
Seemingly Unrelated Mixed-Effects Biomass Models for Black Locust in West Poland


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Material Collection and Preparation
2.3. Individual Mixed-Effect Models
2.4. Seemingly Unrelated Mixed-Effects Model System
2.5. Fixed- and Random-Effects Prediction
3. Results
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huntley, J.C. Robinia pseudoacacia L. black locust. In Silvic of North America 2. Hardwoods, Agriculture Handbook; Burns, R.M., Honkala, B.H., Eds.; United States Department of Agriculture (USDA), Forest Service: Washington, DC, USA, 1990; pp. 755–761. [Google Scholar]
- Nowiński, M. Dzieje Roślin i Upraw Ogrodniczych (History of Plants and Horticultural Crops); PWRiL: Warsaw, Poland, 1977. (In Polish) [Google Scholar]
- Vítková, M.; Müllerová, J.; Sádlo, J.; Pergl, J.; Pyšek, P. Black Locust (Robinia Pseudoacacia) Beloved and Despised: A Story of an Invasive Tree in Central Europe. For. Ecol. Manag. 2017, 384, 287–302. [Google Scholar] [CrossRef]
- Sitzia, T.; Cierjacks, A.; de Rigo, D.; Caudullo, G. Robinia pseudoacacia in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Durrant, T.H., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; pp. 66–167. [Google Scholar]
- Kraszkiewicz, A. Evaluation of the Possibility of Energy Use Black Locust (Robinia Pseudoacacia L.) Dendromass Acquired in Forest Stands Growing on Clay Soils. J. Cent. Eur. Agric. 2013, 14, 388–399. [Google Scholar] [CrossRef]
- Bolat, I.; Kara, Ö.; Sensoy, H.; Yüksel, K. Influences of Black Locust (Robinia Pseudoacacia L.) Afforestation on Soil Microbial Biomass and Activity. Iforest Biogeosci. For. 2015, 9, 171–177. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Wierzcholska, S.; Dyderski, M.K.; Horodecki, P.; Rusińska, A.; Gdula, A.K.; Kasprowicz, M. Tree Species Effects on Bryophyte Guilds on a Reclaimed Post-Mining Site. Ecol. Eng. 2018, 110, 117–127. [Google Scholar] [CrossRef]
- Horodecki, P.; Nowiński, M.; Jagodziński, A.M. Advantages of Mixed Tree Stands in Restoration of Upper Soil Layers on Postmining Sites: A Five-Year Leaf Litter Decomposition Experiment. Land Degrad. Dev. 2019, 30, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Qiu, L.; Zhang, X.; Cheng, J.; Yin, X. Effects of Black Locust (Robinia Pseudoacacia) on Soil Properties in the Loessial Gully Region of the Loess Plateau, China. Plant Soil 2010, 332, 207–217. [Google Scholar] [CrossRef]
- Rice, S.K.; Westerman, B.; Federici, R. Impacts of the Exotic, Nitrogen-Fixing Black Locust (Robinia Pseudoacacia) on Nitrogen-Cycling in a Pine-Oak Ecosystem. Plant Ecol. 2004, 174, 97–107. [Google Scholar] [CrossRef]
- Li, G.; Xu, G.; Guo, K.; Du, S. Mapping the Global Potential Geographical Distribution of Black Locust (Robinia Pseudoacacia L.) Using Herbarium Data and a Maximum Entropy Model. Forests 2014, 5, 2773–2792. [Google Scholar] [CrossRef] [Green Version]
- Rédei, K.; Csiha, I.; Keseru, Z.; Rásó, J.; Végh, Á.K.; Antal, B. Growth and Yield of Black Locust (Robinia Pseudoacacia L.) Stands in Nyírség Growing Region (North-East Hungary). South East Eur. For. 2014, 5, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Nadal-Sala, D.; Hartig, F.; Gracia, C.A.; Sabaté, S. Global Warming Likely to Enhance Black Locust (Robinia Pseudoacacia L.) Growth in a Mediterranean Riparian Forest. For. Ecol. Manag. 2019, 449, 117448. [Google Scholar] [CrossRef]
- Puchałka, R.; Dyderski, M.K.; Vítková, M.; Sádlo, J.; Klisz, M.; Netsvetov, M.; Prokopuk, Y.; Matisons, R.; Mionskowski, M.; Wojda, T.; et al. Black Locust (Robinia Pseudoacacia L.) Range Contraction and Expansion in Europe under Changing Climate. Glob. Chang. Biol. 2021, 27, 1587–1600. [Google Scholar] [CrossRef]
- Tokarska-Guzik, B. The Establishment and Spread of Alien Plant Species (Kenophytes) in the Flora of Poland; Wydawnictwo Uniwersytetu Śląskiego: Katowice, Poland, 2005. [Google Scholar]
- Bronisz, K.; Zasada, M. Taper Models for Black Locust in West Poland. Silva Fenn. 2020, 54. [Google Scholar] [CrossRef]
- Wojda, T.; Klisz, M.; Jastrzębowski, S.; Mionskowski, M.; Szyp-Borowska, I.; Szczygieł, K. The Geographical Distribution Of The Black Locust (Robinia Pseudoacacia L.) In Poland And Its Role On Non-Forest Land. Pap. Glob. Chang. 2015, 22, 101–113. [Google Scholar] [CrossRef]
- Clark, D.A.; Brown, S.; Kicklighter, D.W.; Chambers, J.Q.; Thomlinson, J.R.; Ni, J. Measuring Net Primary Production in Forests: Concepts and Field Methods. Ecol. Appl. 2001, 11, 356–370. [Google Scholar] [CrossRef]
- IPCC Guidelines for National Greenhouse Gas Inventories: Reference Manual; UK Meteorological Office: Bracknell, UK, 1996.
- Cieszewski, C.J.; Zasada, M.; Lowe, R.C.; Liu, S. Estimating Biomass and Carbon Storage by Georgia Forest Types and Species Groups Using the FIA Data Diameters, Basal Areas, Site Indices, and Total Heights. Forests 2021, 12, 141. [Google Scholar] [CrossRef]
- Zianis, D.; Muukkonen, P.; Mäkipää, R.; Mencuccini, M. Biomass and Stem Volume Equations for Tree Species in Europe; Silva Fennica Monographs; Finnish Society of Forest Science, Finnish Forest Research Institute: Tampere, Finland, 2005. [Google Scholar]
- Zianis, D.; Mencuccini, M. Aboveground Biomass Relationships for Beech (Fagus moesiaca Cz.) Trees in Vermio Mountain, Northern Greece, and Generalised Equations for Fagus Sp. Ann. For. Sci. 2003, 60, 439–448. [Google Scholar] [CrossRef] [Green Version]
- Lehtonen, A.; Mäkipää, R.; Heikkinen, J.; Sievänen, R.; Liski, J. Biomass Expansion Factors (BEFs) for Scots Pine, Norway Spruce and Birch According to Stand Age for Boreal Forests. For. Ecol. Manag. 2004, 188, 211–224. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D. Mixed-Effects Models in S and S-PLUS; Springer: New York, NY, USA, 2013. [Google Scholar]
- Mehtätalo, L.; Lappi, J. Forest Biometrics with Examples in R; Chapman & Hall/CRC: New York, NY, USA, 2020. [Google Scholar]
- Fehrmann, L.; Lehtonen, A.; Kleinn, C.; Tomppo, E. Comparison of Linear and Mixed-Effect Regression Models and a k-Nearest Neighbour Approach for Estimation of Single-Tree Biomass. Can. J. For. Res. 2008, 38, 1–9. [Google Scholar] [CrossRef]
- Bronisz, K.; Mehtätalo, L. Seemingly Unrelated Mixed-Effects Biomass Models for Young Silver Birch Stands on Post-Agricultural Lands. Forests 2020, 11, 381. [Google Scholar] [CrossRef] [Green Version]
- Lappi, J. Calibration of Height and Volume Equations with Random Parameters. For. Sci. 1991, 37, 781–801. [Google Scholar]
- Bronisz, K.; Mehtätalo, L. Mixed-Effects Generalized Height–Diameter Model for Young Silver Birch Stands on Post-Agricultural Lands. For. Ecol. Manag. 2020, 460, 117901. [Google Scholar] [CrossRef]
- Ou, G.; Wang, J.; Xu, H.; Chen, K.; Zheng, H.; Zhang, B.; Sun, X.; Xu, T.; Xiao, Y. Incorporating Topographic Factors in Nonlinear Mixed-Effects Models for Aboveground Biomass of Natural Simao Pine in Yunnan, China. J. For. Res. 2016, 27, 119–131. [Google Scholar] [CrossRef]
- Pearce, H.G.; Anderson, W.R.; Fogarty, L.G.; Todoroki, C.L.; Anderson, S.A.J. Linear Mixed-Effects Models for Estimating Biomass and Fuel Loads in Shrublands. Can. J. For. Res. 2010, 40, 2015–2026. [Google Scholar] [CrossRef]
- Kozak, A. Methods for Ensuring Additivity of Biomass Components by Regression Analysis. For. Chron. 1970, 46, 402–405. [Google Scholar] [CrossRef]
- Parresol, B.R. Assessing Tree and Stand Biomass: A Review with Examples and Critical Comparisons. For. Sci. 1999, 45, 573–593. [Google Scholar]
- Vonderach, C.; Kändler, G.; Dormann, C.F. Consistent Set of Additive Biomass Functions for Eight Tree Species in Germany Fit by Nonlinear Seemingly Unrelated Regression. Ann. For. Sci. 2018, 75, 49. [Google Scholar] [CrossRef] [Green Version]
- Ung, C.-H.; Bernier, P.; Guo, X.-J. Canadian National Biomass Equations: New Parameter Estimates That Include British Columbia Data. Can. J. For. Res. 2008, 38, 1123–1132. [Google Scholar] [CrossRef]
- Bronisz, K.; Strub, M.; Cieszewski, C.; Bijak, S.; Bronisz, A.; Tomusiak, R.; Wojtan, R.; Zasada, M. Empirical Equations for Estimating Aboveground Biomass of Betula Pendula Growing on Former Farmland in Central Poland. Silva Fenn. 2016, 50. [Google Scholar] [CrossRef] [Green Version]
- Martyn, D. Klimaty Kuli Ziemskiej (Climates of the Earth); PWN: Warsaw, Poland, 2000. (In Polish) [Google Scholar]
- European Climate Assessment & Dataset. Available online: https://www.ecad.eu (accessed on 5 February 2021).
- Bronisz, K.; Bijak, S.; Wojtan, R.; Tomusiak, R.; Bronisz, A.; Baran, P.; Zasada, M. Biomass_Black_Locuts.xlsx. 2021. Available online: https://figshare.com/articles/dataset/Biomass_Black_Locuts_xlsx/13721107/1 (accessed on 23 March 2021). [CrossRef]
- Samuelsson, R.; Burvall, J.; Jirjis, R. Comparison of Different Methods for the Determination of Moisture Content in Biomass. Biomass Bioenergy 2006, 30, 929–934. [Google Scholar] [CrossRef]
- Snowdon, P.; Raison, J.; Keith, H.; Ritson, P.; Grierson, P.; Adams, M.A.; Montagu, K.; Hui-quan, B.; Burrows, W.; Eamus, D. Protocol for Sampling Tree and Stand Biomass; Australian Greenhouse Office: Canberra, Australia, 2002. [Google Scholar]
- Baskerville, G.L. Use of Logarithmic Regression in the Estimation Of plant Biomass. Can. J. For. Res. 1972, 2, 49–53. [Google Scholar] [CrossRef]
- Pukelsheim, F. Optimal Design of Experiments; Classics in Applied Mathematics; Society for Industrial and Applied Mathematics, University City: Philadelphia, PA, USA, 2006. [Google Scholar]
- Pinheiro, J.; Bates, D.; Saikat, D.; Deepayan, S. Nlme: Linear and Nonlinear Mixed Effects Models; 2020; Available online: https://cran.r-project.org/web/packages/nlme/index.html (accessed on 23 March 2021).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R; RStudio Inc.: Boston, MA, USA, 2015. [Google Scholar]
- Cunia, T.; Briggs, R.D. Forcing Additivity of Biomass Tables—Some Empirical Results. Can. J. For. Res. 1984, 14, 376–384. [Google Scholar] [CrossRef]
- Chiyenda, S.S.; Kozak, A. Additivity of Component Biomass Regression Equations When the Underlying Model Is Linear. Can. J. For. Res. 1984, 14, 441–446. [Google Scholar] [CrossRef]
- Zasada, M.; Bronisz, K.; Bijak, S.; Wojtan, R.; Tomusiak, R.; Dudek, A.; Michalak, K.; Wróblewski, L. Wzory Empiryczne Do Określania Suchej Biomasy Nadziemnej Części Drzew i Ich Komponentów (Empirical Formulae for Determination of the Dry Biomass of Aboveground Parts of the Tree). Sylwan 2008, 152, 27–39. (In Polish) [Google Scholar] [CrossRef]
- Bronisz, K.; Zasada, M. Uproszczone Wzory Empiryczne Do Określania Suchej Biomasy Nadziemnej Części Drzew i Ich Komponentów Dla Sosny Zwyczajnej (Simplified Empirical Formulas to Determine the Dry Biomass of Aboveground Components of Trees for Scots Pine). Sylwan 2016, 160, 277–283. (In Polish) [Google Scholar] [CrossRef]
- Vauhkonen, J.; MehtÄtalo, L.; Packalén, P. Combining Tree Height Samples Produced by Airborne Laser Scanning and Stand Management Records to Estimate Plot Volume in Eucalyptus Plantations. Can. J. For. Res. 2011, 41, 1649–1658. [Google Scholar] [CrossRef]
- Maltamo, M.; Mehtätalo, L.; Vauhkonen, J.; Packalén, P. Predicting and Calibrating Tree Attributes by Means of Airborne Laser Scanning and Field Measurements. Can. J. For. Res. 2012, 42, 1896–1907. [Google Scholar] [CrossRef]
- Stereńczak, K.; Zasada, M. Accuracy of Tree Height Estimation Based on LIDAR Data Analysis. Folia For. Pol. 2011, 53, 123–129. [Google Scholar]
- Repola, J. Biomass Equations for Birch in Finland. Silva Fenn. 2008, 42, 605–624. [Google Scholar] [CrossRef] [Green Version]





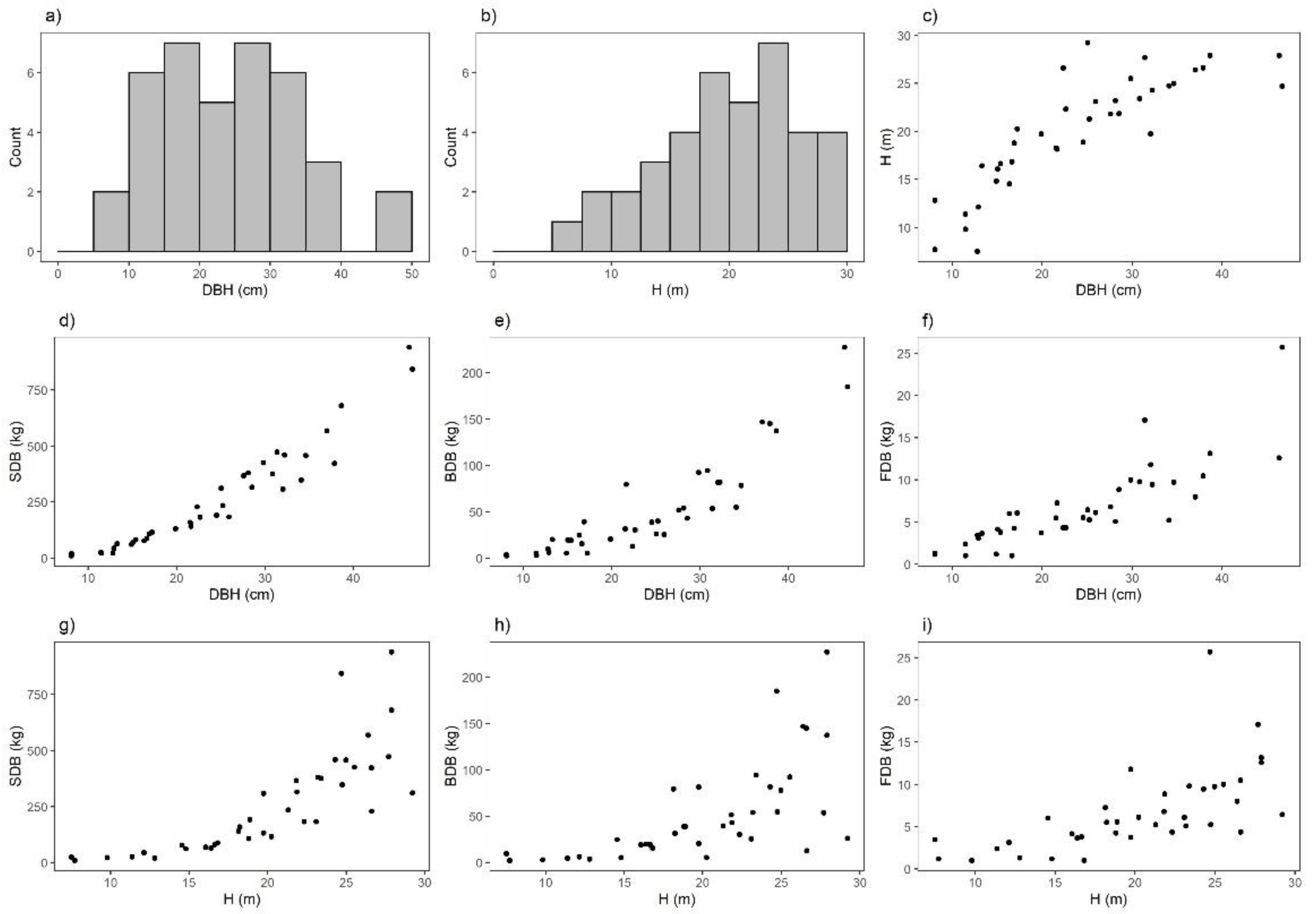{kind=link}
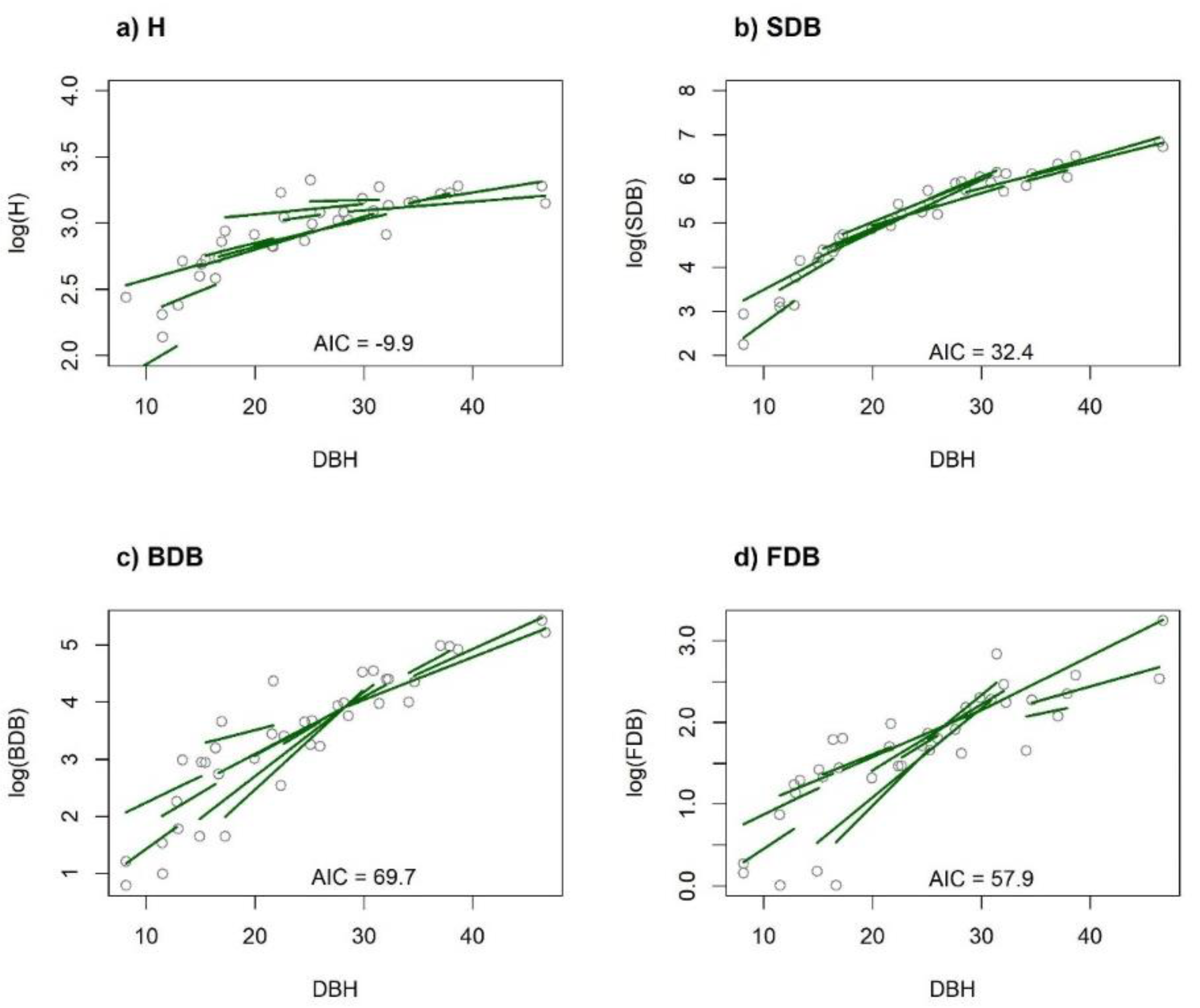{kind=link}
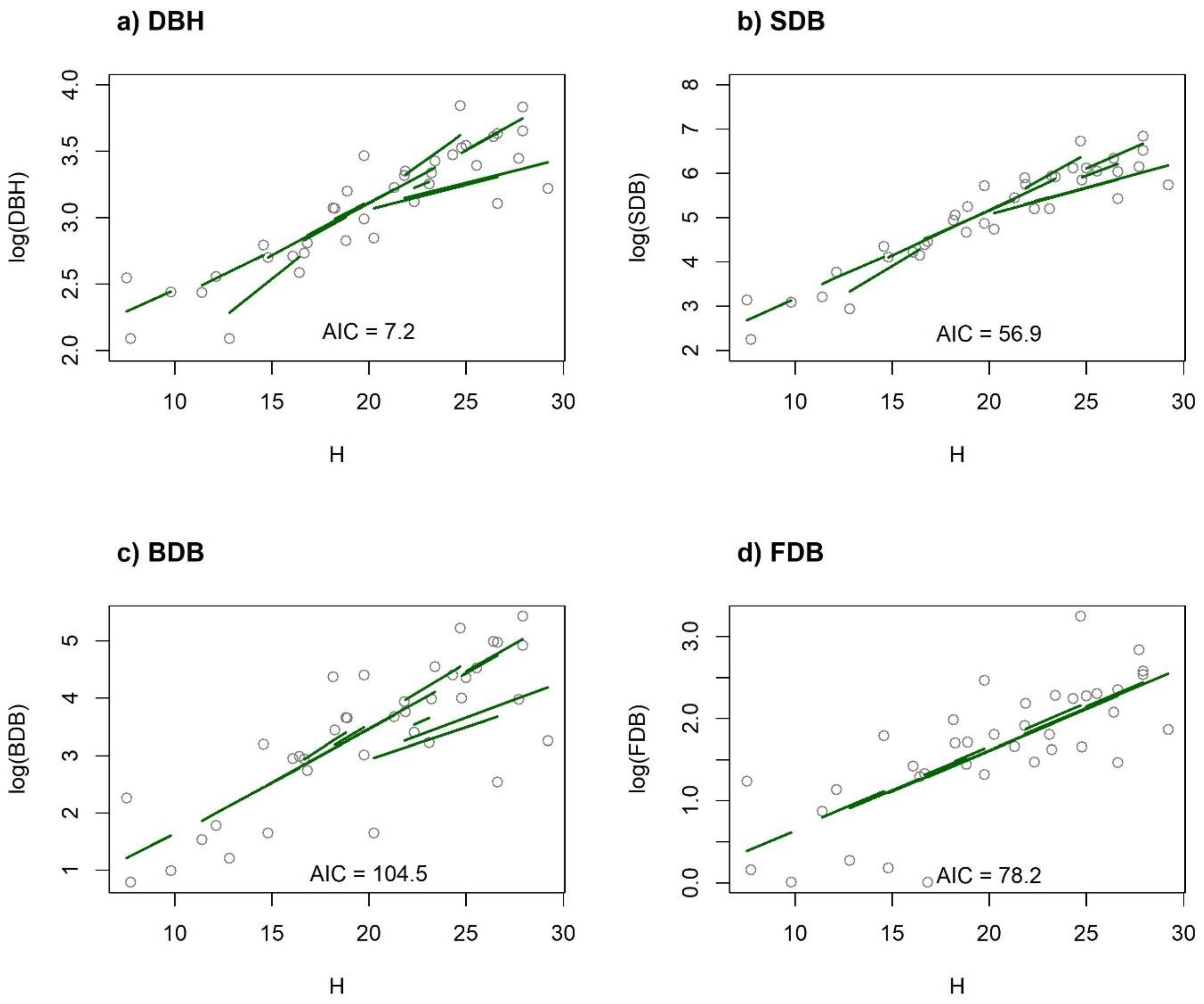{kind=link}
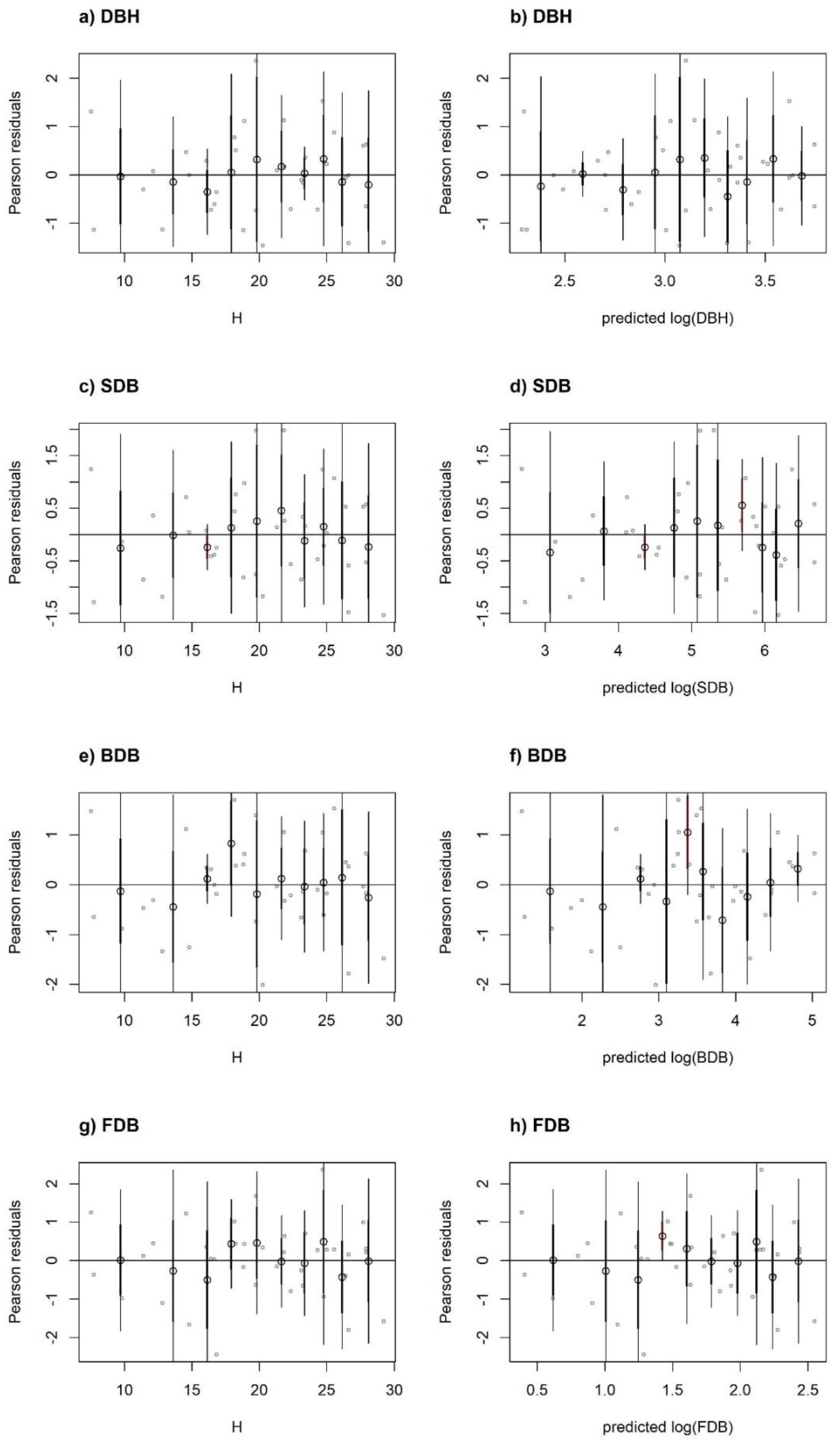{kind=link}
| Minimum | Maximum | Mean | Median | Standard Deviation | |
|---|---|---|---|---|---|
| A | 16 | 85 | 50 | 50 | 21 |
| N | 167 | 1134 | 586 | 507 | 304 |
| BA | 10.06 | 40.52 | 21.73 | 20.53 | 7.72 |
| Dg | 11.47 | 43.94 | 23.94 | 25.23 | 9.08 |
| Hg | 9.84 | 27.19 | 20.30 | 21.18 | 5.19 |
| Minimum | Maximum | Mean | Median | Standard Deviation | |
|---|---|---|---|---|---|
| DBH | 8.1 | 46.7 | 24.1 | 23.6 | 10.13 |
| H | 7.50 | 29.22 | 20.11 | 20.77 | 5.87 |
| SDB | 10 | 940 | 261 | 187 | 230 |
| BDB | 2 | 227 | 53 | 35 | 54 |
| FDB | 1 | 26 | 7 | 6 | 5 |
| First Approach | ||||
| Dependent variable | SDB | BDB | FDB | H |
| 2.868 (0.319) | 0.318 (0.434) | −0.371 (0.325) | 2.593 (0.149) | |
| 0.106 (0.014) | 0.139 (0.019) | 0.092 (0.013) | 0.019 (0.005) | |
| −0.25 | −0.812 | −0.629 | −0.384 | |
| 0.3672 | 4.3812 | 1.9972 | 0.332 | |
| Second Approach | ||||
| Dependent variable | SDB | BDB | FDB | DBH |
| 1.401 (0.322) | −0.908 (0.448) | −0.623 (0.319) | 1.526 (0.186) | |
| 0.189 (0.016) | 0.217 (0.023) | 0.113 (0.015) | 0.079 (0.009) | |
| −0.179 | −0.095 | −0.332 | −0.235 | |
| 0.532 | 0.8652 | 1.3362 | 0.312 | |
| First Approach | |||||||||
| Dependent variable | SDB | BDB | FDB | H | |||||
| SDB | 1.0952 | −0.989 | 0.605 | −0.849 | 0.419 | −0.652 | 0.978 | −0.981 | |
| - | 0.0482 | −0.603 | 0.856 | −0.357 | 0.620 | −0.943 | 0.968 | ||
| BDB | - | - | 1.3942 | −0.920 | −0.027 | −0.024 | 0.677 | −0.748 | |
| - | - | - | 0.0662 | −0.065 | 0.243 | −0.873 | 0.931 | ||
| FDB | - | - | - | - | 1.0252 | −0.926 | 0.417 | −0.346 | |
| - | - | - | - | - | 0.0412 | −0.597 | 0.549 | ||
| H | - | - | - | - | - | - | 0.5022 | −0.984 | |
| - | - | - | - | - | - | - | 0.0172 | ||
| Second Approach | |||||||||
| Dependent variable | SDB | BDB | FDB | DBH | |||||
| SDB | 0.8362 | −0.975 | 0.530 | −0.982 | −0.493 | −0.693 | 0.873 | −0.952 | |
| - | 0.0412 | −0.330 | 0.917 | 0.673 | 0.517 | −0.744 | 0.862 | ||
| BDB | - | - | 0.4572 | −0.679 | 0.475 | −0.978 | 0.876 | −0.763 | |
| - | - | - | 0.0332 | 0.323 | 0.815 | −0.948 | 0.992 | ||
| FDB | - | - | - | - | 0.1292 | −0.285 | −0.007 | 0.205 | |
| - | - | - | - | - | 02 | −0.957 | 0.880 | ||
| DBH | - | - | - | - | - | - | 0.5132 | −0.980 | |
| - | - | - | - | - | - | - | 0.0242 | ||
| First Approach | |||
| Dependent variable | SDB | BDB | FDB |
| BDB | 0.257 | - | - |
| FDB | −0.100 | 0.557 | - |
| H | 0.474 | −0.300 | −0.287 |
| Second Approach | |||
| Dependent variable | SDB | BDB | FDB |
| BDB | 0.889 | - | - |
| FDB | 0.701 | 0.712 | - |
| DBH | 0.933 | 0.844 | 0.766 |
| First Approach | ||||
| Dependent variable | SDB | BDB | FDB | H |
| Fixed-effects prediction | 238.716 | 95.157 | 9.313 | 5.752 |
| Random-effects prediction | 77.603 | 36.018 | 6.887 | 3.297 |
| Second Approach | ||||
| Dependent variable | SDB | BDB | FDB | DBH |
| Fixed-effects prediction | 206.933 | 67.366 | 3.726 | 8.868 |
| Random-effects prediction | 188.139 | 70.629 | 3.507 | 8.491 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bronisz, K.; Bijak, S.; Wojtan, R.; Tomusiak, R.; Bronisz, A.; Baran, P.; Zasada, M. Seemingly Unrelated Mixed-Effects Biomass Models for Black Locust in West Poland. Forests 2021, 12, 380. https://doi.org/10.3390/f12030380
Bronisz K, Bijak S, Wojtan R, Tomusiak R, Bronisz A, Baran P, Zasada M. Seemingly Unrelated Mixed-Effects Biomass Models for Black Locust in West Poland. Forests. 2021; 12(3):380. https://doi.org/10.3390/f12030380
Chicago/Turabian StyleBronisz, Karol, Szymon Bijak, Rafał Wojtan, Robert Tomusiak, Agnieszka Bronisz, Paweł Baran, and Michał Zasada. 2021. "Seemingly Unrelated Mixed-Effects Biomass Models for Black Locust in West Poland" Forests 12, no. 3: 380. https://doi.org/10.3390/f12030380
APA StyleBronisz, K., Bijak, S., Wojtan, R., Tomusiak, R., Bronisz, A., Baran, P., & Zasada, M. (2021). Seemingly Unrelated Mixed-Effects Biomass Models for Black Locust in West Poland. Forests, 12(3), 380. https://doi.org/10.3390/f12030380