer (Agrilus planipennis) and ash (Fraxinus spp.) tree decline: Using visual canopy condition assessments and leaf isotope measurements to assess pest damage. For. Ecol. Manag. 2013, 303, 143–147. [Google Scholar] [CrossRef]
- Tobin, P.C.; Strom, B.L.; Francese, J.A.; Herms, D.A.; McCullough, D.G.; Poland, T.M.; Ryall, K.L.; Scarr, T.S.; Silk, P.J.; Thistle, H.W. Evaluation of trapping schemes to detect emerald ash borer. Environ. Entomol. 2021. Accepted. [Google Scholar]
- Francese, J.; Rietz, M.L.; Crook, D.J.; Fraser, I.; Lance, D.R.; Mastro, V.C. Improving detection tools for the emerald ash borer (Coleoptera: Buprestidae): Comparison of prism and multifunnel traps at varying population densities. J. Econ. Entomol. 2013, 106, 2407–2414. [Google Scholar] [CrossRef]
- Alnosaier, W.S. Kenward-Roger Approximate F Test for Fixed Effects in Mixed Linear Models. Ph.D. Dissertation, Department of Statistics, Oregon State University, Corvallis, OR, USA, 2007; p. 131. [Google Scholar]
- Hilbe, J.M. Negative Binomial Regression, 2nd ed.; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- SAS Institute Inc. Base SAS 9.4 Procedures Guide: Statistical Procedures, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2012. [Google Scholar]
- Mercader, R.J.; McCullough, D.G.; Bedford, J.M. A comparison of girdled ash detection trees and baited artificial traps for emerald ash borer (Agrilus planipennis Fairmaire) detection. Environ. Entomol. 2013, 42, 1027–1039. [Google Scholar] [CrossRef] [Green Version]
- Mercader, R.J.; Siegert, N.W.; Liebhold, A.M.; McCullough, D.G. Dispersal of the emerald ash borer, Agrilus planipennis, in newly-colonized sites. Agric. For. Entomol. 2009, 11, 421–424. [Google Scholar] [CrossRef]
- Mercader, R.J.; Siegert, N.W.; McCullough, D.G. Estimating the influence of population density and dispersal behavior on the ability to detect and monitor Agrilus planipennis (Coleoptera: Buprestidae) populations. J. Econ. Entomol. 2012, 105, 272–281. [Google Scholar] [CrossRef]
- Siegert, N.W.; McCullough, D.G.; Williams, D.W.; Fraser, I.; Poland, T.M. Dispersal of Agrilus planipennis (Coleoptera: Buprestidae) from discrete epicenters in two outlier sites. Environ. Entomol. 2010, 39, 253–265. [Google Scholar] [CrossRef]
- Taylor, R.A.J.; Bauer, L.S.; Poland, T.M.; Windell, K.N. Flight performance of Agrilus planipennis (Coleoptera: Buprestidae) on a flight mill and in free flight. J. Insect Behav. 2010, 23, 128–148. [Google Scholar] [CrossRef]
- Siegert, N.W.; Mercader, R.J.; McCullough, D.G. Spread and dispersal of emerald ash borer (Coleoptera: Buprestidae): Estimating the spatial dynamics of a difficult-to-detect invasive forest pest. Can. Entomol. 2015, 147, 338–348. [Google Scholar] [CrossRef]
- Tluczek, A.R.; McCullough, D.G.; Poland, T.M. Influence of host stress on emerald ash borer (Coleoptera: Buprestidae) adult density, development, and distribution in Fraxinus pennsylvanica trees. Environ. Entomol. 2011, 40, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Crook, D.J.; Khrimian, A.; Francese, J.A.; Fraser, I.; Poland, T.M.; Sawyer, A.J.; Mastro, V.C. Development of a host-based semiochemical lure for trapping emerald ash borer Agrilus planipennis (Coleoptera: Buprestidae). Environ. Entomol. 2008, 37, 356–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crook, D.J.; Francese, J.A.; Zylstra, K.E.; Fraser, I.; Sawyer, A.J.; Bartels, D.W.; Lance, D.R.; Mastro, V.C. Laboratory and field response of the emerald ash borer (Coleoptera: Buprestidae), to selected regions of the electromagnetic spectrum. J. Econ. Entomol. 2009, 102, 2160–2169. [Google Scholar] [CrossRef]
- Anulewicz, A.C.; McCullough, D.G.; Cappaert, D.L.; Poland, T.M. Host range of the emerald ash borer (Agrilus planipennis Fairmaire) (Coleoptera:Buprestidae) in North America: Results of multiple-choice field experiments. Environ. Entomol. 2008, 37, 230–241. [Google Scholar] [CrossRef] [Green Version]
- Cappaert, D.; McCullough, D.G. Phenology of Atanycolus cappaerti (Hymenoptera: Braconidae), a native parasitoid of emerald ash borer, Agrilus planipennis (Coleoptera: Buprestidae). Great Lakes Entomol. 2009, 41, 141–154. [Google Scholar]
- Lindell, C.A.; McCullough, D.G.; Cappaert, D.; Apostolou, N.M.; Roth, M.B. Factors influencing woodpecker predation on emerald ash borer. Am. Midl. Nat. 2008, 159, 434–444. [Google Scholar] [CrossRef]
- Duan, J.J.; Ulyshen, M.D.; Bauer, L.S.; Gould, J.; Van Driesche, R. Measuring the impact of biotic factors on populations of immature emerald ash borers (Coleoptera: Buprestidae). Environ. Entomol. 2010, 39, 1513–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.J.; Bauer, L.S.; Abell, K.J.; Van Driesche, R. Population responses of hymenopteran parasitoids to the emerald ash borer (Coleoptera: Buprestidae) in recently invaded areas in north central United States. BioControl 2012, 57, 199–209. [Google Scholar] [CrossRef]
- Duan, J.J.; Bauer, L.S.; Van Driesche, R.G.; Gould, J.R. Progress and challenges of protecting North American ash trees from the emerald ash borer using biological control. Forests 2018, 9, 142. [Google Scholar] [CrossRef] [Green Version]
- Kashian, D.M.; Bauer, L.S.; Spei, B.A.; Duan, J.J.; Gould, J.R. Potential impacts of emerald ash borer biocontrol on ash health and recovery in southern Michigan. Forests 2018, 9, 296. [Google Scholar] [CrossRef] [Green Version]
- Crook, D.J.; Khrimian, A.; Cossé, A.A.; Fraser, I.; Mastro, V.C. Influence of trap color and host volatiles on capture of the emerald ash borer (Coleoptera: Buprestidae). J. Econ. Entomol. 2012, 105, 429–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francese, J.A.; Crook, D.J.; Fraser, I.; Lance, D.R.; Sawyer, A.J.; Mastro, V.C. Optimization of trap color for emerald ash borer (Coleoptera: Buprestidae). J. Econ. Entomol. 2010, 103, 1235–1241. [Google Scholar] [CrossRef]
- Grant, G.G.; Poland, T.M.; Ciaramitaro, T.; Lyons, D.B.; Jones, G.C. Comparison of male and female emerald ash borer (Coleoptera: Buprestidae) responses to phoebe oil and (Z)-3-hexenol lures in light green prism traps. J. Econ. Entomol. 2011, 104, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Ryall, K.L.; Silk, P.J.; Mayo, P.; Crook, D.; Khrimian, A.; Cossé, A.A.; Sweeney, J.; Scarr, T.A. Attraction of Agrilus planipennis (Coleoptera: Buprestidae) to a volatile pheromone: Effects of release rate, host volatile, and trap placement. Environ. Entomol. 2012, 41, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Lelito, J.P.; Fraser, I.; Mastro, V.C.; Tumlinson, J.H.; Böröczky, K.; Baker, T.C. Visually mediated ‘paratrooper copulations’ in the mating behavior of Agrilus planipennis (Coleoptera: Buprestidae), a highly destructive invasive pest of North American ash trees. Insect Behav. 2007, 20, 537–552. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Saona, C.R.; Miller, J.R.; Poland, T.M.; Kuhn, T.M.; Otis, G.W.; Turk, T.; Ward, D.L. Behaviors of adult Agrilus planipennis (Coleoptera: Buprestidae). Great Lakes Entomol. 2007, 40, 1–16. [Google Scholar]
- Marshall, J.M.; Storer, A.J.; Fraser, I.; Mastro, V.C. Multi-state comparison of detection tools at low emerald ash borer densities. In Proceedings of the Emerald Ash Borer and Asian Longhorned Beetle Research and Technology Development Meeting, Pittsburgh, PA, USA, 19–21 October 2009; Lance, D., Buck, J., Binion, D., Reardon, R., Mastro, V.U.S., Eds.; Forest Service Publication FHTET 2010-01; United States Department of Agriculture: Morgantown, WV, USA, 2010; pp. 124–125. [Google Scholar]
- McCullough, D.G.; Poland, T.M.; Cappaert, D.; Anulewicz, A.C. Attraction of the emerald ash borer to ash trees stressed by girdling, herbicide and wounding. Can. J. For. Res. 2009, 39, 1331–1345. [Google Scholar] [CrossRef]
- McCullough, D.G.; Poland, T.M.; Anulewicz, A.C.; Cappaert, D. Emerald ash borer (Coleoptera: Buprestidae) attraction to stressed or baited ash trees. Environ. Entomol. 2009, 38, 1668–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, K.S.; Brown, J.P.; Long, R.P. Factors affecting the survival of ash (Fraxinus spp.) trees infested by emerald ash borer (Agrilus planipennis). Biol. Invas. 2013, 15, 371–383. [Google Scholar] [CrossRef]
- Koch, J.L.; Carey, D.W.; Mason, M.E.; Poland, T.M.; Knight, K.S. Intraspecific variation in Fraxinus pennsylvanica responses to emerald ash borer (Agrilus planipennis). New For. 2015, 46, 995–1011. [Google Scholar] [CrossRef]
- Mercader, R.J.; McCullough, D.G.; Storer, A.J.; Bedford, J.M.; Heyd, R.; Poland, T.M.; Katovich, S. Evaluation of the potential use of a systemic insecticide and girdled trees in area wide management of the emerald ash borer. For. Ecol. Manag. 2015, 350, 70–80. [Google Scholar] [CrossRef]
- Wei, X.; Reardon, R.; Wu, Y.; Sun, J.H. Emerald ash borer, Agrilus planipennis Fairmaire (Coleoptera: Buprestidae), in China: A review and distribution survey. Acta Entomol. Sin. 2004, 47, 679–685. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Double Decker (GP) Trap | Double Decker (PP) Trap | F; df; p | ||||
|---|---|---|---|---|---|---|
| Green Upper | Light Purple Lower | Dark Purple Upper | Dark Purple Lower | |||
| 2014 | Males | 7.1 ± 1.2a 1 | 4.5 ± 0.9ab | 2.2 ± 0.5b | 1.6 ± 0.6b | 6.21; 3.116; 0.0006 |
| Females | 0.7 ± 0.2ab | 1.9 ± 0.4a | 0.6 ± 0.2b | 0.7 ± 0.3b | 8.73; 3.116; <0.0001 | |
| Total | 7.8 ± 1.3a | 6.4 ± 1.1a | 2.9 ± 0.7b | 2.3 ± 0.9b | 8.76; 3.116; <0.0001 | |
| 2015 | Males | 5.5 ± 1.1a | 4.1 ± 0.8a | 2.3 ± 0.6ab | 1.3 ± 0.4b | 4.72; 3.116; 0.004 |
| Females | 1.4 ± 0.3ab | 2.3 ±o.4a | 1.4 ±0.3ab | 1.1 ± 0.3b | 2.56; 3.116; 0.05 | |
| Total | 6.9 ± 1.3a | 6.4 ± 1.1ab | 3.8 ± 0.9ab | 2.4 ± 0.6b | 4.40; 3.116; 0.006 | |
| 2016 | Males | 8.7 ± 2.0a | 7.9 ± 1.6a | 4.6 ± 0.9ab | 2.8 ± 0.8b | 4.11; 3.116; 0.008 |
| Females | 1.0 ± 0.2b | 3.3 ±0.7a | 3.9 ± 0.8ab | 1.0 ± 0.3b | 5.81; 3.116; 0.001 | |
| Total | 9.8 ± 2.1a | 11.2 ± 2.2a | 6.3 ± 1.2ab | 3.9 ± 0.9b | 4.49; 3.116; 0.005 | |
| All years | Males | 7.1 ± 0.8a | 5.5 ± 0.7ab | 3.1 ± 0.4b | 1.9 ± 0.3b | 14.4; 3.348; <0.0001 |
| Females | 1.0 ± 0.1b | 2.5 ± 0.3a | 1.2 ± 0.2b | 0.9 ± 0.3b | 13.3; 3.348; <0.0001 | |
| Total | 8.1 ± 0.9a | 7.8 ± 0.9a | 4.3 ± 0.6b | 2.9 ± 0.5b | 16.3; 3.348; <0.0001 | |
| 2014 | |||
| Percent Canopy Dieback Class | No. Trees | Total Phloem (m2) | Adjusted Live Phloem (m2) |
| <10.0% | 353 | 829 | 829 |
| 10.0 to 19.9% | 124 | 248 | 233 |
| 20.0 to 39.9% | 131 | 307 | 232 |
| 40.0 to 59.9% | 49 | 151 | 80 |
| 60.0 to 79.9% | 35 | 106 | 37 |
| 80.0 to 99.0% | 44 | 154 | 20 |
| Dead | 373 | 1833 | 0 |
| 2016 | |||
| <10.0% | 608 | 1300 | 1300 |
| 10.1 to 19.9% | 54 | 138 | 124 |
| 20.0 to 39.9% | 50 | 119 | 90 |
| 40.0 to 59.9% | 14 | 72 | 39 |
| 60.0 to 79.9% | 17 | 48 | 16 |
| 80.0 to 99.9% | 14 | 58 | 8 |
| Dead | 430 | 1837 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robinett, M.A.; Poland, T.M.; McCullough, D.G. Captures of Emerald Ash Borer (Agrilus planipennis) Adults in Post-Invasion White Ash Sites with Varying Amounts of Live Phloem. Forests 2021, 12, 262. https://doi.org/10.3390/f12030262
Robinett MA, Poland TM, McCullough DG. Captures of Emerald Ash Borer (Agrilus planipennis) Adults in Post-Invasion White Ash Sites with Varying Amounts of Live Phloem. Forests. 2021; 12(3):262. https://doi.org/10.3390/f12030262
Chicago/Turabian StyleRobinett, Molly A., Therese M. Poland, and Deborah G. McCullough. 2021. "Captures of Emerald Ash Borer (Agrilus planipennis) Adults in Post-Invasion White Ash Sites with Varying Amounts of Live Phloem" Forests 12, no. 3: 262. https://doi.org/10.3390/f12030262
APA StyleRobinett, M. A., Poland, T. M., & McCullough, D. G. (2021). Captures of Emerald Ash Borer (Agrilus planipennis) Adults in Post-Invasion White Ash Sites with Varying Amounts of Live Phloem. Forests, 12(3), 262. https://doi.org/10.3390/f12030262