
First Assessment of the Benthic Meiofauna Sensitivity to Low Human-Impacted Mangroves in French Guiana


 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
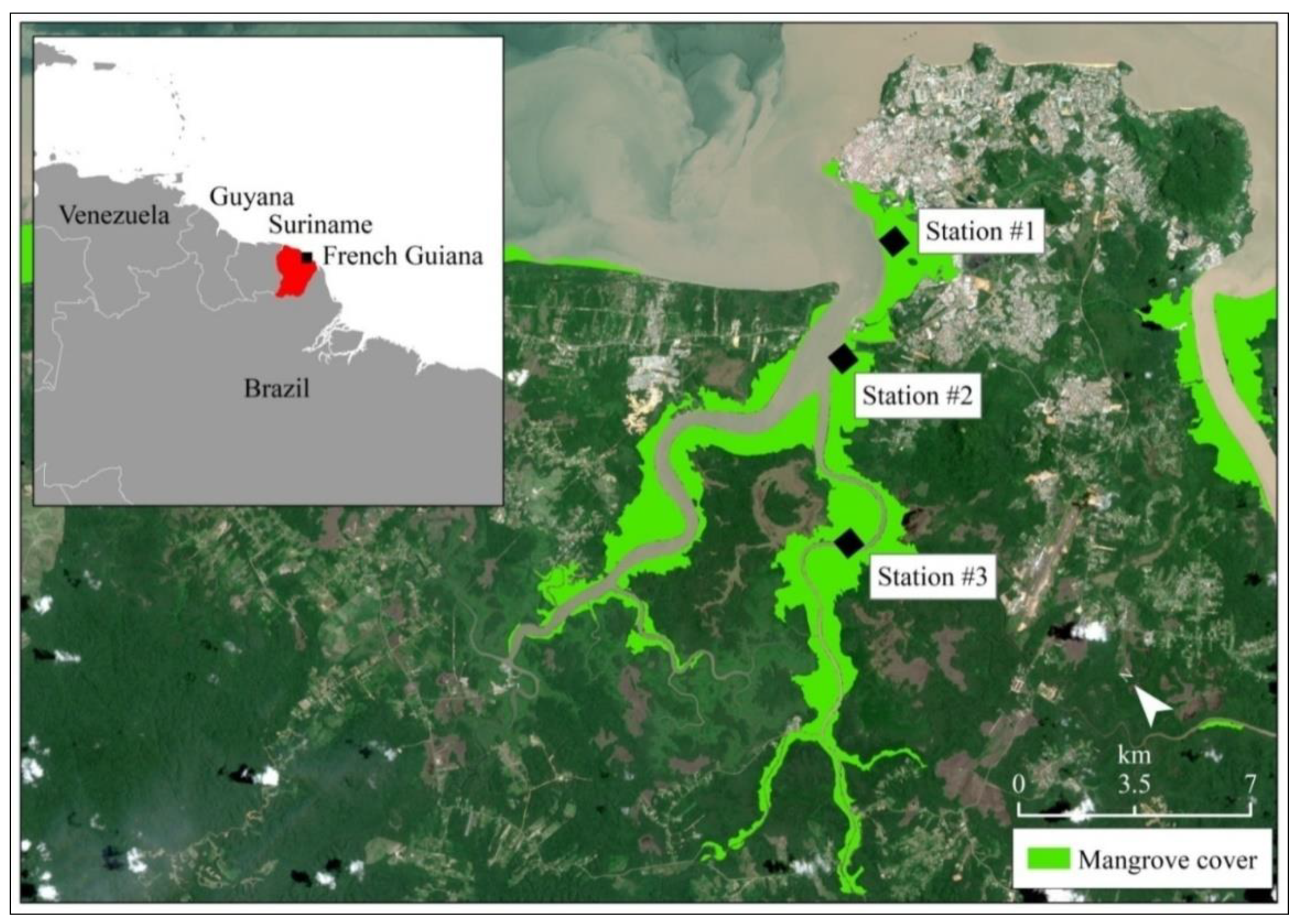
2.1. Study Area
2.2. Field Sampling
2.3. Laboratory Analyses
2.3.1. Grain Size, Carbon and Nitrogen Bulk Analyses
2.3.2. Pigment Analysis
2.3.3. Metals and Metalloids Analysis
2.3.4. Organic Contaminants Analysis
2.3.5. Bacterial and Archaeal Abundance
2.3.6. Meiofauna Analysis
2.4. Statistical Analyses
3. Results
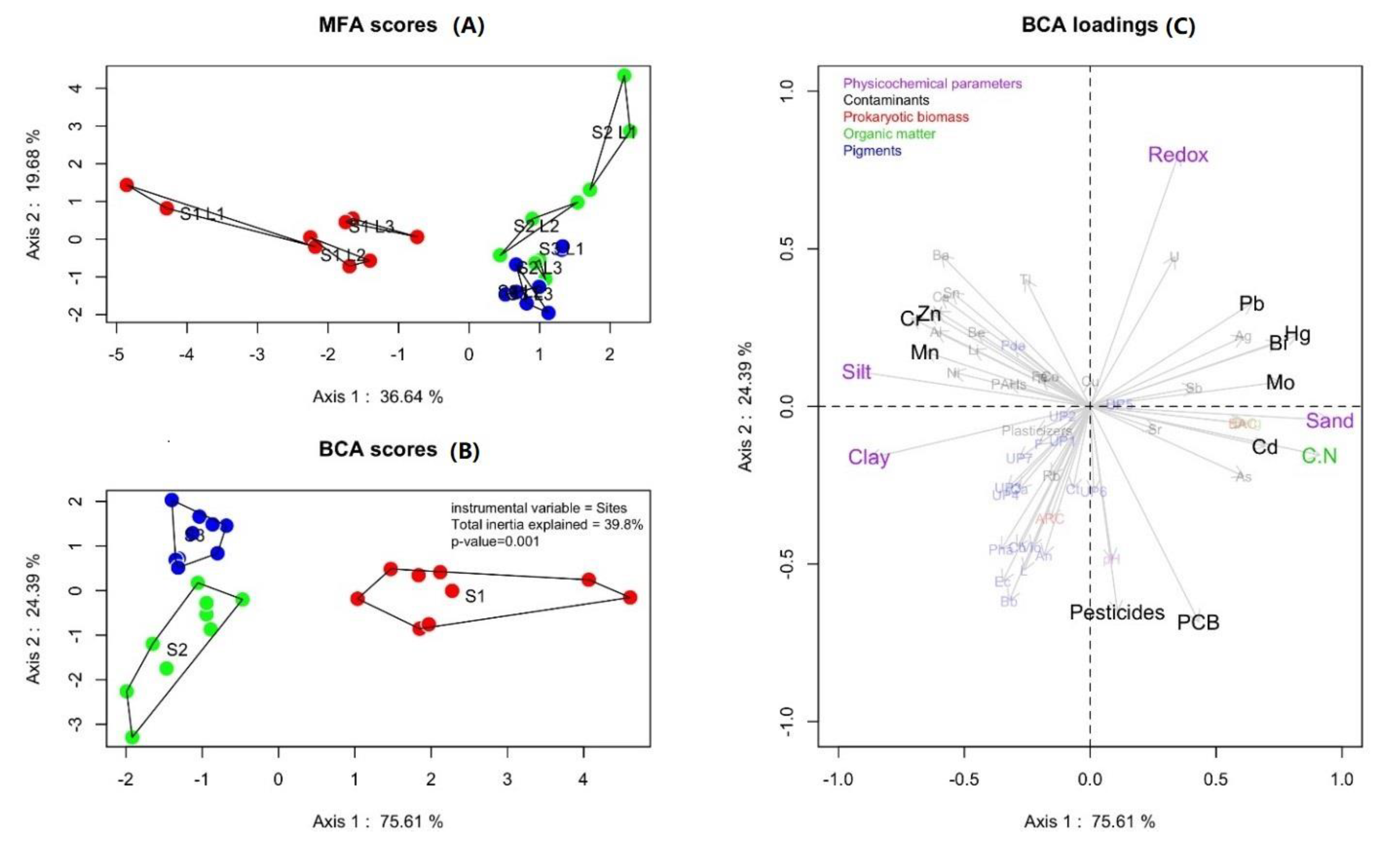
3.1. Environmental Characteristics
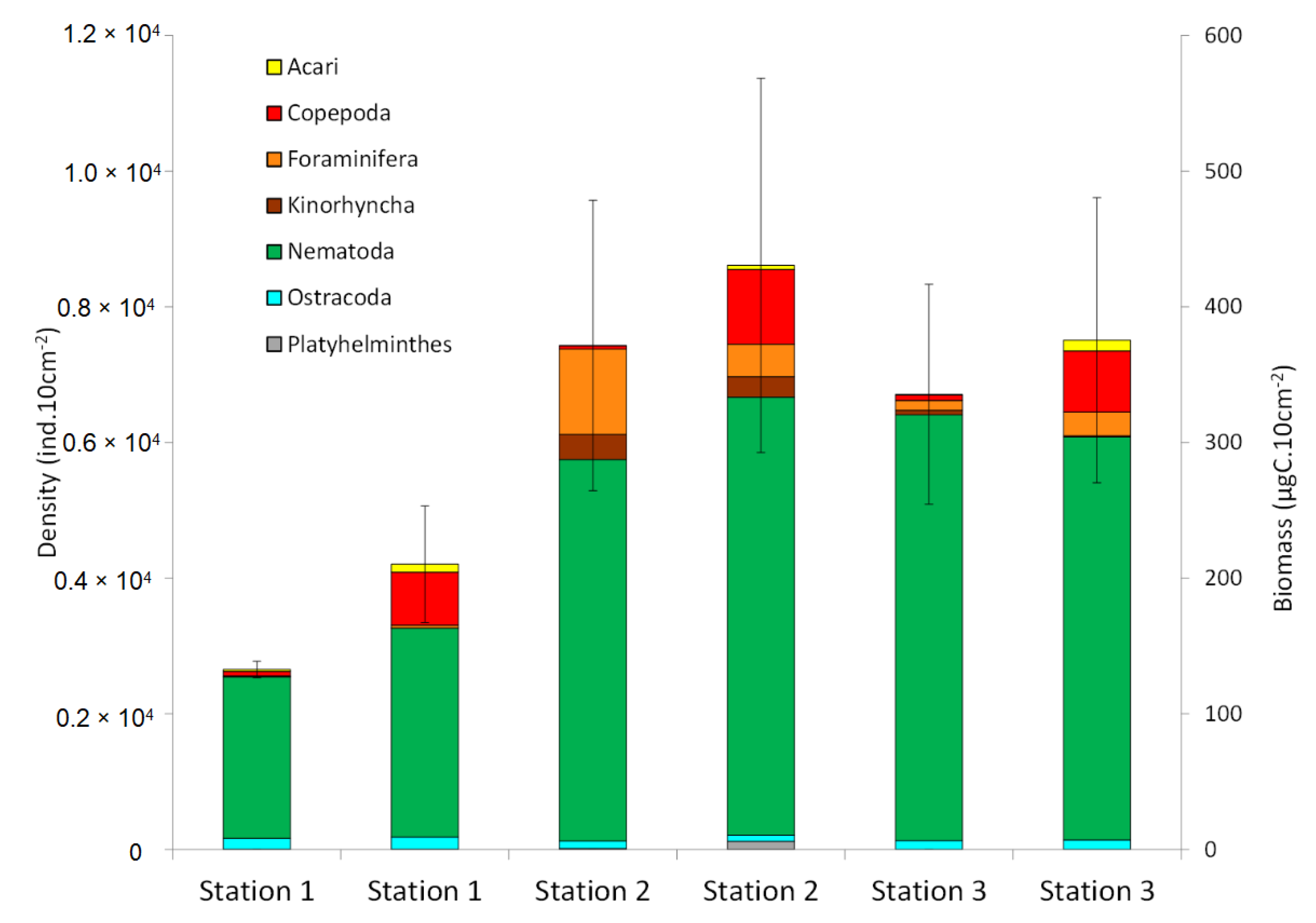
3.2. Total Meiofauna Community
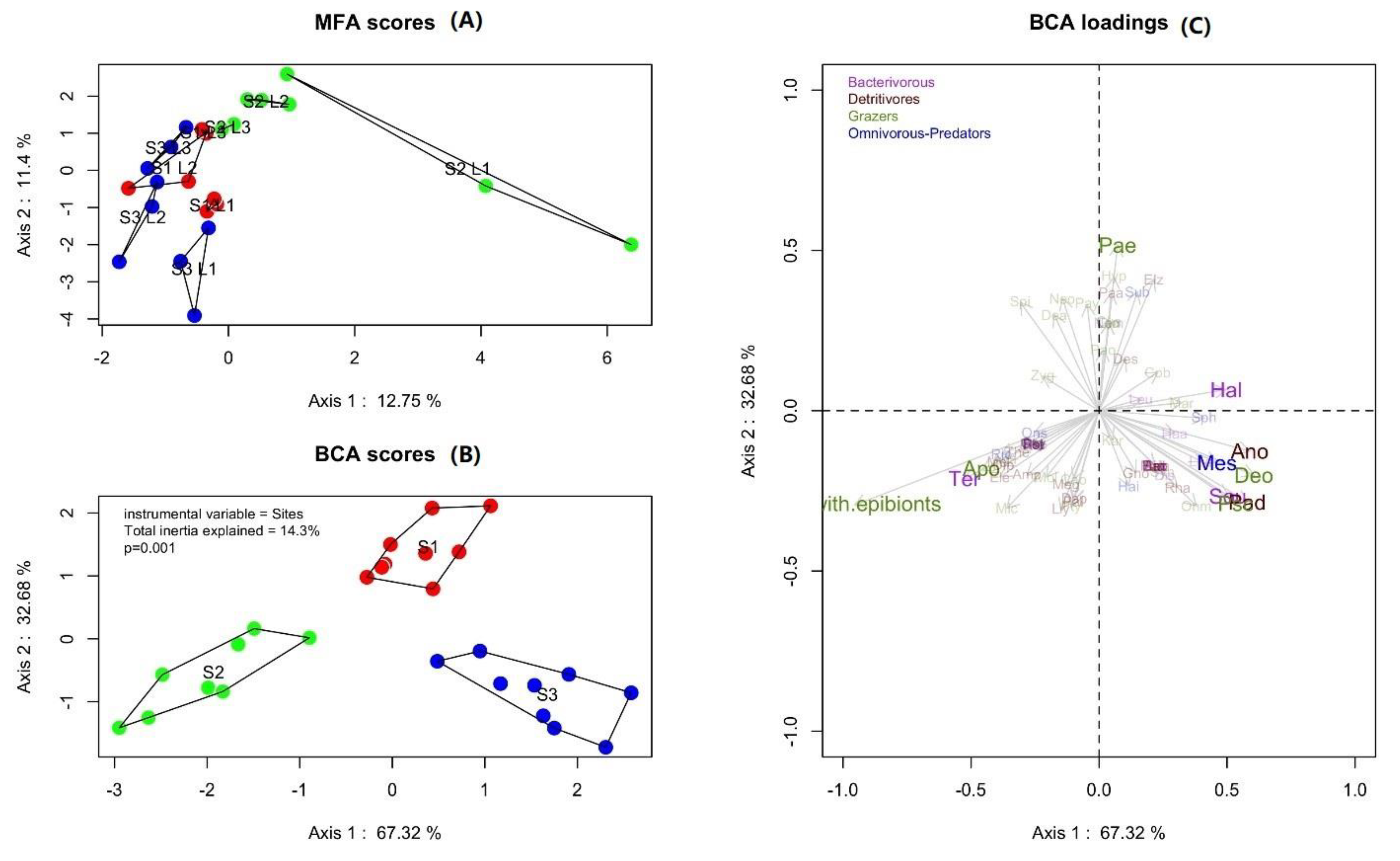
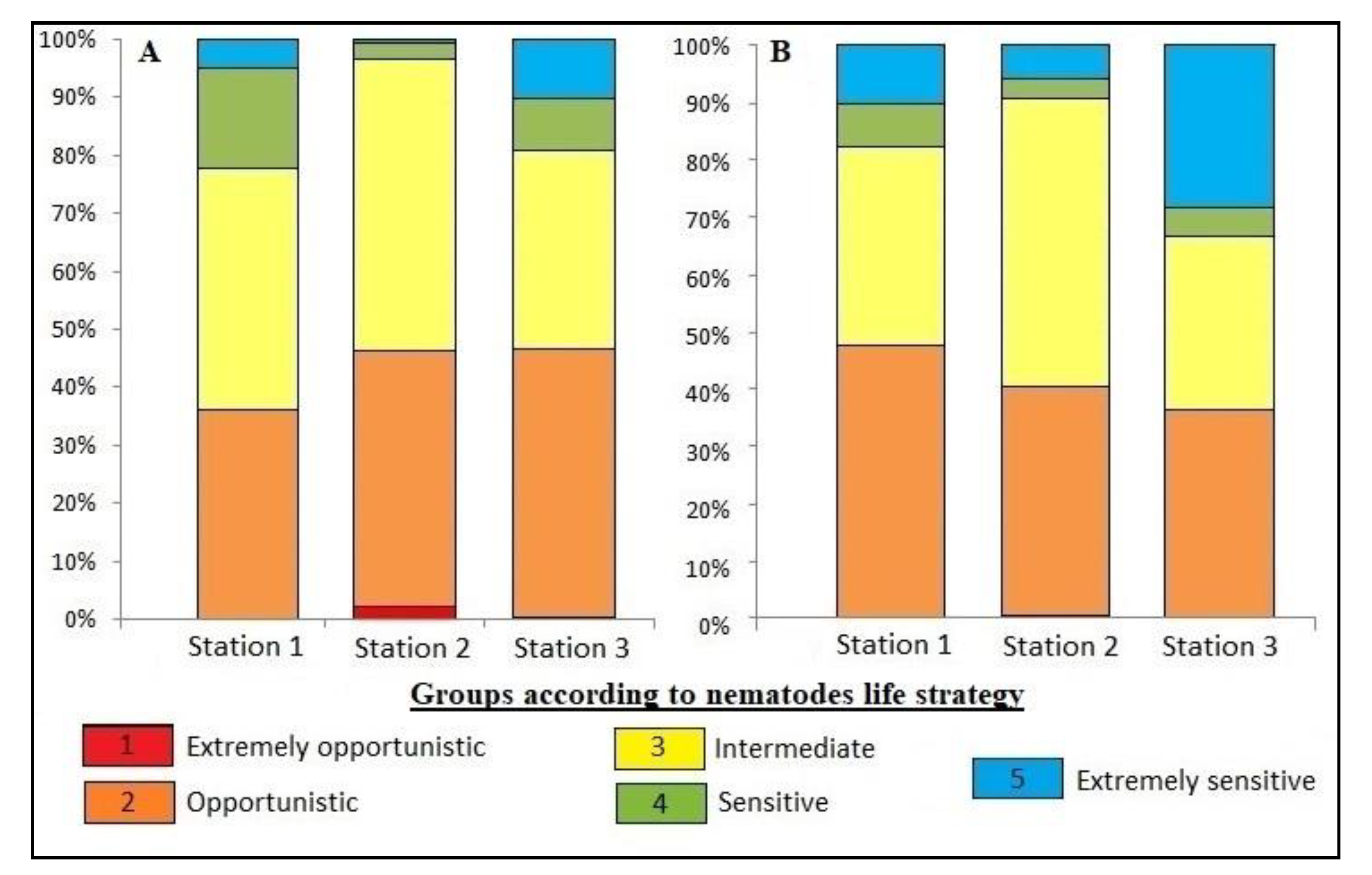
3.3. Nematoda Community
4. Discussion
4.1. Environmental Characteristics of the Three Stations
4.2. Overall Meiofauna Community
4.3. Nematodes Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valiela, I.; Bowen, J.L.; York, J.K. Mangrove Forests: One of the World’s Threatened Major Tropical Environments. BioScience 2001, 51, 807–815. [Google Scholar] [CrossRef] [Green Version]
- Worthington, T.; Spalding, M. Mangrove Restoration Potential: A Global Map Highlighting a Critical Opportunity; University of Cambridge: Cambridge, UK, 2018; pp. 1–34. [Google Scholar] [CrossRef]
- Worthington, T.A.; zu Ermgassen, P.S.E.; Friess, D.A.; Krauss, K.W.; Lovelock, C.E.; Thorley, J.; Tingey, R.; Woodroffe, C.D.; Bunting, P.; Cormier, N.; et al. A Global Biophysical Typology of Mangroves and Its Relevance for Ecosystem Structure and Deforestation. Sci. Rep. 2020, 10, 14652. [Google Scholar] [CrossRef]
- Adeel, Z.; Pomeroy, R. Assessment and Management of Mangrove Ecosystems in Developing Countries. Trees 2002, 16, 235–238. [Google Scholar] [CrossRef]
- Goldberg, L.; Lagomasino, D.; Thomas, N.; Fatoyinbo, T. Global Declines in Human-Driven Mangrove Loss. Glob. Change Biol. 2020, 26, 5844–5855. [Google Scholar] [CrossRef] [PubMed]
- Field, C.B.; Osborn, J.G.; Hoffman, L.L.; Polsenberg, J.F.; Ackerly, D.D.; Berry, J.A.; Bjorkman, O.; Held, A.; Matson, P.A.; Mooney, H.A. Mangrove Biodiversity and Ecosystem Function. Glob. Ecol. Biogeogr. Lett. 1998, 7, 3–14. [Google Scholar] [CrossRef]
- Lee, S.Y.; Primavera, J.H.; Dahdouh-Guebas, F.; McKee, K.; Bosire, J.O.; Cannicci, S.; Diele, K.; Fromard, F.; Koedam, N.; Marchand, C.; et al. Ecological Role and Services of Tropical Mangrove Ecosystems: A Reassessment. Glob. Ecol. Biogeogr. 2014, 23, 726–743. [Google Scholar] [CrossRef]
- Hai, N.T.; Dell, B.; Phuong, V.T.; Harper, R.J. Towards a More Robust Approach for the Restoration of Mangroves in Vietnam. Ann. For. Sci. 2020, 77, 18. [Google Scholar] [CrossRef] [Green Version]
- Kodikara, K.A.S.; Mukherjee, N.; Jayatissa, L.P.; Dahdouh-Guebas, F.; Koedam, N. Have Mangrove Restoration Projects Worked? An in-Depth Study in Sri Lanka. Restor. Ecol. 2017, 25, 705–716. [Google Scholar] [CrossRef]
- Richards, D.R.; Thompson, B.S.; Wijedasa, L. Quantifying Net Loss of Global Mangrove Carbon Stocks from 20 Years of Land Cover Change. Nat. Commun. 2020, 11, 4260. [Google Scholar] [CrossRef]
- Alongi, D.M. Impact of Global Change on Nutrient Dynamics in Mangrove Forests. Forests 2018, 9, 596. [Google Scholar] [CrossRef] [Green Version]
- McKee, K.L.; Cahoon, D.R.; Feller, I.C. Caribbean Mangroves Adjust to Rising Sea Level through Biotic Controls on Change in Soil Elevation. Glob. Ecol. Biogeogr. 2007, 16, 545–556. [Google Scholar] [CrossRef]
- Bosire, J.O.; Dahdouh-Guebas, F.; Walton, M.; Crona, B.I.; Lewis, R.R.; Field, C.; Kairo, J.G.; Koedam, N. Functionality of Restored Mangroves: A Review. Aquat. Bot. 2008, 89, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Thornton, S.R.; Johnstone, R.W. Mangrove Rehabilitation in High Erosion Areas: Assessment Using Bioindicators. Estuar. Coast. Shelf Sci. 2015, 165, 176–184. [Google Scholar] [CrossRef]
- Allard, S.M.; Costa, M.T.; Bulseco, A.N.; Helfer, V.; Wilkins, L.G.E.; Hassenrück, C.; Zengler, K.; Zimmer, M.; Erazo, N.; Rodrigues, J.L.M.; et al. Introducing the Mangrove Microbiome Initiative: Identifying Microbial Research Priorities and Approaches To Better Understand, Protect, and Rehabilitate Mangrove Ecosystems. mSystems 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Romañach, S.S.; DeAngelis, D.L.; Koh, H.L.; Li, Y.; Teh, S.Y.; Raja Barizan, R.S.; Zhai, L. Conservation and Restoration of Mangroves: Global Status, Perspectives, and Prognosis. Ocean Coast. Manag. 2018, 154, 72–82. [Google Scholar] [CrossRef]
- Cuny, P.; Jezequel, R.; Michaud, E.; Sylvi, L.; Gilbert, F.; Fiard, M.; Chevalier, C.; Morel, V.; Militon, C. Oil Spill Response in Mangroves: Why a Specific Ecosystem-Based Management Is Required? The Case of French Guiana – a Mini-Review. Vie et Milieu 2020, in press. [Google Scholar]
- Dirberg, G.; Barnaud, G.; Brivois, O.; Caessteker, P.; Corlier-Salem, M.; Cuny, P.; Fromard, F.; Fiard, M.; Gilbert, F.; Grouard, S.; et al. Towards the Development of Ecosystem-Based Indicators of Mangroves Functionin State in the Context of the EU Water Framework Directive. Vie et Milieu 2020, in press. [Google Scholar]
- Dauvin, J.-C.; Bellan, G.; Bellan-Santini, D. Benthic Indicators: From Subjectivity to Objectivity – Where Is the Line? Mar. Pollut. Bull. 2010, 60, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Sandulli, R.; De Nicola-Giudici, M. Pollution Effects on the Structure of Meiofaunal Communities in the Bay of Naples. Mar. Pollut. Bull. 1990, 21, 144–153. [Google Scholar] [CrossRef]
- Zeppilli, D.; Sarrazin, J.; Leduc, D.; Arbizu, P.M.; Fontaneto, D.; Fontanier, C.; Gooday, A.J.; Kristensen, R.M.; Ivanenko, V.N.; Sørensen, M.V.; et al. Is the Meiofauna a Good Indicator for Climate Change and Anthropogenic Impacts? Mar. Biodivers. 2015, 45, 505–535. [Google Scholar] [CrossRef] [Green Version]
- Alongi, D. Inter-Estuary Variation and Intertidal Zonation of Free-Living Nematode Communities in Tropical Mangrove Systems. Mar. Ecol. Prog. Ser. 1987, 40, 103–114. [Google Scholar] [CrossRef]
- Netto, S.A.; Gallucci, F. Meiofauna and Macrofauna Communities in a Mangrove from the Island of Santa Catarina, South Brazil. Hydrobiologia 2003, 505, 159–170. [Google Scholar] [CrossRef]
- Pinto, T.K.; Austen, M.C.V.; Warwick, R.M.; Somerfield, P.J.; Esteves, A.M.; Castro, F.J.V.; Fonseca-Genevois, V.G.; Santos, P.J.P. Nematode Diversity in Different Microhabitats in a Mangrove Region. Mar. Ecol. 2013, 34, 257–268. [Google Scholar] [CrossRef]
- Aschenbroich, A.; Michaud, E.; Gilbert, F.; Fromard, F.; Alt, A.; Le Garrec, V.; Bihannic, I.; De Coninck, A.; Thouzeau, G. Bioturbation Functional Roles Associated with Mangrove Development in French Guiana, South America. Hydrobiologia 2017, 794, 179–202. [Google Scholar] [CrossRef] [Green Version]
- Riera, P.; Hubas, C. Trophic Ecology of Nematodes from Various Microhabitats of the Roscoff Aber Bay (France): Importance of Stranded Macroalgae Evidenced through Δ13C and Δ15N. Mar. Ecol. Prog. Ser. 2003, 260, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Schratzberger, M.; Ingels, J. Meiofauna Matters: The Roles of Meiofauna in Benthic Ecosystems. J. Exp. Mar. Biol. Ecol. 2018, 502, 12–25. [Google Scholar] [CrossRef]
- Mutua, A.K.; Ntiba, M.J.; Muthumbi, A.; Ngondi, D.; Vanreusel, A. Restoration of Benthic Macro-Endofauna after Reforestation of Rhizophora Mucronata Mangroves in Gazi Bay, Kenya. West. Indian Ocean J Mar Sci 2011, 10, 39–49. [Google Scholar]
- Balsamo, M.; Semprucci, F.; Frontalini, F.; Coccioni, R. Meiofauna as a Tool for Marine Ecosystem Biomonitoring. Mar. Ecosyst. 2012. [Google Scholar] [CrossRef] [Green Version]
- Della Patrona, L.; Marchand, C.; Hubas, C.; Molnar, N.; Deborde, J.; Meziane, T. Meiofauna Distribution in a Mangrove Forest Exposed to Shrimp Farm Effluents (New Caledonia). Mar. Environ. Res. 2016, 119, 100–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, A.W. An Alternative Method of Cross-Validation for the Smoothing of Density Estimates. Biometrika 1984, 71, 353–360. [Google Scholar] [CrossRef]
- Gee, J.M.; Warwick, R.M.; Schaanning, M.; Berge, J.A.; Ambrose, W.G. Effects of Organic Enrichment on Meiofaunal Abundance and Community Structure in Sublittoral Soft Sediments. J. Exp. Mar. Biol. Ecol. 1985, 91, 247–262. [Google Scholar] [CrossRef]
- Haegerbaeumer, A.; Höss, S.; Ristau, K.; Claus, E.; Heininger, P.; Traunspurger, W. The Use of Meiofauna in Freshwater Sediment Assessments: Structural and Functional Responses of Meiobenthic Communities to Metal and Organics Contamination. Ecol. Indic. 2017, 78, 512–525. [Google Scholar] [CrossRef]
- Zeppilli, D.; Leduc, D.; Fontanier, C.; Fontaneto, D.; Fuchs, S.; Gooday, A.J.; Goineau, A.; Ingels, J.; Ivanenko, V.N.; Kristensen, R.M.; et al. Characteristics of Meiofauna in Extreme Marine Ecosystems: A Review. Mar. Biodivers. 2018, 48, 35–71. [Google Scholar] [CrossRef] [Green Version]
- Brustolin, M.C.; Nagelkerken, I.; Fonseca, G. Large-Scale Distribution Patterns of Mangrove Nematodes: A Global Meta-Analysis. Ecol. Evol. 2018, 8, 4734–4742. [Google Scholar] [CrossRef] [PubMed]
- Warwick, R.M.; Price, R. Ecological and Metabolic Studies on Free-Living Nematodes from an Estuarine Mud-Flat. Estuar. Coast. Mar. Sci. 1979, 9, 257–271. [Google Scholar] [CrossRef]
- Sahoo, G.; Suchiang, S.R.; Ansari, Z.A. Meiofauna-Mangrove Interaction: A Pilot Study from a Tropical Mangrove Habitat. Cah. Biol. Mar. 2013, 54, 349–358. [Google Scholar]
- Gyedu-Ababio, T.K.; Baird, D. Response of Meiofauna and Nematode Communities to Increased Levels of Contaminants in a Laboratory Microcosm Experiment. Ecotoxicol. Environ. Saf. 2006, 63, 443–450. [Google Scholar] [CrossRef]
- Austen, M.C.; Somerfield, P.J. A Community Level Sediment Bioassay Applied to an Estuarine Heavy Metal Gradient. Mar. Environ. Res. 1997, 43, 315–328. [Google Scholar] [CrossRef]
- Nasira, K.; Shahina, F.; Kamran, M. Response of Free-Living Marine Nematode Community to Heavy Metal Contaminantion along the Coastal Areas of Sindh and Balochistan Pakistan. Pak. J. Nematol. 2010, 28, 263–278. [Google Scholar]
- Monteiro, L.; Brinke, M.; dos Santos, G.; Traunspurger, W.; Moens, T. Effects of Heavy Metals on Free-Living Nematodes: A Multifaceted Approach Using Growth, Reproduction and Behavioural Assays. Eur. J. Soil Biol. 2014, 62, 1–7. [Google Scholar] [CrossRef]
- Bunting, P.; Rosenqvist, A.; Lucas, R.M.; Rebelo, L.M.; Hilarides, L.; Thomas, N.; Hardy, A.; Itoh, T.; Shimada, M.; Finlayson, C.M. The Global Mangrove Watch - A New 2010 Global Baseline of Mangrove Extent. Remote Sens. 2018, 10, 1669. [Google Scholar] [CrossRef] [Green Version]
- Anthony, E.J.; Gardel, A.; Gratiot, N.; Proisy, C.; Allison, M.A.; Dolique, F.; Fromard, F. The Amazon-Influenced Muddy Coast of South America: A Review of Mud-Bank–Shoreline Interactions. Earth-Sci. Rev. 2010, 103, 99–121. [Google Scholar] [CrossRef]
- Proisy, C.; Couteron, P.; Fromard, F. Predicting and Mapping Mangrove Biomass from Canopy Grain Analysis Using Fourier-Based Textural Ordination of IKONOS Images. Remote Sens. Environ. 2007, 109, 379–392. [Google Scholar] [CrossRef]
- Lescure, J.-P.; Tostain, O. Les mangroves guyanaises. Bois et Forêts. 1988, 220, 35–42. [Google Scholar]
- Lambs, L.; Muller, E.; Fromard, F. The Guianese Paradox: How Can the Freshwater Outflow from the Amazon Increase the Salinity of the Guianan Shore? J. Hydrol. 2007, 342, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Comte, J.; Moungin, B.; Renault, O. Inventaire Des Points de Rejet Sur La Zone Littorale; Report; BRGM: Cayenne, Guyane Française, 2000; p. 46, R 40917 SGR/GUY. [Google Scholar]
- Recensement de la population en Guyane, 1er Janvier 2018. INSEE. 2020. Available online: https://www.insee.fr/fr/statistiques/5005684 (accessed on 25 January 2021).
- Fromard, F.; Vega, C.; Proisy, C. Half a Century of Dynamic Coastal Change Affecting Mangrove Shorelines of French Guiana. A Case Study Based on Remote Sensing Data Analyses and Field Surveys. Mar. Geol. 2004, 208, 265–280. [Google Scholar] [CrossRef]
- Chaneac, L.; Legrand, C. Synthèse Bibliographique Sur Les Zones Humides de Guyane; Rapport Final; BRGM: Cayenne, Guyane Francaise, 2009; p. 137, BRGM/RP-57709-FR. [Google Scholar]
- Marchand, C.; Lallier-Vergès, E.; Baltzer, F. The Composition of Sedimentary Organic Matter in Relation to the Dynamic Features of a Mangrove-Fringed Coast in French Guiana. Estuar. Coast. Shelf Sci. 2003, 56, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Brotas, V.; Plante-Cuny, M.-R. The Use of HPLC Pigment Analysis to Study Microphytobenthos Communities. Acta Oecologica 2003, 24, S109–S115. [Google Scholar] [CrossRef]
- Munschy, C.; Tronczynski, J.; Heas-Moisan, K.; Guiot, N.; Truquet, I. Analyse de contaminants organiques (PCB, OCP, HAP) dans les organismes marins; Méthodes d’analyse en milieu marin; Ifremer/Ministère de l’écologie et du développement durable; Editions Quae: Versailles, France, 2005; ISBN 978-2-84433-144-1. [Google Scholar]
- Alzaga, R.; Montuori, P.; Ortiz, L.; Bayona, J.M.; Albaigés, J. Fast Solid-Phase Extraction–Gas Chromatography–Mass Spectrometry Procedure for Oil Fingerprinting: Application to the Prestige Oil Spill. J. Chromatogr. A 2004, 1025, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, C.; Le Cuff, N.; Receveur, J.; Moraga, D.; Auffret, M.; Guyomarch, J. Development of an Innovative and “Green” Stir Bar Sorptive Extraction–Thermal Desorption–Gas Chromatography–Tandem Mass Spectrometry Method for Quantification of Polycyclic Aromatic Hydrocarbons in Marine Biota. J. Chromatogr. A 2014, 1349, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jézéquel, R.; Duboscq, K.; Sylvi, L.; Michaud, E.; Ferriz, L.M.; Roic, E.; Duran, R.; Cravo-laureau, C.; Michotey, V.; Bonin, P.; et al. Assessment of Oil Weathering and Impact in Mangrove Ecosystem: PRISME Experiment. Int. Oil Spill Conf. Proc. 2017, 634–656. [Google Scholar] [CrossRef]
- Danovaro, R. Methods for the Study of Deep-Sea Sediments, Their Functioning and Biodiversity; CRC Press: Boca Raton, FL, USA, 2009; ISBN 978-0-429-13096-0. [Google Scholar]
- Warwick, R.; Platt, H.; Somerfield, P. Free-Living Marine Nematodes: Pictorial Key to World Genera and Notes for the Identification of British Species. Part 3: Monchysterids; Linnean Society of London and the Estuarine and Coastal Sciences Association: Shrewsbury, UK, 1998; p. 296. [Google Scholar]
- Wieser, W. Free-living marine nematodes. III. Axonolaimoidea and Monhysteroidea. Acta Universitatis Lund 1956, 52, 1–115. [Google Scholar]
- Moens, T.; Vincx, M. Observations on the Feeding Ecology of Estuarine Nematodes. J. Mar. Biol. Assoc. UK 1997, 77, 211–227. [Google Scholar] [CrossRef]
- Bongers, T. The Maturity Index: An Ecological Measure of Environmental Disturbance Based on Nematode Species Composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Escofier, B.; Pagès, J. Multiple Factor Analysis (AFMULT Package). Comput. Stat. Data Anal. 1994, 18, 121–140. [Google Scholar] [CrossRef]
- Dray, S.; Dufour, A.-B. The Ade4 Package: Implementing the Duality Diagram for Ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Abdi, H.; Williams, L.J.; Valentin, D. Multiple Factor Analysis: Principal Component Analysis for Multitable and Multiblock Data Sets. WIREs Comput. Stat. 2013, 5, 149–179. [Google Scholar] [CrossRef]
- Lavergne, C.; Hugoni, M.; Hubas, C.; Debroas, D.; Dupuy, C.; Agogué, H. Diel Rhythm Does Not Shape the Vertical Distribution of Bacterial and Archaeal 16S RRNA Transcript Diversity in Intertidal Sediments: A Mesocosm Study. Microb. Ecol. 2018, 75, 364–374. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed.; W. H. Freeman and Co: New York, NY, USA, 1995. [Google Scholar]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; Wiley Online Library: Hoboken, NJ, USA, 2017; pp. 1–15. [Google Scholar] [CrossRef]
- Tam, N.F.Y.; Ke, L.; Wang, X.H.; Wong, Y.S. Contamination of Polycyclic Aromatic Hydrocarbons in Surface Sediments of Mangrove Swamps. Environ. Pollut. 2001, 114, 255–263. [Google Scholar] [CrossRef]
- Remy, C.C.; Fleury, M.; Beauchêne, J.; Rivier, M.; Goli, T. Analysis of PAH Residues and Amounts of Phenols in Fish Smoked with Woods Traditionally Used in French Guiana. J. Ethnobiol. 2016, 36, 312–325. [Google Scholar] [CrossRef]
- Evans, K.M.; Gill, R.A.; Robotham, P.W.J. The PAH and Organic Content of Sediment Particle Size Fractions. Water. Air. Soil Pollut. 1990, 51, 13–31. [Google Scholar] [CrossRef]
- Arblaster, J.; Ikonomou, M.G.; Gobas, F.A. Toward Ecosystem-Based Sediment Quality Guidelines for Polychlorinated Biphenyls (PCBs). Integr. Environ. Assess. Manag. 2015, 11, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Pouch, A.; Zaborska, A.; Pazdro, K. The History of Hexachlorobenzene Accumulation in Svalbard Fjords. Environ. Monit. Assess. 2018, 190, 360. [Google Scholar] [CrossRef] [Green Version]
- Marchand, C.; Lallier-Vergès, E.; Baltzer, F.; Albéric, P.; Cossa, D.; Baillif, P. Heavy Metals Distribution in Mangrove Sediments along the Mobile Coastline of French Guiana. Mar. Chem. 2006, 98, 1–17. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, D.D.; Ingersoll, C.G.; Berger, T.A. Development and Evaluation of Consensus-Based Sediment Quality Guidelines for Freshwater Ecosystems. Arch. Environ. Contam. Toxicol. 2000, 39, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Burton, G., Jr. Allen Sediment Quality Criteria in Use around the World. Limnology 2002, 3, 65–76. [Google Scholar] [CrossRef]
- Conder, J.M.; Fuchsman, P.C.; Grover, M.M.; Magar, V.S.; Henning, M.H. Critical Review of Mercury Sediment Quality Values for the Protection of Benthic Invertebrates. Environ. Toxicol. Chem. 2015, 34, 6–21. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Llewellyn, C.A.; Egeland, E.S.; Johnsen, G. Phytoplankton Pigments: Characterization, Chemotaxonomy and Applications in Oceanography; Cambridge Environmental Chemistry Series; Cambridge University Press: Cambridge, UK, 2011; ISBN 978-1-107-00066-7. [Google Scholar]
- Hörtensteiner, S. Chlorophyll Degradation During Senescence. Annu. Rev. Plant Biol. 2006, 57, 55–77. [Google Scholar] [CrossRef]
- Fox, J.F. Intermediate-Disturbance Hypothesis. Science 1979, 204, 1344–1345. [Google Scholar] [CrossRef] [Green Version]
- Colen, C.V.; Montserrat, F.; Verbist, K.; Vincx, M.; Steyaert, M.; Vanaverbeke, J.; Herman, P.M.J.; Degraer, S.; Ysebaert, T. Tidal Flat Nematode Responses to Hypoxia and Subsequent Macrofauna-Mediated Alterations of Sediment Properties. Mar. Ecol. Prog. Ser. 2009, 381, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Carugati, L.; Gatto, B.; Rastelli, E.; Lo Martire, M.; Coral, C.; Greco, S.; Danovaro, R. Impact of Mangrove Forests Degradation on Biodiversity and Ecosystem Functioning. Sci. Rep. 2018, 8, 13298. [Google Scholar] [CrossRef] [Green Version]
- Alve, E. Benthic Foraminiferal Responses to Estuarine Pollution; a Review. J. Foraminifer. Res. 1995, 25, 190–203. [Google Scholar] [CrossRef]
- Mojtahid, M.; Jorissen, F.; Pearson, T.H. Comparison of Benthic Foraminiferal and Macrofaunal Responses to Organic Pollution in the Firth of Clyde (Scotland). Mar. Pollut. Bull. 2008, 56, 42–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, B.T.; Debenay, J.-P. Foraminifera, Environmental Bioindicators in the Highly ImpactedEnvironments of the Mekong Delta. Hydrobiologia 2005, 548, 75–83. [Google Scholar] [CrossRef]
- Bernhard, J.M.; Sen Gupta, B.K. Foraminifera of oxygen-depleted environments. In Modern Foraminifera; Sen Gupta, B.K., Ed.; Springer: Dordrecht, The Netherlands, 2003; ISBN 978-0-306-48104-8. [Google Scholar]
- Levin, L.A. Oxygen Minimum Zone Benthos: Adaptation and Community Response to Hypoxia. Oceanogr. Mar. Biol. Annu. Rev. 2003, 41, 46. [Google Scholar]
- Ostmann, A.; Nordhaus, I.; Sørensen, M.V. First Recording of Kinorhynchs from Java, with the Description of a New Brackish Water Species from a Mangrove-Fringed Lagoon. Mar. Biodivers. 2012, 42, 79–91. [Google Scholar] [CrossRef]
- Yamasaki, H.; Kajihara, H. A New Brackish-Water Species of Echinoderes (Kinorhyncha: Cyclorhagida) from the Seto Inland Sea, Japan. Species Divers. 2012, 17, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, M.V. First Account of Echinoderid Kinorhynchs from Brazil, with the Description of Three New Species. Mar. Biodivers. 2014, 44, 251–274. [Google Scholar] [CrossRef]
- Montagna, P.A.; Palmer, T.A.; Beseres Pollack, J. Summary: Water Supply, People, and the Future. In Hydrological Changes and Estuarine Dynamics; Montagna, P.A., Palmer, T.A., Beseres Pollack, J., Eds.; SpringerBriefs in Environmental Science; Springer: New York, NY, USA, 2013; pp. 79–81. ISBN 978-1-4614-5833-3. [Google Scholar]
- Alves, A.S.; Adão, H.; Ferrero, T.J.; Marques, J.C.; Costa, M.J.; Patrício, J. Benthic Meiofauna as Indicator of Ecological Changes in Estuarine Ecosystems: The Use of Nematodes in Ecological Quality Assessment. Ecol. Indic. 2013, 24, 462–475. [Google Scholar] [CrossRef]
- Ray, R.; Michaud, E.; Aller, R.C.; Vantrepotte, V.; Gleixner, G.; Walcker, R.; Devesa, J.; Le Goff, M.; Morvan, S.; Thouzeau, G. The Sources and Distribution of Carbon (DOC, POC, DIC) in a Mangrove Dominated Estuary (French Guiana, South America). Biogeochemistry 2018, 138, 297–321. [Google Scholar] [CrossRef]
- Ngo, Q.X.; Nguyen, N.C.; Nguyen, D.T.; Pham, V.L.; Vanreusel, A. Distribution Pattern of Free Living Nematode Communities in the Eight Mekong Estuaries by Seasonal Factor. J. Vietnam. Environ. 2013, 4, 28–33. [Google Scholar] [CrossRef]
- Haegerbaeumer, A.; Höss, S.; Heininger, P.; Traunspurger, W. Response of Nematode Communities to Metals and PAHs in Freshwater Microcosms. Ecotoxicol. Environ. Saf. 2018, 148, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, E.; Beyrem, H.; Aissa, P. Les Peuplements de Nématodes Libres, Indicateurs Du Degré d’anthropisation de La Lagune de Bou Ghrara (Tunisie). Vie et Milieu 2003, 53, 47–59. [Google Scholar]
- Sahraeian, N.; Sahafi, H.H.; Mosallanejad, H.; Ingels, J.; Semprucci, F. Temporal and Spatial Variability of Free-Living Nematodes in a Beach System Characterized by Domestic and Industrial Impacts (Bandar Abbas, Persian Gulf, Iran). Ecol. Indic. 2020, 118, 106697. [Google Scholar] [CrossRef]
- Semprucci, F.; Losi, V.; Moreno, M. A Review of Italian Research on Free-Living Marine Nematodes and the Future Perspectives on Their Use as Ecological Indicators (EcoInds). Mediterr. Mar. Sci. 2015, 16, 352–365. [Google Scholar] [CrossRef] [Green Version]
- Boufahja, F.; Semprucci, F.; Beyrem, H. An Experimental Protocol to Select Nematode Species from an Entire Community Using Progressive Sedimentary Enrichment. Ecol. Indic. 2016, 60, 292–309. [Google Scholar] [CrossRef]
- Armenteros, M.; Pérez-García, J.A.; Ruiz-Abierno, A.; Díaz-Asencio, L.; Helguera, Y.; Vincx, M.; Decraemer, W. Effects of Organic Enrichment on Nematode Assemblages in a Microcosm Experiment. Mar. Environ. Res. 2010, 70, 374–382. [Google Scholar] [CrossRef] [Green Version]
- Schratzberger, M.; Warwick, R. Differential Effects of Various Types of Disturbances on the Structure of Nematode Assemblages:An Experimental Approach. Mar. Ecol. Prog. Ser. 1999, 181, 227–236. [Google Scholar] [CrossRef]
- Soko, M.I.; Gyedu-Ababio, T.K. Free-Living Nematodes as Pollution Indicator in Incomati River Estuary, Mozambique. Open J. Ecol. 2019, 9, 117–133. [Google Scholar] [CrossRef] [Green Version]
- Steyaert, M.; Moodley, L.; Nadong, T.; Moens, T.; Soetaert, K.; Vincx, M. Responses of Intertidal Nematodes to Short-Term Anoxic Events. J. Exp. Mar. Biol. Ecol. 2007, 345, 175–184. [Google Scholar] [CrossRef]
- Warwick, R.M.; Gee, J.M. Community Structure of Estuarine Meiobenthos. Mar. Ecol. Prog. Ser. 1984, 18, 97–111. [Google Scholar] [CrossRef]
- Baldrighi, E.; Dovgal, I.; Zeppilli, D.; Abibulaeva, A.; Michelet, C.; Michaud, E.; Franzo, A.; Grassi, E.; Cesaroni, L.; Guidi, L.; et al. The Cost for Biodiversity: Records of Ciliate–Nematode Epibiosis with the Description of Three New Suctorian Species. Diversity 2020, 12, 224. [Google Scholar] [CrossRef]
- Fernandez-Leborans, G.; Chatterjee, T.; Grego, M. New Records of Epibiont Ciliates (Ciliophora) on Harpacticoida (Copepoda, Crustacea) from the Bay of Piran (Gulf of Trieste, Northern Adriatic). Cah Biol Mar 2012, 53, 53–63. [Google Scholar]
- Ansari, K.G.M.T.; Bhadury, P. Occurrence of Epibionts Associated with Meiofaunal Basibionts from the World’s Largest Mangrove Ecosystem, the Sundarbans. Mar. Biodivers. 2017, 47, 539–548. [Google Scholar] [CrossRef]
- Demopoulos, A.W.J.; Fry, B.; Smith, C.R. Food Web Structure in Exotic and Native Mangroves: A Hawaii-Puerto Rico Comparison. Oecologia 2007, 153, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Tietjen, J.H.; Deming, J.W.; Rowe, G.T.; Macko, S.; Wilke, R.J. Meiobenthos of the Hatteras Abyssal Plain and Puerto Rico Trench: Abundance, Biomass and Associations with Bacteria and Particulate Fluxes. Deep Sea Res. Part Oceanogr. Res. Pap. 1989, 36, 1567–1577. [Google Scholar] [CrossRef]
- Patrício, J.; Adão, H.; Neto, J.M.; Alves, A.S.; Traunspurger, W.; Marques, J.C. Do Nematode and Macrofauna Assemblages Provide Similar Ecological Assessment Information? Ecol. Indic. 2012, 14, 124–137. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station 1 | Station 2 | Station 3 | |
|---|---|---|---|
| Physico-chemical parameters | |||
| Ph | 6.26 ± 0.03 | 6.36 ± 0.11 | 6.02 ± 0.07 |
| Redox potential (mV) | 119 ± 8.51 | −90.1 ± 24.4 | 159 ± 5.78 |
| Granulometry (%) | |||
| Clay | 3.59 ± 0.40 | 8.34 ± 0.55 | 7.23 ± 0.44 |
| Silt | 75.3 ± 0.68 | 87.2 ± 1.33 | 88.3 ± 0.57 |
| Sand | 20.2 ± 0.34 | 4.20 ± 0.91 | 4.08 ± 0.93 |
| Carbon and Nitrogen | |||
| Total Organic Carbon (TOC %) | 0.04 ± 0.01 | 0.02 ± 0.002 | 0.01 ± 0.0005 |
| C:N (mol/mol) | 15.5 ± 0.86 | 10.3 ± 0.70 | 9.49 ± 0.18 |
| Prokaryotic community (number of 16S gene copies mg−1d.w.) | |||
| ARC 16S | 5.50×103 ± 1.87×103 | 1.18×104 ± 5.46×103 | 4.37×103 ± 2.56×103 |
| BAC 16S | 3.89×106 ± 1.40×106 | 5.76×105 ± 1.99×104 | 3.83×105 ± 6.80×104 |
| Pigments concentrations (µg cm−3) | |||
| Antheraxanthin (An) | 5.84 ± 1.29 | 20.7 ± 6.66 | 2.07 ± 0.52 |
| Beta-caroten (Bb) | 0.16 ± 0.06 | 1.06± 0.25 | 0.06 ± 0.04 |
| Canthaxanthin (Ct) | 0.38 ± 0.27 | 0.58 ± 0.38 | 0.29 ± 0.19 |
| Chlorophyll a (Ca) | 1.22 ± 0.42 | 5.20 ± 2.31 | 2.27 ± 0.71 |
| Chlorophyllb (Cb) | 0.14± 0.04 | 1.37 ± 0.58 | 0.16 ± 0.03 |
| Echinenone (Ec) | 0.21 ± 0.14 | 2.64 ± 1.76 | 0.20 ± 0.13 |
| Fucoxanthin (F) | 3.20 ± 0.89 | 11.4 ± 6.87 | 6.97 ± 4.53 |
| Lutein (L) | 1.68 ± 0.24 | 7.20 ± 1.75 | 1.03± 0.10 |
| Pheophorbidea (Pda) | 3.93± 1.56 | 6.32± 0.51 | 8.74± 1.95 |
| Pheophytin a (Pha) | 20.8 ± 4.95 | 47.0 ± 1.36 | 23.6± 0.86 |
| UnknownPigment 1 (UP1) | 1.51 ± 0.94 | 2.90 ± 2.46 | 1.83 ± 1.14 |
| Unknown Pigment 2 (UP2) | 1.66 ± 1.23 | 2.71 ± 1.80 | 2.38 ± 0.82 |
| UnknownPigment 3 (UP3) | 0.91 ± 0.24 | 4.15 ± 0.86 | 2.01 ± 0.56 |
| UnknownPigment 4 (UP4) | 0.15 ± 0.20 | 1.21 ± 0.23 | 0.48 ± 0.34 |
| UnknownPigment 5 (UP5) | 1.13 ± 0.26 | 0.89 ± 0.29 | 0.92 ± 0.34 |
| UnknownPigment 6 (UP6) | 0.38 ± 0.21 | 0.45 ± 0.17 | 0.26 ± 0.15 |
| UnknownPigment 7 (UP7) | 0.23 ± 0.07 | 0.44 ± 0.01 | 0.33 ± 0.06 |
| Violaxanthin (Vio) | 1.65 ± 0.97 | 4.06 ± 1.08 | 1.41 ± 0.23 |
| Concentrations of organic contaminants (ng g−1) | |||
| Hydrocarbons | 48.6 ± 17.6 | 212 ± 20.3 | 250 ± 79.0 |
| Pesticides | 11.9 ± 1.81 | 10.8 ± 0.87 | 0.03 ± 0.04 |
| PCB | 3.43 ± 0.31 | 4.95 ± 2.48 | 0.21 ± 0.28 |
| Plasticizers | 0.00 | 0.45 ± 0.60 | 0.28 ± 0.21 |
| Concentrations of metals ad metalloids (µg g−1) | |||
| Aluminum (Al)(mg g−1) | 110 ± 1.86 | 115 ± 1.78 | 118 ± 1.37 |
| Antimony (Sb) | 0.64 ± 0.07 | 0.54 ± 0.02 | 0.56 ± 0.002 |
| Arsenic (As) | 25.9 ± 1.28 | 22.7 ± 1.25 | 21.3 ± 0.31 |
| Barium (Ba) | 400 ± 6.27 | 416 ± 6.76 | 440 ± 4.79 |
| Beryllium (Be) | 3.13 ± 0.10 | 3.24 ± 0.07 | 3.32 ± 0.04 |
| Bismuth (Bi) | 0.59 ± 0.01 | 0.53 ± 0.01 | 0.55 ± 0.01 |
| Cadmium (Cd) | 0.08 ± 0.01 | 0.07 ± 0.005 | 0.07 ± 0.002 |
| Cesium (Cs) | 11.5 ± 0.08 | 11.9 ± 0.24 | 12.4 ± 0.14 |
| Chromium (Cr) | 78.6 ± 2.75 | 84.5 ± 1.50 | 87.5 ± 0.73 |
| Cobalt (Co) | 20.7 ± 2.14 | 21.3 ± 1.32 | 21.8 ± 0.18 |
| Copper (Cu) | 29.6 ± 1.79 | 29.4 ± 0.65 | 29.8 ± 0.47 |
| Iron (Fe) (mg g-1) | 63.0 ± 3.31 | 64.2 ± 4.71 | 65.0 ± 2.85 |
| Lead (Pb) | 36.8 ± 1.11 | 32.6 ± 0.60 | 34.7 ± 0.23 |
| Lithium (Li) | 83.4 ±2.23 | 86.3 ± 1.43 | 87.8 ± 1.12 |
| Manganese (Mn) | 420 ± 41.3 | 862 ± 182 | 992 ± 87.4 |
| Mercury (Hg) (ng g-1) | 59.7 ± 2.70 | 47.0 ± 0.05 | 51.0 ± 1.11 |
| Molybdenum (Mo) | 2.12 ± 0.09 | 1.31 ± 0.09 | 1.43 ± 0.01 |
| Nickel (Ni) | 36.7 ± 1.31 | 38.5 ± 0.72 | 38.8 ± 0.35 |
| Rubidium (Rb) | 129 ± 1.47 | 200 ± 93.7 | 132 ± 1.36 |
| Silver (Ag) | 0.23 ± 0.01 | 0.16 ± 0.02 | 0.19 ± 0.01 |
| Strontium (Sr) | 122 ± 14.6 | 116 ± 0.99 | 114 ± 1.09 |
| Thallium (Tl) | 0.92 ± 0.02 | 0.92 ± 0.02 | 0.97 ± 0.01 |
| Tin (Sn) | 3.58 ± 0.07 | 3.70 ± 0.08 | 3.83 ± 0.04 |
| Titanium (Ti)(mg g-1) | 2.70 ± 0.44 | 2.80 ± 0.26 | 2.86 ± 0.13 |
| Uranium (U) | 2.25 ± 0.10 | 1.78 ± 0.21 | 2.23 ± 0.02 |
| Vanadium (V) | 153 ± 5.9 | 161 ± 3.50 | 167 ± 1.49 |
| Zinc (Zn) | 136 ± 9.07 | 148 ± 3.01 | 156 ± 1.48 |
| Family | Genus | AbbreViation | Station 1 | Station 2 | Station 3 | |||
|---|---|---|---|---|---|---|---|---|
| Density ind.10cm−2 | Relative Density (%) | Density ind.10cm−2 | Relative Density (%) | Density ind.10cm−2 | Relative Density (%) | |||
| Anoplostomatidae | Anoplostoma | Ano | 113 ± 71 | 5 ± 3 | 0 | 0 | 452 ± 128 | 7 ± 0.5 |
| Axonolaimidae | Parodontophora | Pad | 28 ± 19 | 12 ± 8 | 0 | 0 | 294 ± 43 | 5 ± 1 |
| Camacolaimidae | Deontolaimus | Deo | 99 ± 55 | 4 ± 2 | 0 | 0 | 508 ± 79 | 9± 3 |
| Chromadoridae | Hypodontolaimus | Hyp | 130± 63 | 6 ± 3 | 0 | 0 | 0 | 0 |
| Neochromadora | Neo | 56 ± 27 | 2 ± 1 | 73 ± 69 | 1 ± 1 | 8 ± 10 | 0.2 ± 0.2 | |
| Ptycholaimellus | Pty | 0 | 0 | 214 ± 121 | 3 ± 2 | 100± 52 | 2 ± 1 | |
| Desmodoridae | Desmodora | Dea | 648 ± 158 | 27 ± 6 | 912 ± 517 | 14.5 ± 7 | 293 ± 132 | 6 ± 3 |
| Spirinia | Spi | 252 ± 154 | 10 ± 6 | 343 ± 92 | 7 ± 4 | 0 | 0 | |
| Diplopeltidae | Southerniella | Sou | 7 ± 9 | 0.3 ± 0.4 | 0 | 0 | 163 ± 94 | 2 ± 1 |
| Ethmolaimidae | Paraethmolaimus | Pae | 114 ± 118 | 5 ± 5 | 0 | 0 | 0 | 0 |
| Leptosomatidae | Pseudocella | Psc | 77 ± 21 | 3 ± 1 | 39 ± 52 | 1 ± 1 | 670± 275 | 10 ± 3 |
| Linhomoeidae | Metalinhomoeus | Mel | 0 | 0 | 193 ± 107 | 3 ± 2 | 0 | 0 |
| Paralinhomoeus | Pal | 5 ± 6 | 0.2 ± 0.2 | 164 ± 109 | 2.5 ± 2 | 61 ± 58 | 1 ± 1 | |
| Terschellingia | Ter | 96 ± 47 | 4 ± 2 | 1410± 525 | 28 ± 12 | 14.5 ± 19 | 3 ± 4 | |
| Microlaimidae | Microlaimus | Mic | 15 ± 8 | 1 ± 0.4 | 367 ± 248 | 8 ± 7 | 1178 ± 560 | 17 ± 6.5 |
| Molgolaimus | Mol | 5 ± 6 | 0.2 ± 0.3 | 140 ± 102 | 2 ± 2 | 39 ± 17 | 1 ± 0.1 | |
| Oxystominidae | Halalaimus | Hal | 185 ± 41 | 8 ± 2 | 144 ± 96 | 2 ± 1.5 | 583 ± 273 | 9 ± 2 |
| Selachinematidae | Richtersia | Ric | 5 ± 6 | 0.2 ± 0.3 | 245 ± 198 | 4 ± 3 | 0 | 0 |
| Sphaerolaimidae | Metasphaerolaimus | Mes | 320± 220 | 1 ± 1 | 0 | 0 | 151 ± 70 | 2.5 ± 1 |
| Sphaerolaimus | Sph | 64 ± 40 | 3 ± 2 | 39 ± 52 | 1 ± 1 | 222 ± 84 | 3.5 ± 1 | |
| Subsphaerolaimus | Sub | 48 ± 17 | 2 ± 1 | 0 | 0 | 7± 6 | 1 ± 1 | |
| Xyalidae | Daptonema | Dap | 0 | 0 | 183 ± 122 | 3 ± 2 | 84 ± 62 | 1 ± 1 |
| Cobbia | Cob | 36 ± 10 | 1.5 ± 0.5 | 65 ± 33 | 1 ± 1 | 19 ± 8 | 3 ± 1 | |
| Elzalia | Elz | 109 ± 34 | 5 ± 2 | 43 ± 50 | 1 ± 1 | 263 ± 105 | 4 ± 1 | |
| Zygonemella | Zyg | 8 ± 4 | 3 ± 1.5 | 287 ± 191 | 4 ± 3 | 35 ± 23 | 1 ± 0.4 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michelet, C.; Zeppilli, D.; Hubas, C.; Baldrighi, E.; Cuny, P.; Dirberg, G.; Militon, C.; Walcker, R.; Lamy, D.; Jézéquel, R.; et al. First Assessment of the Benthic Meiofauna Sensitivity to Low Human-Impacted Mangroves in French Guiana. Forests 2021, 12, 338. https://doi.org/10.3390/f12030338
Michelet C, Zeppilli D, Hubas C, Baldrighi E, Cuny P, Dirberg G, Militon C, Walcker R, Lamy D, Jézéquel R, et al. First Assessment of the Benthic Meiofauna Sensitivity to Low Human-Impacted Mangroves in French Guiana. Forests. 2021; 12(3):338. https://doi.org/10.3390/f12030338
Chicago/Turabian StyleMichelet, Claire, Daniela Zeppilli, Cédric Hubas, Elisa Baldrighi, Philippe Cuny, Guillaume Dirberg, Cécile Militon, Romain Walcker, Dominique Lamy, Ronan Jézéquel, and et al. 2021. "First Assessment of the Benthic Meiofauna Sensitivity to Low Human-Impacted Mangroves in French Guiana" Forests 12, no. 3: 338. https://doi.org/10.3390/f12030338
APA StyleMichelet, C., Zeppilli, D., Hubas, C., Baldrighi, E., Cuny, P., Dirberg, G., Militon, C., Walcker, R., Lamy, D., Jézéquel, R., Receveur, J., Gilbert, F., Houssainy, A. E., Dufour, A., Heimbürger-Boavida, L. -E., Bihannic, I., Sylvi, L., Vivier, B., & Michaud, E. (2021). First Assessment of the Benthic Meiofauna Sensitivity to Low Human-Impacted Mangroves in French Guiana. Forests, 12(3), 338. https://doi.org/10.3390/f12030338