
What Factors Shape Spatial Distribution of Biomass in Riparian Forests? Insights from a LiDAR Survey over a Large Area

 , ,
, ,
Abstract
:1. Introduction
- To propose an approach to estimate aboveground biomass at tree level, and to compare two biomass estimates based on variables from a raster-format LiDAR CHM or from the original LiDAR point cloud;
- To highlight environmental factors structuring biomass distribution in riparian forests at a sub-regional scale (200 km of river and their riparian zone) in the context of low-energy temperate rivers (Meuse catchment, Western Europe).
2. Materials and Methods
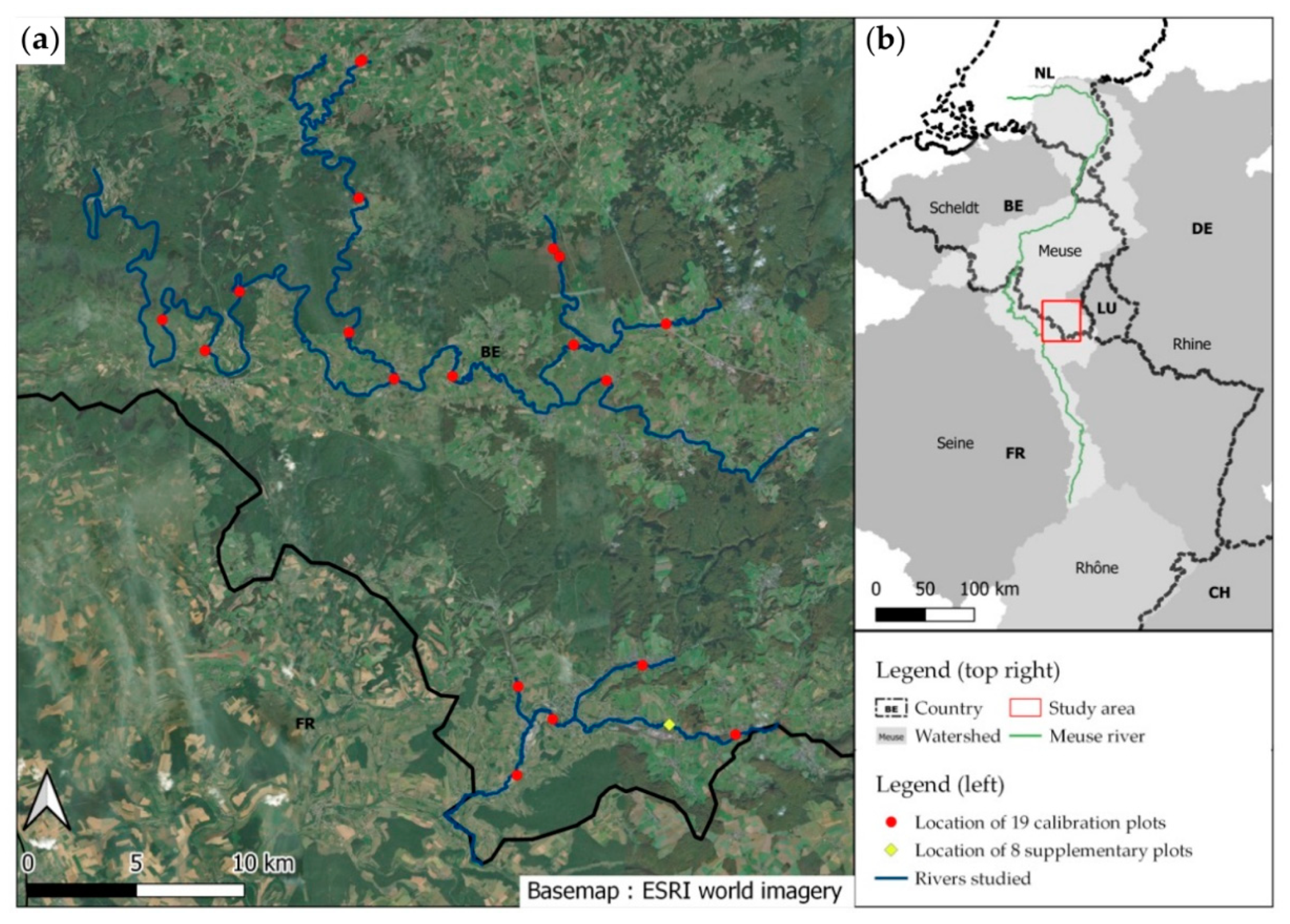
2.1. Study Area and Available Data
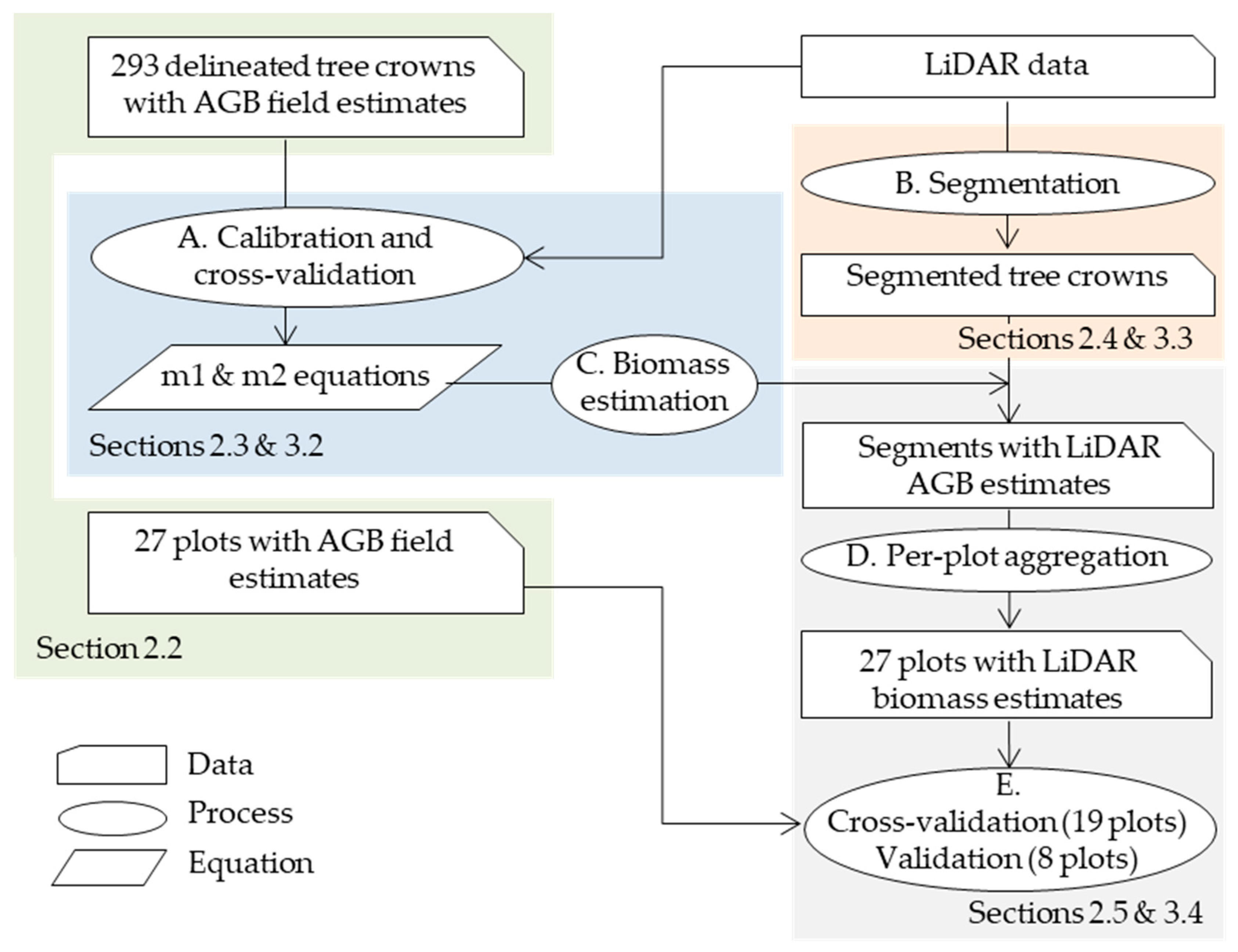
2.2. Biomass Field Data and Equations
2.3. Biomass Prediction from LiDAR Data at Tree Level
2.4. Individual Tree Segmentation
2.5. Validation at Plot Level
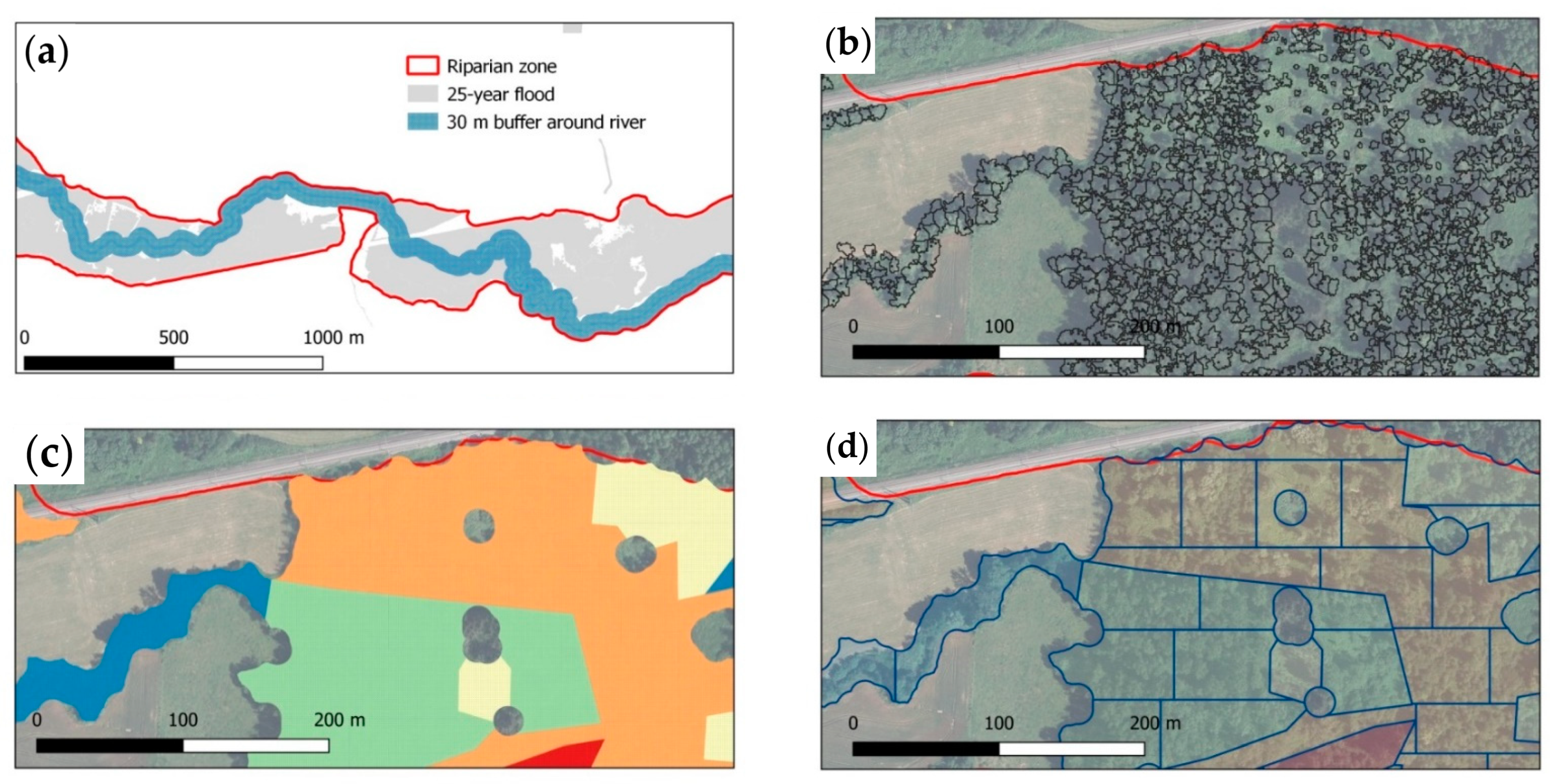
2.6. Riparian Forest Delineation and Upscaling of Biomass Prediction
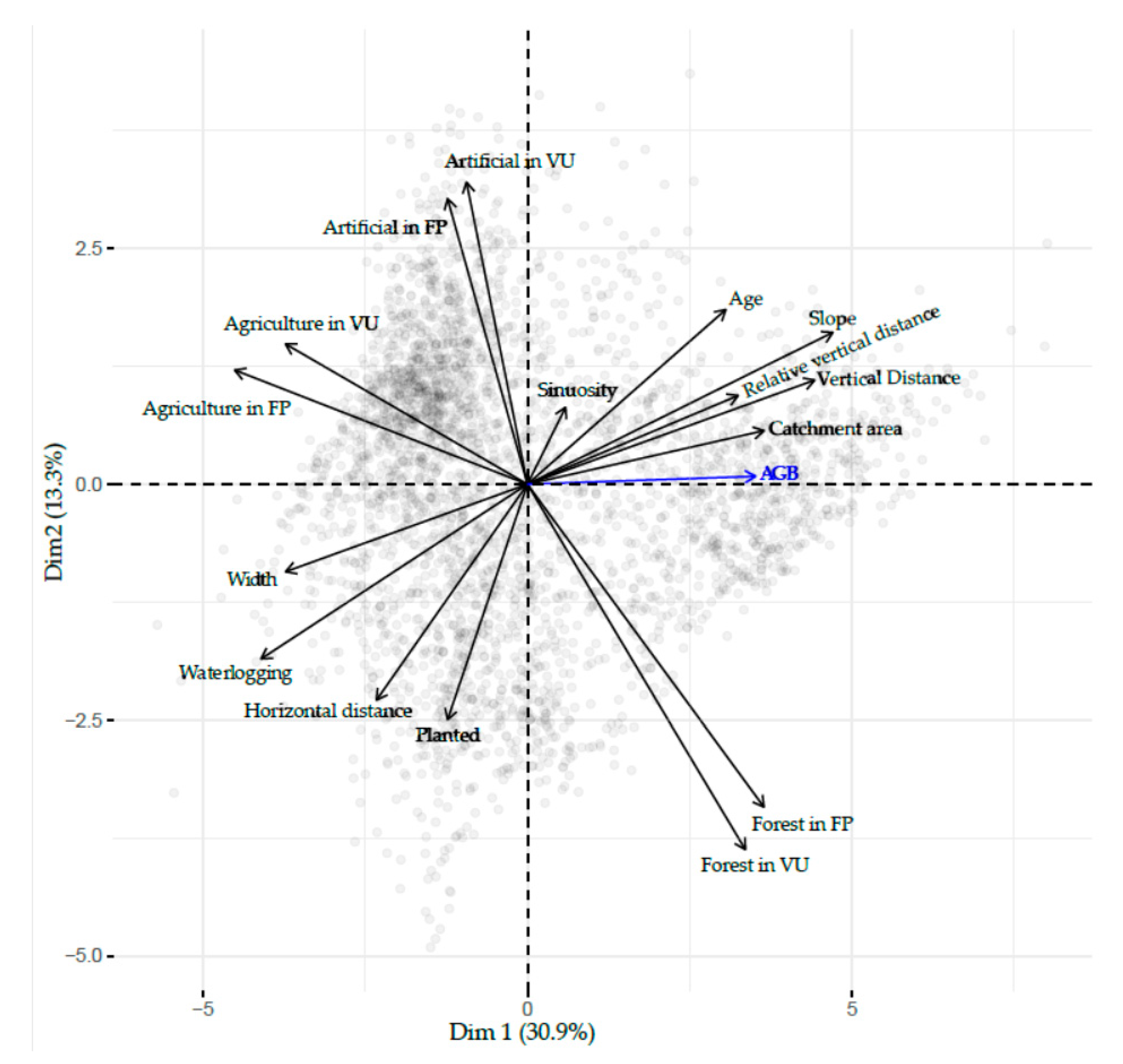
2.7. Analysis of Environmental Factors Structuring the Spatial Distribution of Biomass
3. Results
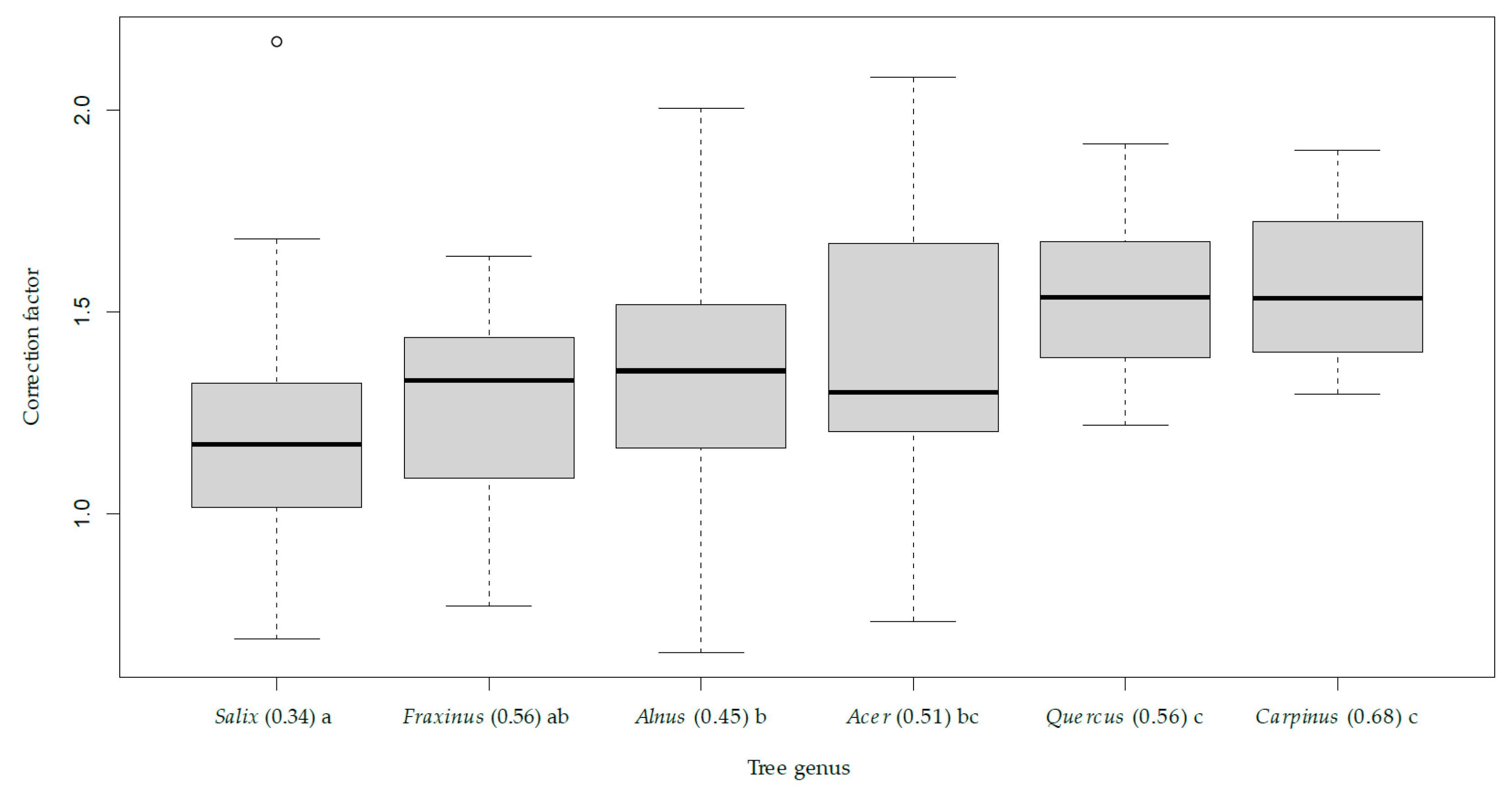
3.1. Volume Equations for Alnus and Salix
3.2. Biomass Prediction from LiDAR Data at Tree Level
3.3. Individual Tree Segmentation
3.4. Validation at Plot Level
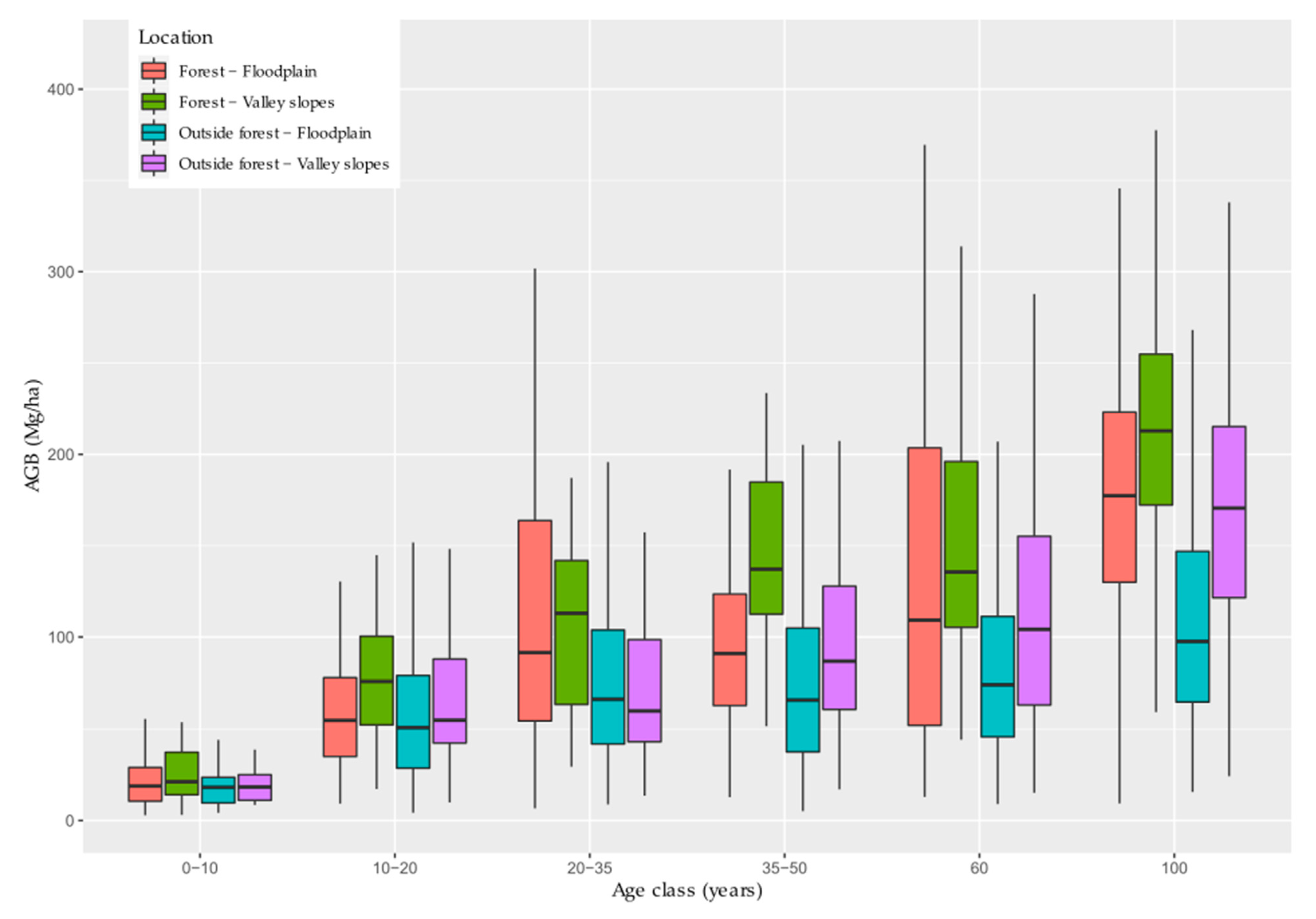
3.5. Analysis of Environmental Factors Structuring Spatial Distribution of Biomass
4. Discussion
4.1. LiDAR Biomass Estimates
4.2. Spatial Distribution of Biomass and Influencing Factors
4.3. Perspectives for Generalization of the Approach
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naiman, R.J.; Decamps, H.; Pollock, M. The Role of Riparian Corridors in Maintaining Regional Biodiversity. Ecol. Appl. 1993, 3, 209–212. [Google Scholar] [CrossRef]
- Riis, T.; Kelly-Quinn, M.; Aguiar, F.C.; Manolaki, P.; Bruno, D.; Bejarano, M.D.; Clerici, N.; Fernandes, M.R.; Franco, J.C.; Pettit, N.; et al. Global Overview of Ecosystem Services Provided by Riparian Vegetation. BioScience 2020, 70, 501–514. [Google Scholar] [CrossRef]
- Stella, J.C.; Bendix, J. Chapter 5—Multiple Stressors in Riparian Ecosystems. In Multiple Stressors in River Ecosystems; Sabater, S., Elosegi, A., Ludwig, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 81–110. ISBN 978-0-12-811713-2. [Google Scholar] [CrossRef]
- González, E.; Sher, A.A.; Tabacchi, E.; Masip, A.; Poulin, M. Restoration of riparian vegetation: A global review of implementation and evaluation approaches in the international, peer-reviewed literature. J. Environ. Manag. 2015, 158, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Dybala, K.E.; Matzek, V.; Gardali, T.; Seavy, N.E. Carbon sequestration in riparian forests: A global synthesis and meta-analysis. Glob. Chang. Biol. 2019, 25, 57–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzek, V.; Stella, J.; Ropion, P. Development of a carbon calculator tool for riparian forest restoration. Appl. Veg. Sci. 2018, 21, 584–594. [Google Scholar] [CrossRef]
- Sutfin, N.A.; Wohl, E.E.; Dwire, K.A. Banking carbon: A review of organic carbon storage and physical factors influencing retention in floodplains and riparian ecosystems. Earth Surf. Process. Landf. 2016, 41, 38–60. [Google Scholar] [CrossRef]
- Balian, E.V.; Naiman, R.J. Abundance and Production of Riparian Trees in the Lowland Floodplain of the Queets River, Washington. Ecosystems 2005, 8, 841–861. [Google Scholar] [CrossRef]
- Keeton, W.S.; Kraft, C.E.; Warren, D.R. Mature and Old-Growth Riparian Forests: Structure, Dynamics, and Effects on Adirondack Stream Habitats. Ecol. Appl. 2007, 17, 852–868. [Google Scholar] [CrossRef]
- Dosskey, M.G.; Vidon, P.; Gurwick, N.P.; Allan, C.J.; Duval, T.P.; Lowrance, R. The Role of Riparian Vegetation in Protecting and Improving Chemical Water Quality in Streams1. JAWRA J. Am. Water Resour. Assoc. 2010, 46, 261–277. [Google Scholar] [CrossRef]
- Tufekcioglu, A.; Raich, J.W.; Isenhart, T.M.; Schultz, R.C. Biomass, carbon and nitrogen dynamics of multi-species riparian buffers within an agricultural watershed in Iowa, USA. Agrofor. Syst. 2003, 57, 187–198. [Google Scholar] [CrossRef]
- Matzek, V.; Lewis, D.; O’Geen, A.; Lennox, M.; Hogan, S.D.; Feirer, S.T.; Eviner, V.; Tate, K.W. Increases in soil and woody biomass carbon stocks as a result of rangeland riparian restoration. Carbon Balance Manag. 2020, 15, 16. [Google Scholar] [CrossRef] [PubMed]
- Forzieri, G.; Castelli, F.; Preti, F. Advances in remote sensing of hydraulic roughness. Int. J. Remote Sens. 2012, 33, 630–654. [Google Scholar] [CrossRef]
- Dybala, K.E.; Steger, K.; Walsh, R.G.; Smart, D.R.; Gardali, T.; Seavy, N.E. Optimizing carbon storage and biodiversity co-benefits in reforested riparian zones. J. Appl. Ecol. 2019, 56, 343–353. [Google Scholar] [CrossRef]
- Dufour, S.; Piégay, H. Geomorphological Controls of Fraxinus Excelsior Growth and Regeneration in Floodplain Forests. Ecology 2008, 89, 205–215. [Google Scholar] [CrossRef]
- Megonigal, J.P.; Conner, W.H.; Kroeger, S.; Sharitz, R.R. Aboveground Production in Southeastern Floodplain Forests: A Test of the Subsidy–Stress Hypothesis. Ecology 1997, 78, 370–384. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-González, P.M.; Stella, J.C.; Campelo, F.; Ferreira, M.T.; Albuquerque, A. Subsidy or stress? Tree structure and growth in wetland forests along a hydrological gradient in Southern Europe. For. Ecol. Manag. 2010, 259, 2015–2025. [Google Scholar] [CrossRef]
- Marks, C.O.; Yellen, B.C.; Wood, S.A.; Martin, E.H.; Nislow, K.H. Variation in Tree Growth along Soil Formation and Microtopographic Gradients in Riparian Forests. Wetlands 2020. [Google Scholar] [CrossRef]
- Kramer, K.; Vreugdenhil, S.J.; van der Werf, D.C. Effects of flooding on the recruitment, damage and mortality of riparian tree species: A field and simulation study on the Rhine floodplain. For. Ecol. Manag. 2008, 255, 3893–3903. [Google Scholar] [CrossRef]
- Wohl, E.; Dwire, K.; Sutfin, N.; Polvi, L.; Bazan, R. Mechanisms of carbon storage in mountainous headwater rivers. Nat. Commun. 2012, 3, 1263. [Google Scholar] [CrossRef] [Green Version]
- Lucas, C.M.; Schöngart, J.; Sheikh, P.; Wittmann, F.; Piedade, M.T.F.; McGrath, D.G. Effects of land-use and hydroperiod on aboveground biomass and productivity of secondary Amazonian floodplain forests. For. Ecol. Manag. 2014, 319, 116–127. [Google Scholar] [CrossRef]
- Michez, A.; Piégay, H.; Lejeune, P.; Claessens, H. Multi-temporal monitoring of a regional riparian buffer network (>12,000 km) with LiDAR and photogrammetric point clouds. J. Environ. Manag. 2017, 202, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Wasser, L.; Chasmer, L.; Day, R.; Taylor, A. Quantifying land use effects on forested riparian buffer vegetation structure using LiDAR data. Ecosphere 2015, 6, art10. [Google Scholar] [CrossRef]
- da Silva, R.L.; Leite, M.F.A.; Muniz, F.H.; de Souza, L.A.G.; de Moraes, F.H.R.; Gehring, C. Degradation impacts on riparian forests of the lower Mearim river, eastern periphery of Amazonia. For. Ecol. Manag. 2017, 402, 92–101. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Aguiar, F.C.; Martins, M.J.; Rico, N.; Ferreira, M.T.; Correia, A.C. Carbon Stock Estimations in a Mediterranean Riparian Forest: A Case Study Combining Field Data and UAV Imagery. Forests 2020, 11, 376. [Google Scholar] [CrossRef] [Green Version]
- Lu, D. The potential and challenge of remote sensing-based biomass estimation. Int. J. Remote Sens. 2006, 27, 1297–1328. [Google Scholar] [CrossRef]
- Goetz, S.; Dubayah, R. Advances in remote sensing technology and implications for measuring and monitoring forest carbon stocks and change. Carbon Manag. 2011, 2, 231–244. [Google Scholar] [CrossRef]
- Huylenbroeck, L.; Laslier, M.; Dufour, S.; Georges, B.; Lejeune, P.; Michez, A. Using remote sensing to characterize riparian vegetation: A review of available tools and perspectives for managers. J. Environ. Manag. 2020, 267, 110652. [Google Scholar] [CrossRef]
- Mendez-Estrella, R.; Romo-Leon, J.R.; Castellanos, A.E. Mapping Changes in Carbon Storage and Productivity Services Provided by Riparian Ecosystems of Semi-Arid Environments in Northwestern Mexico. ISPRS Int. J. Geo Inf. 2017, 6, 298. [Google Scholar] [CrossRef]
- Husson, E.; Lindgren, F.; Ecke, F. Assessing Biomass and Metal Contents in Riparian Vegetation Along a Pollution Gradient Using an Unmanned Aircraft System. Water Air Soil Pollut. 2014, 225, 1957. [Google Scholar] [CrossRef]
- Mitchard, E.T.A.; Saatchi, S.S.; White, L.J.T.; Abernethy, K.A.; Jeffery, K.J.; Lewis, S.L.; Collins, M.; Lefsky, M.A.; Leal, M.E.; Woodhouse, I.H.; et al. Mapping tropical forest biomass with radar and spaceborne LiDAR in Lopé National Park, Gabon: Overcoming problems of high biomass and persistent cloud. Biogeosciences 2012, 9, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Filippi, A.M.; Güneralp, İ.; Randall, J. Hyperspectral remote sensing of aboveground biomass on a river meander bend using multivariate adaptive regression splines and stochastic gradient boosting. Remote Sens. Lett. 2014, 5, 432–441. [Google Scholar] [CrossRef]
- Forzieri, G. Satellite retrieval of woody biomass for energetic reuse of riparian vegetation. Biomass Bioenergy 2012, 36, 432–438. [Google Scholar] [CrossRef]
- Suchenwirth, L.; Stümer, W.; Schmidt, T.; Förster, M.; Kleinschmit, B. Large-Scale Mapping of Carbon Stocks in Riparian Forests with Self-Organizing Maps and the k-Nearest-Neighbor Algorithm. Forests 2014, 5, 1635–1652. [Google Scholar] [CrossRef] [Green Version]
- Fassnacht, F.E.; Hartig, F.; Latifi, H.; Berger, C.; Hernández, J.; Corvalán, P.; Koch, B. Importance of sample size, data type and prediction method for remote sensing-based estimations of aboveground forest biomass. Remote Sens. Environ. 2014, 154, 102–114. [Google Scholar] [CrossRef]
- Laslier, M.; Hubert-Moy, L.; Dufour, S. Mapping Riparian Vegetation Functions Using 3D Bispectral LiDAR Data. Water 2019, 11, 483. [Google Scholar] [CrossRef] [Green Version]
- Zolkos, S.G.; Goetz, S.J.; Dubayah, R. A meta-analysis of terrestrial aboveground biomass estimation using lidar remote sensing. Remote Sens. Environ. 2013, 128, 289–298. [Google Scholar] [CrossRef]
- Cartisano, R.; Mattioli, W.; Corona, P.; Mugnozza, G.S.; Sabatti, M.; Ferrari, B.; Cimini, D.; Giuliarelli, D. Assessing and mapping biomass potential productivity from poplar-dominated riparian forests: A case study. Biomass Bioenergy 2013, 54, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Karrenberg, S.; Edwards, P.J.; Kollmann, J. The life history of Salicaceae living in the active zone of floodplains. Freshw. Biol. 2002, 47, 733–748. [Google Scholar] [CrossRef]
- Naiman, R.J.; Decamps, H.; McClain, M.E. Riparia: Ecology, Conservation, and Management of Streamside Communities; Elsevier: Amsterdam, The Netherlands, 2005; ISBN 978-0-08-047068-9. [Google Scholar]
- Dalponte, M.; Coomes, D.A. Tree-centric mapping of forest carbon density from airborne laser scanning and hyperspectral data. Methods Ecol. Evol. 2016, 7, 1236–1245. [Google Scholar] [CrossRef] [Green Version]
- Cadol, D.; Wine, M.L. Geomorphology as a first order control on the connectivity of riparian ecohydrology. Geomorphology 2017, 277, 154–170. [Google Scholar] [CrossRef]
- Gob, F.; Houbrechts, G.; Hiver, J.M.; Petit, F. River dredging, channel dynamics and bedload transport in an incised meandering river (the River Semois, Belgium). River Res. Appl. 2005, 21, 791–804. [Google Scholar] [CrossRef]
- Service Public Wallonie. Nuage de Points LIDAR 2013–2014. Available online: http://geoportail.wallonie.be/catalogue/cd7578ef-c726-46cb-a29e-a90b3d4cd368.html (accessed on 5 March 2021).
- Service Public Wallonie. Notice Méthodologique D’élaboration des Cartographies des Zones Soumises À L’aléa D’inondation et du Risque de Dommages dus Aux Inondations. Available online: http://environnement.wallonie.be/inondations/files/2016_carto/Methodo_GW20160310_final.pdf (accessed on 14 December 2020).
- Corral-Rivas, S.; Álvarez-González, J.G.; Crecente-Campo, F.; Corral-Rivas, J.J. Local and generalized height-diameter models with random parameters for mixed, uneven-aged forests in Northwestern Durango, Mexico. For. Ecosyst. 2014, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, K.; Alavi, S.J.; Kouchaksaraei, M.T. Constructing site quality curves and productivity assessment for uneven-aged and mixed stands of oriental beech (Fagus oriental Lipsky) in Hyrcanian forest, Iran. For. Sci. Technol. 2017, 13, 41–46. [Google Scholar] [CrossRef]
- Zanne, A.; Lopez-Gonzalez, G.; Coomes, D.; Ilic, J.; Jansen, S.; Lewis, S.; Miller, R.; Swenson, N.; Wiemann, M.; Chave, J. Data from: Towards a worldwide wood economics spectrum. Dryad Digit Repos. Dryad 2009. [Google Scholar] [CrossRef]
- Dagnelie, P.; Palm, R.; Rondeux, J. Cubage des Arbres et des Peuplements Forestiers. Tables et Équations; Presses Agronomiques de Gembloux: Gembloux, Belgium, 2013. [Google Scholar]
- Longuetaud, F.; Santenoise, P.; Mothe, F.; Senga Kiessé, T.; Rivoire, M.; Saint-André, L.; Ognouabi, N.; Deleuze, C. Modeling volume expansion factors for temperate tree species in France. For. Ecol. Manag. 2013, 292, 111–121. [Google Scholar] [CrossRef]
- Zianis, D.; Muukkonen, P.; Mäkipää, R.; Mencuccini, M. Biomass and Stem Volume Equations for Tree Species in Europe; Finnish Society of Forest Science: Helsinki, Finland, 2005; ISBN 978-951-40-1983-8. [Google Scholar]
- Baskerville, G.L. Use of Logarithmic Regression in the Estimation of Plant Biomass. Can. J. For. Res. 2011. [Google Scholar] [CrossRef]
- Roussel, J.-R.; Auty, D.; De Boissieu, F.; Sánchez Meador, A.; Bourdon, J.-F.; Gatziolis, D. lidR: Airborne LiDAR Data Manipulation and Visualization for Forestry Applications. 2020; R package, version 2.2.4. [Google Scholar]
- Lamar, W.R.; McGraw, J.B.; Warner, T.A. Multitemporal censusing of a population of eastern hemlock (Tsuga canadensis L.) from remotely sensed imagery using an automated segmentation and reconciliation procedure. Remote Sens. Environ. 2005, 94, 133–143. [Google Scholar] [CrossRef]
- Gurnell, A.M.; Corenblit, D.; de Jalón, D.G.; del Tánago, M.G.; Grabowski, R.C.; O’Hare, M.T.; Szewczyk, M. A Conceptual Model of Vegetation–hydrogeomorphology Interactions Within River Corridors. River Res. Appl. 2016, 32, 142–163. [Google Scholar] [CrossRef]
- Clerici, N.; Weissteiner, C.J.; Paracchini, M.L.; Boschetti, L.; Baraldi, A.; Strobl, P. Pan-European distribution modelling of stream riparian zones based on multi-source Earth Observation data. Ecol. Indic. 2013, 24, 211–223. [Google Scholar] [CrossRef]
- Radoux, J.; Bourdouxhe, A.; Coos, W.; Dufrêne, M.; Defourny, P. Improving Ecotope Segmentation by Combining Topographic and Spectral Data. Remote Sens. 2019, 11, 354. [Google Scholar] [CrossRef] [Green Version]
- Huck, J. jonnyhuck/RFCL-PolygonDivider. 2020; QGIS plugin, version 0.6. [Google Scholar]
- Service Public Wallonie. Carte Numérique des Sols de Wallonie—Série. Available online: http://geoportail.wallonie.be/catalogue/c5bedf2b-1cac-4231-9d9a-854e0ef2c9ce.html (accessed on 15 December 2020).
- Service Public Wallonie. Occupation et Utilisation du sol en Wallonie—COSW 2007—Série—Donnée Historique. Available online: http://geoportail.wallonie.be/catalogue/290e1fe8-0d99-410e-967b-a02f389b931a.html (accessed on 14 December 2020).
- Kreuzwieser, J.; Papadopoulou, E.; Rennenberg, H. Interaction of Flooding with Carbon Metabolism of Forest Trees. Plant Biol. 2004, 6, 299–306. [Google Scholar] [CrossRef]
- Singer, M.B.; Stella, J.C.; Dufour, S.; Piégay, H.; Wilson, R.J.S.; Johnstone, L. Contrasting water-uptake and growth responses to drought in co-occurring riparian tree species. Ecohydrology 2013, 6, 402–412. [Google Scholar] [CrossRef]
- Schifman, L.A.; Stella, J.C.; Volk, T.A.; Teece, M.A. Carbon isotope variation in shrub willow (Salix spp.) ring-wood as an indicator of long-term water status, growth and survival. Biomass Bioenergy 2012, 36, 316–326. [Google Scholar] [CrossRef]
- Alber, A.; Piégay, H. Spatial disaggregation and aggregation procedures for characterizing fluvial features at the network-scale: Application to the Rhône basin (France). Geomorphology 2011, 125, 343–360. [Google Scholar] [CrossRef]
- Christophe, R.; Samuel, D. Fluvial Corridor Toolbox QGis Plugin; Zenodo: Genève, Switzerland, 2020. [Google Scholar] [CrossRef]
- Camporeale, C.; Ridolfi, L. Interplay among river meandering, discharge stochasticity and riparian vegetation. J. Hydrol. 2010, 382, 138–144. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2020; R package, version 2.5-7. [Google Scholar]
- Lindeman, R.; Merenda, P.; Gold, R. Introduction to Bivariate and Multivariate Analysis; Scott Foresman: Chicago, IL, USA, 1980. [Google Scholar]
- Groemping, U.; Lehrkamp, M. Relaimpo: Relative Importance of Regressors in Linear Models. 2018; R package, version 2.2-3. [Google Scholar]
- Garcia, M.; Saatchi, S.; Ferraz, A.; Silva, C.A.; Ustin, S.; Koltunov, A.; Balzter, H. Impact of data model and point density on aboveground forest biomass estimation from airborne LiDAR. Carbon Balance Manag. 2017, 12, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chirici, G.; McRoberts, R.E.; Fattorini, L.; Mura, M.; Marchetti, M. Comparing echo-based and canopy height model-based metrics for enhancing estimation of forest aboveground biomass in a model-assisted framework. Remote Sens. Environ. 2016, 174, 1–9. [Google Scholar] [CrossRef]
- Holmgren, J.; Nilsson, M.; Olsson, H. Simulating the effects of lidar scanning angle for estimation of mean tree height and canopy closure. Can. J. Remote Sens. 2003, 29, 623–632. [Google Scholar] [CrossRef]
- Nasset, E. Effects of different flying altitudes on biophysical stand properties estimated from canopy height and density measured with a small-footprint airborne scanning laser. Remote Sens. Environ. 2004, 91, 243–255. [Google Scholar] [CrossRef]
- Zhao, K.; Suarez, J.C.; Garcia, M.; Hu, T.; Wang, C.; Londo, A. Utility of multitemporal lidar for forest and carbon monitoring: Tree growth, biomass dynamics, and carbon flux. Remote Sens. Environ. 2018, 204, 883–897. [Google Scholar] [CrossRef]
- Duncanson, L.; Dubayah, R. Monitoring individual tree-based change with airborne lidar. Ecol. Evol. 2018, 8, 5079–5089. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Skidmore, A.K.; Jones, S.; Wang, T.; Heurich, M.; Zhu, X.; Shi, Y. Large off-nadir scan angle of airborne LiDAR can severely affect the estimates of forest structure metrics. ISPRS J. Photogramm. Remote Sens. 2018, 136, 13–25. [Google Scholar] [CrossRef]
- Michez, A.; Huylenbroeck, L.; Bolyn, C.; Latte, N.; Bauwens, S.; Lejeune, P. Can regional aerial images from orthophoto surveys produce high quality photogrammetric Canopy Height Model? A single tree approach in Western Europe. Int. J. Appl. Earth Obs. Geoinf. 2020, 92, 102190. [Google Scholar] [CrossRef]
- Giese, L.A.B.; Aust, W.M.; Kolka, R.K.; Trettin, C.C. Biomass and carbon pools of disturbed riparian forests. For. Ecol. Manag. 2003, 180, 493–508. [Google Scholar] [CrossRef]
- Cierjacks, A.; Kleinschmit, B.; Babinsky, M.; Kleinschroth, F.; Markert, A.; Menzel, M.; Ziechmann, U.; Schiller, T.; Graf, M.; Lang, F. Carbon stocks of soil and vegetation on Danubian floodplains. J. Plant Nutr. Soil Sci. 2010, 173, 644–653. [Google Scholar] [CrossRef]
- Latte, N.; Colinet, G.; Fayolle, A.; Lejeune, P.; Hébert, J.; Claessens, H.; Bauwens, S. Description of a new procedure to estimate the carbon stocks of all forest pools and impact assessment of methodological choices on the estimates. Eur. J. For. Res. 2013, 132, 565–577. [Google Scholar] [CrossRef] [Green Version]
- Dufour, S.; Rodríguez-González, P.M.; Laslier, M. Tracing the scientific trajectory of riparian vegetation studies: Main topics, approaches and needs in a globally changing world. Sci. Total Environ. 2019, 653, 1168–1185. [Google Scholar] [CrossRef]
- Strnadová, V.; Černý, K.; Holub, V.; Gregorová, B. The effects of flooding and Phytophthora alni infection on black alder. J. For. Sci. 2010, 56, 6. [Google Scholar] [CrossRef] [Green Version]
- Marçais, B.; Husson, C.; Godart, L.; Caël, O. Influence of site and stand factors on Hymenoscyphus fraxineus-induced basal lesions. Plant Pathol. 2016, 65, 1452–1461. [Google Scholar] [CrossRef]
- Ferry, B.; Morneau, F.; Bontemps, J.-D.; Blanc, L.; Freycon, V. Higher treefall rates on slopes and waterlogged soils result in lower stand biomass and productivity in a tropical rain forest. J. Ecol. 2010, 98, 106–116. [Google Scholar] [CrossRef]
- Cavalcanti, G.G.; Lockaby, B.G. Effects of sediment deposition on aboveground net primary productivity, vegetation composition, and structure in riparian forests. Wetlands 2006, 26, 400–409. [Google Scholar] [CrossRef]
- Jolley, R.L.; Lockaby, B.G.; Cavalcanti, G.G. Productivity of Ephemeral Headwater Riparian Forests Impacted by Sedimentation in the Southeastern United States Coastal Plain. J. Environ. Qual. 2009, 38, 965–979. [Google Scholar] [CrossRef] [PubMed]
- Clawson, R.G.; Lockaby, B.G.; Rummer, B. Changes in Production and Nutrient Cycling across a Wetness Gradient within a Floodplain Forest. Ecosystems 2001, 4, 126–138. [Google Scholar] [CrossRef]
- Amlin, N.M.; Rood, S.B. Drought stress and recovery of riparian cottonwoods due to water table alteration along Willow Creek, Alberta. Trees 2003, 17, 351–358. [Google Scholar] [CrossRef]
- Odum, E.P.; Finn, J.T.; Franz, E.H. Perturbation Theory and the Subsidy-Stress Gradient. BioScience 1979, 29, 349–352. [Google Scholar] [CrossRef]
- Schilling, E.B.; Lockaby, B.G. Relationships between productivity and nutrient circulation within two contrasting southeastern U.S. floodplain forests. Wetlands 2006, 26, 181–192. [Google Scholar] [CrossRef]
- Francalanci, S.; Paris, E.; Solari, L. On the vulnerability of woody riparian vegetation during flood events. Environ. Fluid Mech. 2019. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Variable | Definition | Interest for Biomass Prediction |
|---|---|---|---|
| CHM | H90 (m) | 90th height percentile within the canopy | Tree size |
| Area (m2) | Tree crown area (digitized or automatically segmented) | Tree size | |
| Point cloud (std.metrics) | Zq30 (m) | 30th height percentile within the canopy | Crown shape: trees located inside forests have more branches at the top of the crown |
| Pground (%) | Proportion of returns classified as “ground” | Crown porosity: heliophilous species have less dense branching |
| Thematic Group | Scale | Name | Detail | Source |
|---|---|---|---|---|
| History | Vegetation unit (0.3 ha) | Age (years) | Age estimated by photo-interpretation of historical aerial images | Historical orthophotos |
| Planted | Regeneration type (1 = planting, 0 = natural regeneration or undescribed). Described only for VUs less than 40 years old | |||
| Geomorphology | Vegetation unit (0.3 ha) | Horizontal distance (m) | Horizontal distance to the main channel | Hydrographic Network |
| Vertical distance (m) | Vertical distance to river mean water level. | LiDAR digital terrain model (DTM) | ||
| Relative vertical distance | Vertical distance to river mean water level, divided by the relative altitude of the 25-year flood stage. A value less than 1 means that the VU is located in the 25-year floodplain, while a value superior to 1 corresponds to valley slopes. Values higher than 2 were thresholded at 2. | |||
| Slope (%) | Terrain slope | |||
| Waterlogging | Waterlogging classes. Anoxia traces are found beyond 125 cm deep (class 1), between 80 and 125 cm (class 2), between 80 and 50 cm (class 3), between 30 et 50 cm (class 4), before 30 cm deep (class 5). | Regional soil map [59] | ||
| Floodplain (250 m upstream and downstream of VU) | Width (m) | Floodplain width (25-year flood) | 25-year flood map [45] | |
| Sinuosity | River sinuosity | Hydrographic network | ||
| Catchment area (m2) | Catchment area | LiDAR digital terrain model (DTM) | ||
| Land use | Vegetation unit (0.3 ha) | Artificial in VU (%) | % artificial areas | Regional land use map [60] |
| Agriculture in VU (%) | % agricultural areas | |||
| Forest in VU (%) | % forest and other natural areas | |||
| Floodplain (250 m upstream and downstream of VU) | Artificial in FP (%) | % artificial areas | ||
| Agriculture in FP (%) | % agricultural areas | |||
| Forest in FP (%) | % forest and other natural areas |
| Model | R2 | Mean Error | MAE |
|---|---|---|---|
| m1 | 0.79 | 0.0029 (1.0029) | 0.4644 a (1.5911) |
| m2 | 0.81 | 0.0022 (1.0022) | 0.4513 b (1.5703) |
| Model | R2 | Mean Error (Mg/ha) | MAE (Mg/ha) | RMSE (Mg/ha) | RMSEr |
|---|---|---|---|---|---|
| m1 | 0.83 | 1.75 a (n.s.) | 12.79 a | 19.44 a | 0.27 |
| m2 | 0.90 | 1.70 a (n.s.) | 11.26 a | 15.52 a | 0.22 |
| Term | Estimate | Std.Error | Statistic | p-Value | Relative Importance (%) | Relative Importance (Rank) |
|---|---|---|---|---|---|---|
| (Intercept) | 113.51 | 1.18 | 95.86 | 0 | ||
| Age | 39.09 | 1.25 | 31.16 | 5.33 × 10−186 | 25.17 | 1 |
| Agriculture in FP | −15.33 | 1.37 | −11.22 | 1.12 × 10−28 | 9.53 | 2 |
| Vertical distance | 7.99 | 1.28 | 6.27 | 4.20 × 10−10 | 4.8 | 3 |
| Forest in VU | 10.6 | 1.41 | 7.54 | 6.18 × 10−14 | 4.26 | 4 |
| Age:Forest in VU | 10.93 | 1.26 | 8.69 | 5.67 × 10−18 | 2.12 | 5 |
| Horizontal distance | −5.19 | 1.13 | −4.57 | 4.98 × 10−6 | 1.04 | 6 |
| Planted | 25.78 | 4.66 | 5.53 | 3.38 × 10−8 | 0.94 | 7 |
| Age:Agriculture in FP | −5.84 | 1.27 | −4.58 | 4.73 × 10−6 | 0.96 | 8 |
| Artificial in VU | −5.55 | 1.14 | −4.85 | 1.29 × 10−6 | 0.83 | 9 |
| Age:Planted | 15.57 | 4.31 | 3.62 | 3.04 × 10−4 | 0.23 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huylenbroeck, L.; Latte, N.; Lejeune, P.; Georges, B.; Claessens, H.; Michez, A. What Factors Shape Spatial Distribution of Biomass in Riparian Forests? Insights from a LiDAR Survey over a Large Area. Forests 2021, 12, 371. https://doi.org/10.3390/f12030371
Huylenbroeck L, Latte N, Lejeune P, Georges B, Claessens H, Michez A. What Factors Shape Spatial Distribution of Biomass in Riparian Forests? Insights from a LiDAR Survey over a Large Area. Forests. 2021; 12(3):371. https://doi.org/10.3390/f12030371
Chicago/Turabian StyleHuylenbroeck, Leo, Nicolas Latte, Philippe Lejeune, Blandine Georges, Hugues Claessens, and Adrien Michez. 2021. "What Factors Shape Spatial Distribution of Biomass in Riparian Forests? Insights from a LiDAR Survey over a Large Area" Forests 12, no. 3: 371. https://doi.org/10.3390/f12030371
APA StyleHuylenbroeck, L., Latte, N., Lejeune, P., Georges, B., Claessens, H., & Michez, A. (2021). What Factors Shape Spatial Distribution of Biomass in Riparian Forests? Insights from a LiDAR Survey over a Large Area. Forests, 12(3), 371. https://doi.org/10.3390/f12030371