
Productivity of Black Locust (Robinia pseudoacacia L.) Grown on a Varying Habitats in Southeastern Poland


Abstract
:1. Introduction
2. Materials and Methods
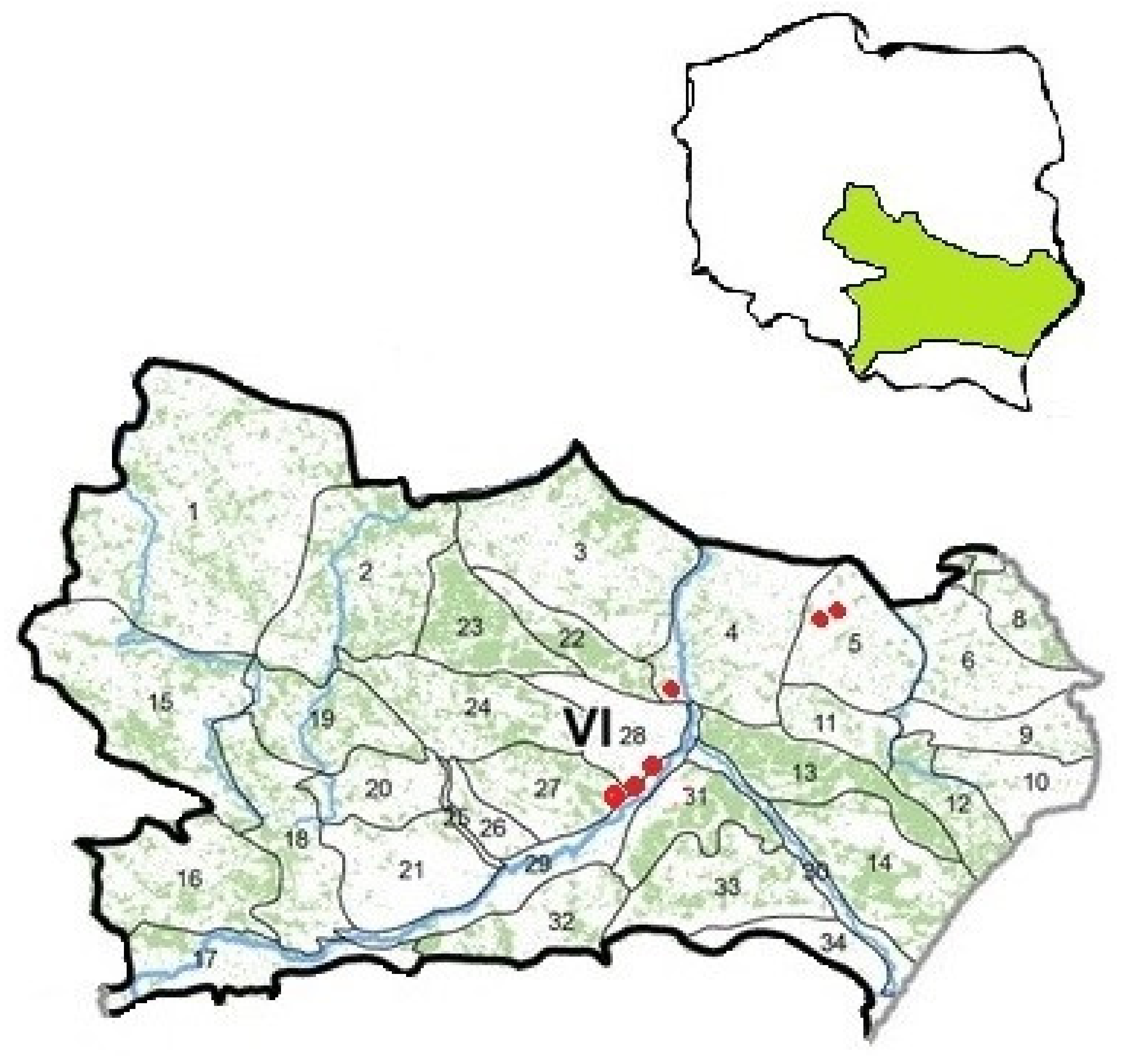
2.1. Study Sites
- ⮚
- five on sands (loose sands)—No. 1–5—All in the Piaseczno site, derived from planting, at the age of 35, on the slopes of the dump with various exposures (N—Surfaces No. 1 and 2 located in the upper and lower parts of the slope, respectively, SE—No. 3 and 4—The upper and lower part of the slope, S—No. 5—The upper part of the slope), without felling (without harvesting);
- ⮚
- three in the clay—No. 6–8—two in the Piaseczno site, derived from planting, at the age of 35, (located in the upper parts of the slopes, No. 6—N exposure, and No. 7—S exposure) under development conditions such as stands No. 1–5, and one in the Zawidza site (No. 8) in the managed forest (with intermediate cutting—Breeding cuts), at the age of 41, in the plain;
- ⮚
- six on dust formations (loess and loess-like)—In the sites: Dębno (No. 9, the lower part of the valley slope, with S exposure and 40% slope)—33-year-old stand from self-seeding, systematically cut; Skrzypaczowice (No. 10)—A 64-year-old commercial stand, planted, located in the central part of the eroded slope with an SE exposure and a 15% slope; Lublin (No. 11)—4-year self-seeding trees located on a plain area; Snopków (No. 12–14)—3-row mid-field, 8-year-old plantings, No. 12 and 13 located in a flat area, and No. 14 in the upper part of the valley slope, with S exposure and a 15% slope.
2.2. Biomass Sampling
- (a).
- Stand parameters—By the method of sample plots of 500 m2 (20 × 25 m2) in stands No. 1–11 and 400 m2 in stands No. 12–14 (rows of trees 80 m long and 5 m wide):
- ⮚
- the number and social structure of trees according to the tree biological classification of Kraft, considering the main and secondary stands;
- ⮚
- average height of trees (in main and secondary stands)—as the arithmetic mean of DBH measured with the SUUNTO altimeter with an accuracy of 0.25 m;
- ⮚
- mean DBH (in main and secondary stands)—as the arithmetic mean of DBH measured with a precision HAGLOF caliper with an accuracy of 0.5 cm;
- (b).
- Volume with division into trunks and branches and wood thickness classes (0.0–1.0 cm, 1.1–5.0 cm, 5.1–10.0 cm and then every 5 cm)—using the sample tree’s method. One tree of average height and DBH each and average conformation from each sample plot (main and secondary stands) [47]; a conductor was considered as a trunk from the point of cut (5–10 cm above the ground) to a diameter of 5 cm in the bark (in the upper end); the remaining, thinner part (top) was classified as a branch; the sample trees were cut with a chainsaw at the end of December:
- ⮚
- trunk volume in the bark—sectional method (section length 1 m);
- ⮚
- branch volume in the bark—using the xylometric method.
2.3. Statistical Analysis
3. Results
3.1. Characteristics of Soils
3.2. Characteristics of Forest Stands
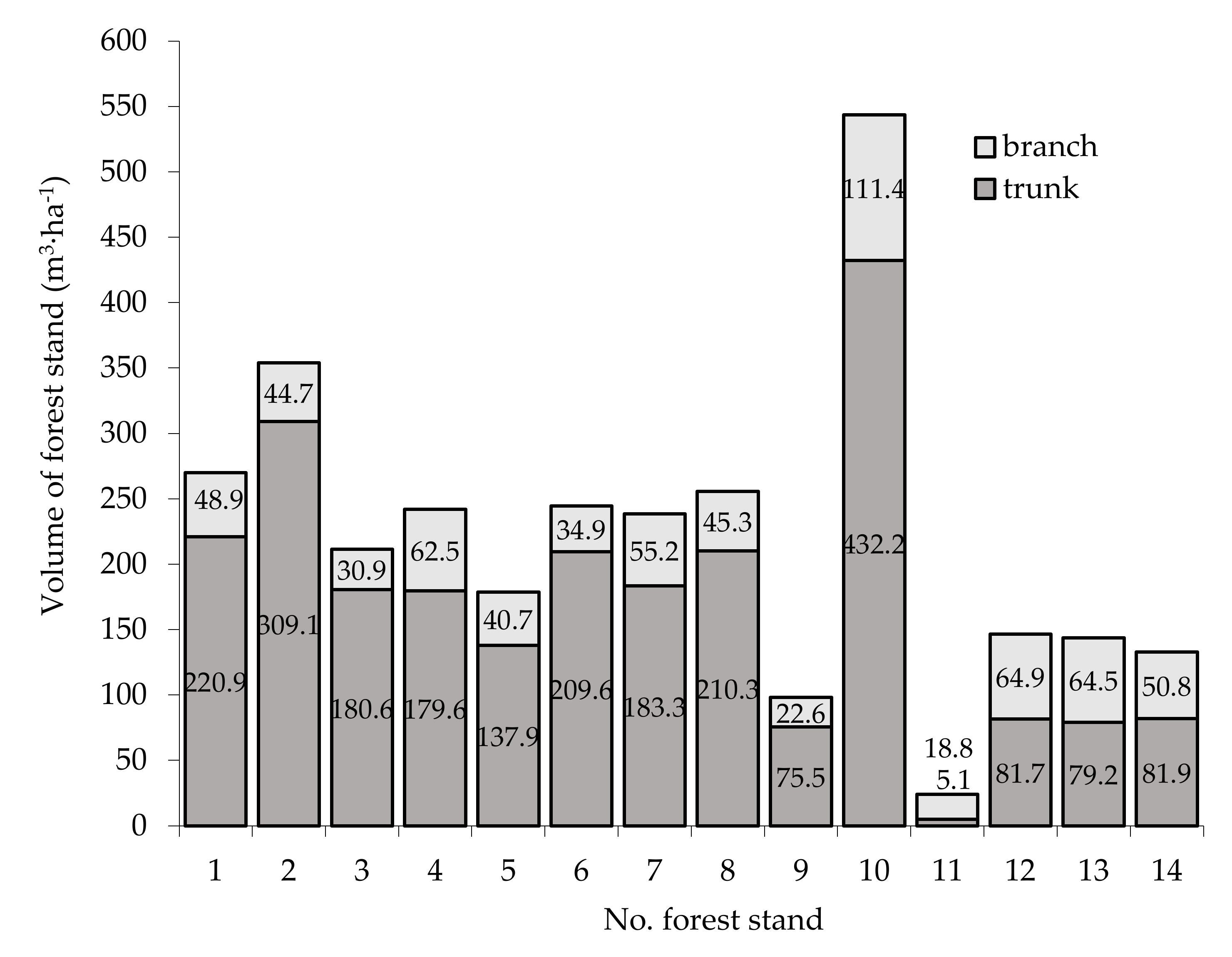
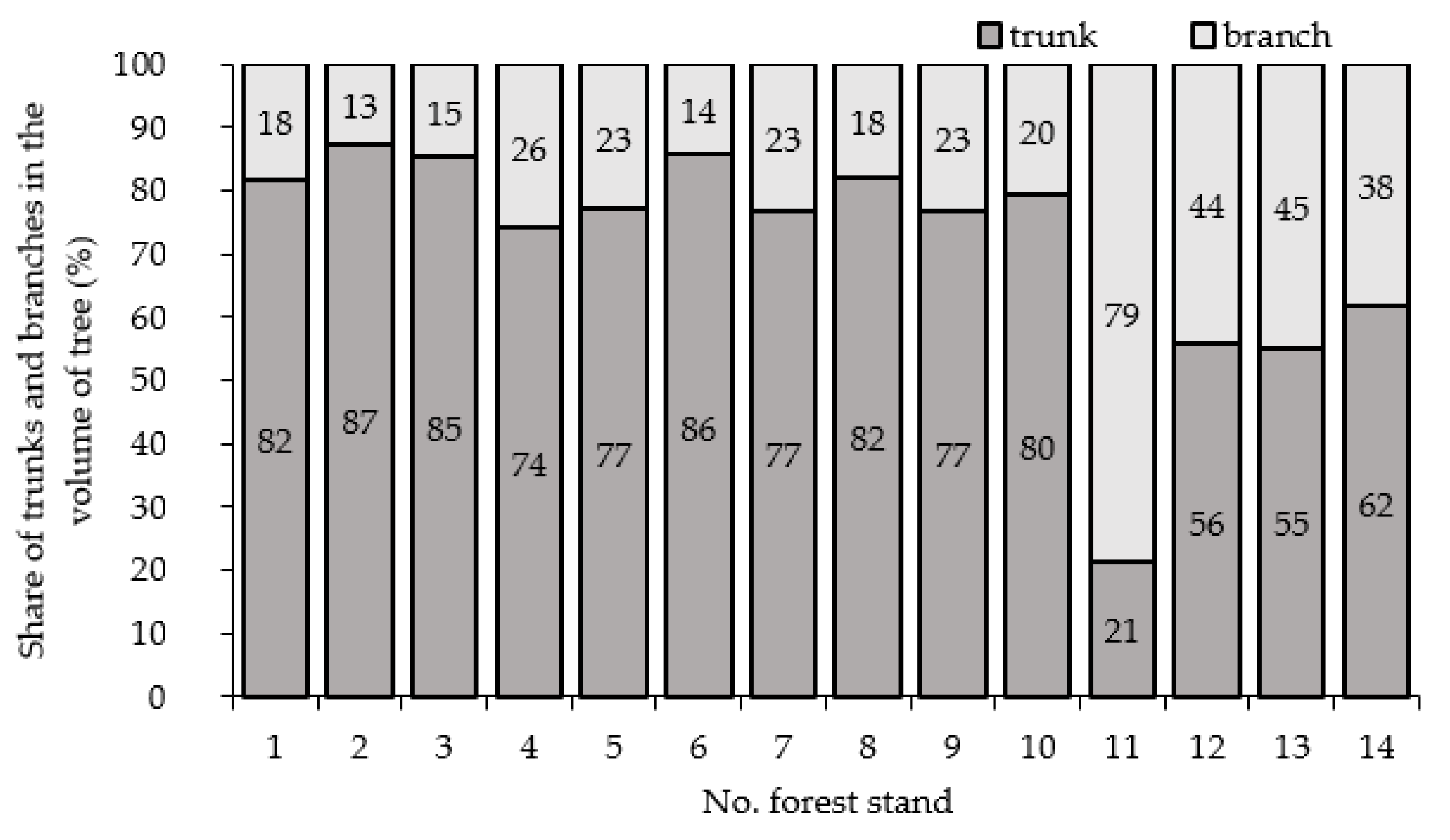
3.3. Characteristics Volume of Forest Stand
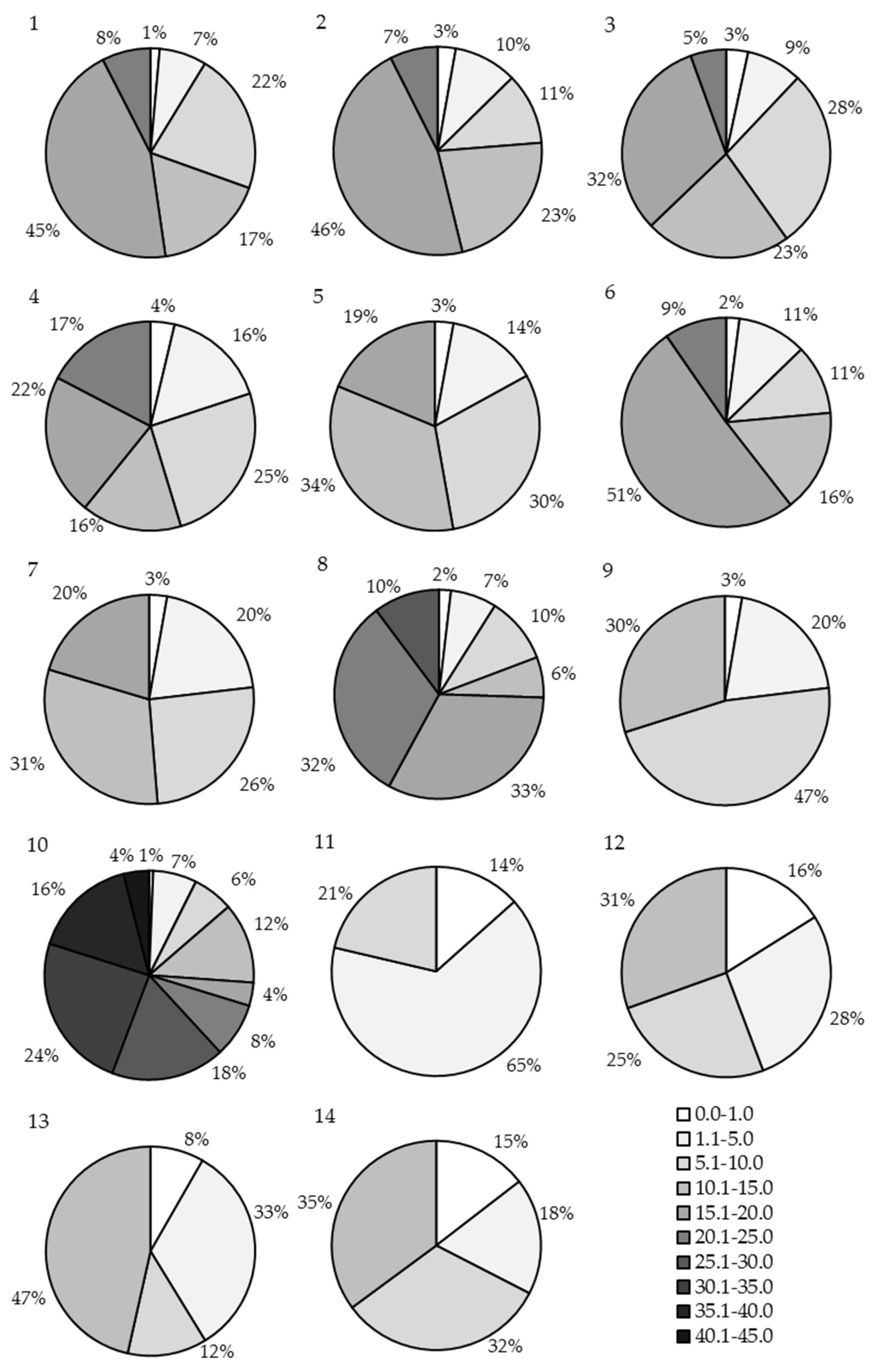
3.4. Stand Thickness Classes
4. Discussion
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. Forest Stand | Thickness Classes (cm) | Main Forest Stand | Secondary Forest Stand | Total Forest Stand | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Trunk | Branch | Σ | Trunk | Branch | Σ | Trunk | Branch | Σ | ||
| 1 | 0.0-1.0 | - | 3.1 | 3.1 | - | 0.7 | 0.7 | - | 3.8 | 3.8 |
| 1.1–5.0 | - | 17.7 | 17.7 | - | 2.0 | 2.0 | - | 19.7 | 19.7 | |
| 5.1–10.0 | 13.8 | 25.4 | 39.2 | 19.5 | - | 19.5 | 33.3 | 25.4 | 58.7 | |
| 10.1–15.0 | 43.2 | - | 43.2 | 3.3 | - | 3.3 | 46.5 | - | 46.5 | |
| 15.1–20.0 | 120.8 | - | 120.8 | - | - | - | 120.8 | - | 120.8 | |
| 20.1–25.0 | 20.3 | - | 20.3 | - | - | - | 20.3 | - | 20.3 | |
| Σ | 198.1 | 46.2 | 244.3 | 22.8 | 2.7 | 25.5 | 220.9 | 48.9 | 269.8 | |
| 2 | 0.0-1.0 | - | 7.5 | 7.5 | - | 2.4 | 2.4 | - | 9.9 | 9.9 |
| 1.1–5.0 | - | 24.9 | 24.9 | - | 9.9 | 9.9 | - | 34.8 | 34.8 | |
| 5.1–10.0 | 17.7 | - | 17.7 | 21.6 | - | 21.6 | 39.3 | - | 39.3 | |
| 10.1–15.0 | 70.9 | - | 70.9 | 8.5 | - | 8.5 | 79.4 | - | 79.4 | |
| 15.1–20.0 | 164.1 | - | 164.1 | - | - | - | 164.1 | - | 164.1 | |
| 20.1–25.0 | 26.3 | - | 26.3 | - | - | - | 26.3 | - | 26.3 | |
| Σ | 279.0 | 32.4 | 311.4 | 30.1 | 12.3 | 42.4 | 309.1 | 44.7 | 353.8 | |
| 3 | 0.0-1.0 | - | 4.1 | 4.1 | - | 3.1 | 3.1 | - | 7.2 | 7.2 |
| 1.1–5.0 | - | 10.4 | 10.4 | - | 7.9 | 7.9 | - | 18.3 | 18.3 | |
| 5.1–10.0 | 9.2 | 5.4 | 14.6 | 44.8 | - | 44.8 | 54.0 | 5.4 | 59.4 | |
| 10.1–15.0 | 38.7 | - | 38.7 | 9.3 | - | 9.3 | 48.0 | - | 48.0 | |
| 15.1–20.0 | 66.9 | - | 66.9 | - | - | - | 66.9 | - | 66.9 | |
| 20.1–25.0 | 11.7 | - | 11.7 | - | - | - | 11.7 | - | 11.7 | |
| Σ | 126.5 | 19.9 | 146.4 | 54.1 | 11.0 | 65.1 | 180.6 | 30.9 | 211.5 | |
| 4 | 0.0-1.0 | - | 3.3 | 3.3 | - | 5.7 | 5.7 | - | 9.0 | 9.0 |
| 1.1–5.0 | - | 29.4 | 29.4 | - | 9.9 | 9.9 | - | 39.3 | 39.3 | |
| 5.1–10.0 | 11.9 | 14.2 | 26.1 | 35.3 | - | 35.3 | 47.2 | 14.2 | 61.4 | |
| 10.1–15.0 | 37.5 | - | 37.5 | - | - | - | 37.5 | - | 37.5 | |
| 15.1–20.0 | 52.9 | - | 52.9 | - | - | - | 52.9 | - | 52.9 | |
| 20.1–25.0 | 42.0 | - | 42.0 | - | - | - | 42.0 | - | 42.0 | |
| Σ | 144.3 | 46.9 | 191.2 | 35.3 | 15.6 | 50.9 | 179.6 | 62.5 | 242.1 | |
| 5 | 0.0-1.0 | - | 4.1 | 4.1 | - | 1.0 | 1.0 | - | 5.1 | 5.1 |
| 1.1–5.0 | - | 22.8 | 22.8 | - | 2.5 | 2.5 | - | 25.3 | 25.3 | |
| 5.1–10.0 | 11.8 | 10.3 | 22.1 | 31.8 | - | 31.8 | 43.6 | 10.3 | 53.9 | |
| 10.1–15.0 | 60.8 | - | 60.8 | - | - | - | 60.8 | - | 60.8 | |
| 15.1–20.0 | 33.5 | - | 33.5 | - | - | - | 33.5 | - | 33.5 | |
| Σ | 106.1 | 37.2 | 143.3 | 31.8 | 3.5 | 35.3 | 137.9 | 40.7 | 178.6 | |
| 6 | 0.0-1.0 | - | 4.3 | 4.3 | - | 0.7 | 0.7 | - | 5.0 | 5.0 |
| 1.1–5.0 | - | 20.3 | 20.3 | - | 5.8 | 5.8 | - | 26.1 | 26.1 | |
| 5.1–10.0 | 6.3 | 3.8 | 10.1 | 16.4 | - | 16.4 | 22.7 | 3.8 | 26.5 | |
| 10.1–15.0 | 37.6 | - | 37.6 | 1.3 | - | 1.3 | 38.9 | - | 38.9 | |
| 15.1–20.0 | 124.5 | - | 124.5 | - | - | - | 124.5 | - | 124.5 | |
| 20.1–25.0 | 23.5 | - | 23.5 | - | - | - | 23.5 | - | 23.5 | |
| Σ | 191.9 | 28.4 | 220.3 | 17.7 | 6.5 | 24.2 | 209.6 | 34.9 | 244.5 | |
| 7 | 0.0-1.0 | - | 4.9 | 4.9 | - | 1.7 | 1.7 | - | 6.6 | 6.6 |
| 1.1–5.0 | - | 42.5 | 42.5 | - | 6.1 | 6.1 | - | 48.6 | 48.6 | |
| 5.1–10.0 | 18.5 | - | 18.5 | 42.5 | - | 42.5 | 61.0 | - | 61.0 | |
| 10.1–15.0 | 73.7 | - | 73.7 | - | - | - | 73.7 | - | 73.7 | |
| 15.1–20.0 | 48.6 | - | 48.6 | - | - | - | 48.6 | - | 48.6 | |
| Σ | 140.8 | 47.4 | 188.2 | 42.5 | 7.8 | 50.3 | 183.3 | 55.2 | 238.5 | |
| 8 | 0.0-1.0 | - | 4.7 | 4.7 | - | - | - | - | 4.7 | 4.7 |
| 1.1–5.0 | - | 18.2 | 18.2 | - | - | - | - | 18.2 | 18.2 | |
| 5.1–10.0 | 11.5 | 14.8 | 26.3 | - | - | - | 11.5 | 14.8 | 26.3 | |
| 10.1–15.0 | 8.4 | 7.6 | 16.0 | - | - | - | 8.4 | 7.6 | 16.0 | |
| 15.1–20.0 | 82.9 | - | 82.9 | - | - | - | 82.9 | - | 82.9 | |
| 20.1–25.0 | 81.4 | - | 81.4 | - | - | - | 81.4 | - | 81.4 | |
| 25.1–30.0 | 26.1 | - | 26.1 | - | - | - | 26.1 | - | 26.1 | |
| Σ | 210.3 | 45.3 | 255.6 | - | - | - | 210.3 | 45.3 | 255.6 | |
| 9 | 0.0-1.0 | - | 2.6 | 2.6 | - | - | - | - | 2.6 | 2.6 |
| 1.1–5.0 | - | 20.0 | 20.0 | - | - | - | - | 20.0 | 20.0 | |
| 5.1–10.0 | 46.2 | - | 46.2 | - | - | - | 46.2 | - | 46.2 | |
| 10.1–15.0 | 29.3 | - | 29.3 | - | - | - | 29.3 | - | 29.3 | |
| Σ | 75.5 | 22.6 | 98.1 | - | - | - | 75.5 | 22.6 | 98.1 | |
| 10 | 0.0-1.0 | - | 3.4 | 3.4 | - | - | - | - | 3.4 | 3.4 |
| 1.1–5.0 | - | 36.7 | 36.7 | - | - | - | - | 36.7 | 36.7 | |
| 5.1–10.0 | 4.2 | 29.9 | 34.1 | - | - | - | 4.2 | 29.9 | 34.1 | |
| 10.1–15.0 | 57.0 | 10.8 | 67.8 | - | - | - | 57.0 | 10.8 | 67.8 | |
| 15.1–20.0 | 8.2 | 11.4 | 19.6 | - | - | - | 8.2 | 11.4 | 19.6 | |
| 20.1–25.0 | 26.6 | 19.2 | 45.8 | - | - | - | 26.6 | 19.2 | 45.8 | |
| 25.1–30.0 | 95.7 | - | 95.7 | - | - | - | 95.7 | - | 95.7 | |
| 30.1–35.0 | 131.2 | - | 131.2 | - | - | - | 131.2 | - | 131.2 | |
| 35.1–40.0 | 87.9 | - | 87.9 | - | - | - | 87.9 | - | 87.9 | |
| 40.1–45.0 | 21.4 | - | 21.4 | - | - | - | 21.4 | - | 21.4 | |
| Σ | 432.2 | 111.4 | 543.6 | - | - | - | 432.2 | 111.4 | 543.6 | |
| 11 | 0.0-1.0 | - | 3.2 | 3.2 | - | - | - | - | 3.2 | 3.2 |
| 1.1–5.0 | - | 15.6 | 15.6 | - | - | - | - | 15.6 | 15.6 | |
| 5.1–10.0 | 5.1 | - | 5.1 | - | - | - | 5.1 | - | 5.1 | |
| Σ | 5.1 | 18.8 | 23.9 | - | - | - | 5.1 | 18.8 | 23.9 | |
| 12 | 0.0-1.0 | - | 23.6 | 23.6 | - | - | - | - | 23.6 | 23.6 |
| 1.1–5.0 | - | 41.3 | 41.3 | - | - | - | - | 41.3 | 41.3 | |
| 5.1–10.0 | 37.0 | - | 37.0 | - | - | - | 37.0 | - | 37.0 | |
| 10.1–15.0 | 44.7 | - | 44.7 | - | - | - | 44.7 | - | 44.7 | |
| Σ | 81.7 | 64.9 | 146.6 | - | - | - | 81.7 | 64.9 | 146.6 | |
| 13 | 0.0-1.0 | - | 11.9 | 11.9 | - | - | - | - | 11.9 | 11.9 |
| 1.1–5.0 | - | 47.4 | 47.4 | - | - | - | - | 47.4 | 47.4 | |
| 5.1–10.0 | 12.4 | 5.2 | 17.6 | - | - | - | 12.4 | 5.2 | 17.6 | |
| 10.1–15.0 | 66.8 | - | 66.8 | - | - | - | 66.8 | - | 66.8 | |
| Σ | 79.2 | 64.5 | 143.7 | - | - | - | 79.2 | 64.5 | 143.7 | |
| 14 | 0.0-1.0 | - | 19.3 | 19.3 | - | - | - | - | 19.3 | 19.3 |
| 1.1–5.0 | - | 23.9 | 23.9 | - | - | - | - | 23.9 | 23.9 | |
| 5.1–10.0 | 35.3 | 7.6 | 42.9 | - | - | - | 35.3 | 7.6 | 42.9 | |
| 10.1–15.0 | 46.6 | - | 46.6 | - | - | - | 46.6 | - | 46.6 | |
| Σ | 81.9 | 50.8 | 132.7 | - | - | - | 81.9 | 50.8 | 132.7 | |
References
- Huntley, J.C. Robinia pseudoacacia L. black locust. In Silvics of North. America: Volume 2—Hardwoods; Agriculture Handbook 654; United States Department of Agriculture: Washington, DC, USA, 1990; pp. 755–761. [Google Scholar]
- Rédei, K.; Csiha, I.; Keserû, Z.; Gál, J. Influence of Regeneration Method on the Yield and Stem Quality of Black Locust (Robinia pseudoacacia L.) Stands: A Case Study. Acta Silv. Lign. Hung. 2012, 8, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Andrašev, S.; Rončević, S.; Ivanišević, P.; Pekeč, S.; Bobinac, M. Productivity of black locust (Robinia pseudoacacia L.) stands on chernozem in Vojvodina. Bull. Fac. For. 2014, 10, 9–32. [Google Scholar] [CrossRef]
- Pacyniak, C. Black locust (Robinia pseudoacacia L.) in the conditions of the forest environment in Poland. Rocz. AR Pozn. Rozpr. Nauk. 1981, 111, 1–85. (In Polish) [Google Scholar]
- Strączyńska, S.; Strączyński, S. Characteristics of the habitat conditions of black locust (Robinia pseudacacia L.) planting in a combustion waste landfill. Zesz. Probl. Post. Nauk Rol. 2004, 501, 417–423. (In Polish) [Google Scholar]
- Kraszkiewicz, A. Chemical composition and selected energy properties of black locust bark (Robinia pseudoacacia L.). Agric. Eng. 2016, 20, 117–124. [Google Scholar] [CrossRef]
- Kraszkiewicz, A. The Combustion of Wood Biomass in Low Power Coal-Fired Boilers. Combust. Sci. Technol. 2015, 188, 389–396. [Google Scholar] [CrossRef]
- Bilkic, B.; Haykiri-Acma, H.; Yaman, S. Combustion reactivity estimation parameters of biomass compared with lignite based on thermogravimetric analysis. Energy Sources Part A Recover. Util. Environ. Eff. 2020, 1–14. [Google Scholar] [CrossRef]
- Bender, J.; Gilewska, M.; Wójcik, A. Usefulness of black locust for woodlot in post-mining land. Arch. Ochr. Środow. 1985, 3–4, 113–133. (In Polish) [Google Scholar]
- Gilewska, M. Biological reclamation of ash landfills from lignite. Rocz. Glebozn. 2004, 55, 103–110. (In Polish) [Google Scholar]
- Węgorek, T. Changes in some properties of the earth material and development of phytocoenoses in the external dump of sulfur mine as a result of target forest reclamation. Rozpr. Nauk. AR Lublinie 2003, 275, 1–140. (In Polish) [Google Scholar]
- Carl, C.; Biber, P.; Veste, M.; Landgraf, D.; Pretzsch, H. Key drivers of competition and growth partitioning among Robinia pseudoacacia L. trees. For. Ecol. Manag. 2018, 430, 86–93. [Google Scholar] [CrossRef]
- Seserman, D.M.; Veste, M.; Freese, D. Optimisation of biomass productivity of black locust (Robinia pseudoacacia L.) on marginal lands—A case study in Lower Lusatia, NE Germany. In Proceedings of the 19th EGU General Assembly (EGU2017), Vienna, Austria, 23–28 April 2017. [Google Scholar]
- Nicolescu, V.N.; Buzatu-Goant, C.; Bakti, B.; Keserű, Z.; Antal, B.; Rédei, K. Black locust (Robinia pseudoacacia L.) as a multi-purpose tree species in Hungary and Romania: A review. J. For. Res. 2018, 29, 1449–1463. [Google Scholar] [CrossRef]
- González, I.; Sixto, H.; Rodríguez-Soalleiro, R.; Oliveira, N. Nutrient Contribution of Litterfall in a Short Rotation Plantation of Pure or Mixed Plots of Populus alba L. and Robinia pseudoacacia L. Forests 2020, 11, 1133. [Google Scholar] [CrossRef]
- Orlik, T.; Węgorek, T.; Zubala, T. Efficiency and growth of warty birch, European larch and black locust in mid-field strip plantations. Folia Univ. Agric. Stetin., Agric. 2001, No. 87(217). 167–170. (In Polish) [Google Scholar]
- Węgorek, T.; Kraszkiewicz, A. Growth dynamics of false acacia (Robinia pseudacacia L.) in middle-field shelterbelt on loess soils. Acta Agrophys. 2005, 5, 211–218. (In Polish) [Google Scholar]
- European Commission. Communication from The Commission to The European Parliament, The Council, The European Economic and Social Committee and The Committee of The Regions Energy Roadmap 2050. Available online: https://eur-lex.europa.eu/legal-content/PL/TXT/PDF/?uri=CELEX:52011DC0885&from=EN (accessed on 28 December 2020).
- Renewable Energy Development Strategy, 2000: Report of the Ministry of the Environment Republic of Poland, Warsaw. Available online: https://ieo.pl/pl/raporty/82-strategia-rozwoju-energetyki-odnawialnej/file (accessed on 27 January 2021). (In Polish).
- Poland’s Energy Policy until 2030. Ministry of Economy: Warsaw, Poland, 10 November 2009. Available online: http://nfosigw.gov.pl/download/gfx/nfosigw/pl/nfoopisy/1328/1/4/polityka_energetyczna_polski_do_2030r.pdf (accessed on 11 January 2021). (In Polish)
- Abbasi, T.; Abbasi, S.A. Biomass energy and the environmental impacts associated with its production and utilization. Renew. Sust. Energ. Rev. 2010, 14, 919–937. [Google Scholar] [CrossRef]
- Evans, A.; Strezov, V.; Evans, T.J. Sustainability considerations for electricity generation from biomass. Renew. Sust. Energ. Rev. 2010, 14, 1419–1427. [Google Scholar] [CrossRef]
- Ferrè, C.; Comolli, R. Comparison of soil CO2 emissions between short-rotation coppice poplar stands and arable lands. iForest Biogeosci. For. 2018, 11, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Landgraf, D.; Carl, C.; Neupert, M. Biomass Yield of 37 Different SRC Poplar Varieties Grown on a Typical Site in North Eastern Germany. Forests 2020, 11, 1048. [Google Scholar] [CrossRef]
- Cichy, W. Wood in combined heat and power plants—Innovation or sabotage? Ekopartner 2005, 2, 8–9. (In Polish) [Google Scholar]
- Szpil, Z. Energy prices, market—Ecology—Diversification. Aura 2006, 3, 18–19. (In Polish) [Google Scholar]
- Update of the National Program for Increasing Forest Cover 2014. Ministry of the Environment, Sękocin Stary 2014. Available online: https://nfosigw.gov.pl/download/gfx/nfosigw/pl/nfoekspertyzy/858/184/1/2013-772.pdf (accessed on 19 January 2021). (In Polish)
- Lewandowski, P. Renewable Energy in Western Pomerania; Wyd. Hogben: Szczecin, Poland, 2005; p. 326. (In Polish) [Google Scholar]
- Mantovani, D.; Veste, M.; Freese, D. How Much Water is Used by a Black Locust (Robinia pseudoacacia L.) Short-Rotation Plantation on Degraded Soil? Tagungsbeitrag zu: Jahrestagung der DBG Kommission IV Titel der Tagung: Böden verstehen—Böden nutzen—Böden fit machen Veranstalter: DBG, September 2011, Berlin Berichte der DBG. Available online: https://www.researchgate.net/publication/235695650 (accessed on 1 April 2021).
- Mantovani, D.; Veste, M.; Freese, D. Black locust (Robinia pseudoacacia L.) ecophysiological and morphological adaptations to drought andtheir consequence on biomass production and water-use efficiency. N. Z. J. For. Sci. 2014, 44, 29. [Google Scholar] [CrossRef] [Green Version]
- Plomion, C.; Leprovost, G.; Stokes, A. Wood Formation in Trees. Plant. Physiol. 2001, 127, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Fabisiak, E. Variability of basic anatomical elements and wood density of selected tree species. Rocz. AR Poznaniu. Rozpr. Nauk. 2005, 369, 1–176. (In Polish) [Google Scholar]
- Rocha, M.F.V.; Veiga, T.R.L.A.; Soares, B.C.h.D.; Araújo, A.C.C.; Carvalho, A.M.M.; Hein, P.R.G. Do the Growing Conditions of Trees Influence the Wood Properties? Floresta Ambient. 2019, 26, e20180353. [Google Scholar] [CrossRef]
- Bach, E.M.; Ramirez, K.S.; Fraser, T.D.; Wall, D.H. Soil Biodiversity Integrates Solutions for a Sustainable Future. Sustainability 2020, 12, 2662. [Google Scholar] [CrossRef] [Green Version]
- Bruchwald, A. Natural basics of building growth models. Sylwan 1988, 11, 1–10. (In Polish) [Google Scholar]
- Cui, Y.; Bi, H.; Liu, S.; Hou, G.; Wang, N.; Ma, X.; Zhao, D.; Wang, S.; Yun, H. Developing Additive Systems of Biomass Equations for Robinia pseudoacacia L. in the Region of Loess Plateau of Western Shanxi Province, China. Forests 2020, 11, 1332. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Li, F. Developing Two Additive Biomass Equations for Three Coniferous Plantation Species in Northeast China. Forests 2016, 7, 136. [Google Scholar] [CrossRef] [Green Version]
- Yuen, J.Q.; Fung, T.; Ziegler, A.D. Review of allometric equations for major land covers in SE Asia: Uncertainty and implications for above- and below-ground carbon estimates. For. Ecol. Manag. 2016, 360, 323–340. [Google Scholar] [CrossRef]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Change Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wang, X.; Ouyang, Z.; Lu, F.; Feng, L.; Tao, J. A review of biomass equations for China’s tree species. Earth Syst. Sci. Data 2020, 12, 21–40. [Google Scholar] [CrossRef] [Green Version]
- Paul, K.I.; Roxburgh, S.H.; Chave, J.; England, J.R.; Zerihun, A.; Specht, A.; Lewis, T.; Bennett, L.T.; Baker, T.G.; Adams, M.A.; et al. Testing the generality of above-ground biomass allometry across plant functional types at the continent scale. Glob. Chang. Biol. 2016, 22, 2106–2124. [Google Scholar] [CrossRef] [PubMed]
- Basuki, T.; Van Laake, P.; Skidmore, A.; Hussin, Y. Allometric equations for estimating the above-ground biomass in tropical lowland Dipterocarp forests. For. Ecol. Manag. 2009, 257, 1684–1694. [Google Scholar] [CrossRef]
- Kuyah, S.; Dietz, J.; Muthuri, C.; Jamnadass, R.; Mwangi, P.; Coe, R.; Neufeldt, H. Allometric equations for estimating biomass in agricultural landscapes: I. Aboveground biomass. Agric. Ecosyst. Environ. 2012, 158, 216–224. [Google Scholar] [CrossRef]
- Zielony, R.; Kliczkowska, A. Poland’s Natural and Forest Regionalization 2010; CILP: Warsaw, Poland, 2012; pp. 1–359. (In Polish) [Google Scholar]
- Ziemnicki, S.; Fijałkowski, D.; Repelewska-Pękalowa, J.; Węgorek, T. Reclamation of an Opencast Mine Heap (on the Example of Piaseczno); PWN: Warsaw, Poland, 1980; pp. 1–100. (In Polish) [Google Scholar]
- Kraszkiewicz, A.; Węgorek, T. Structure of robinia stands on sandy slopes threatened with erosion. Zesz. Nauk. Akad. Rol. Poznaniu. CCCLXXV Rol. 2006, 65, 69–74. (In Polish) [Google Scholar]
- Bruchwald, A. Dendrometry; Wyd. SGGW: Warsaw, Poland, 1999; pp. 1–261. (In Polish) [Google Scholar]
- Kowalkowski, A. Mineral. Fertilization of Stands; SGGW-AR: Warsaw, Poland, 1982; pp. 1–86. (In Polish) [Google Scholar]
- Baule, H.; Fricker, C. Fertilization of Forest Trees; PWRiL: Warsaw, Poland, 1971; pp. 1–314. (In Polish) [Google Scholar]
- Pokojska, U. The role of humus in shaping the reaction, buffer properties and ion-exchange capacity of forest soils. Rocz. Glebozn. 1986, 37, 249–262. (In Polish) [Google Scholar]
- Szczukowski, S.; Tworkowski, J.; Stolarski, M.; Grzelczyk, M. Productivity of willow (Salix spp.) and characterization of willow biomass as fuel. Zesz. Probl. Post. Nauk Rol. 2005, 507, 495–503. [Google Scholar]
- Zajączkowski, K.; Kwiecień, R.; Zajączkowska, B.; Wajda, T.; Zawadzki, M. Production Possibilities of Selected Varieties of Poplar and Willow in Plantations with a Shortened Cycle; IBL: Warsaw, Poland, 2001; pp. 1–66. (In Polish) [Google Scholar]
- Niemczyk, M.; Wojda, T.; Kantorowicz, W. The breeding usefulness of selected poplar varieties in energy plantations with a short production cycle. Sylwan 2016, 160, 292–298. (In Polish) [Google Scholar]
- Szostak, A.; Bidzińska, G.; Ratajczak, E.; Herbeć, M. Wood biomass from plantations of fast-growing trees as an alternative source of wood raw material in Poland. Drewno 2013, 56, 85–112. [Google Scholar] [CrossRef]
- Dimobe, K.; Mensah, S.; Goetze, D.; Ouédraogo, A.; Kuyah, S.; Porembski, S.; Thiombiano, A. Aboveground biomass partitioning and additive models for Combretum glutinosum and Terminalia laxiflora inWest Africa. Biomass Bioenergy 2018, 115, 151–159. [Google Scholar] [CrossRef]
- Júnior, L.R.N.; Engel, V.L.; Parrotta, J.A.; De Melo, A.C.G.; Ré, D.S. Allometric equations for estimating tree biomass in restored mixed-species Atlantic Forest stands. Biota Neotrop. 2014, 14, 1–9. [Google Scholar]
- Dong, L.H.; Zhang, L.J.; Li, F. Additive Biomass Equations Based on Di_erent Dendrometric V ariables for Two Dominant Species (Larix gmelini Rupr. and Betula platyphylla Suk.) in Natural Forests in the Eastern Daxing’an Mountains, Northeast China. Forests 2018, 9, 261. [Google Scholar] [CrossRef] [Green Version]
- Riofrío, J.; Herrero, C.; Grijalva, J.; Bravo, F. Aboveground tree additive biomass models in Ecuadorian highland agroforestry systems. Biomass Bioenergy 2015, 80, 252–259. [Google Scholar] [CrossRef]




| Forest Stand/Area Number | Soil | Surface Coverage (%) | Litter (cm) | Humus Layer (cm) | ||
|---|---|---|---|---|---|---|
| Trees | Shrubs | Undergrowth | ||||
| 1 | sand | 80 | 60 | 30 | 2 | 4 |
| 2 | 80 | 60 | 30 | 2 | 2 | |
| 3 | 80 | 20 | 70 | 4 | 5 | |
| 4 | 80 | 20 | 70 | 5 | 7 | |
| 5 | 80 | 20 | 100 | 5 | 8 | |
| 6 | clay | 70 | 50 | 30 | 1 | 5 |
| 7 | 80 | 30 | 70 | 2 | 6 | |
| 8 | 50 | 50 | 80 | 3 | 10 | |
| 9 | dust | 80 | 20 | 80 | 3 | 8 |
| 10 | 50 | 50 | 60 | 5 | 10 | |
| 11 | − | 70 | 90 | 1 | 20 | |
| 12 | 90 | 60 | 90 | 1 | 24 | |
| 13 | 90 | 60 | 90 | 1 | 24 | |
| 14 | 90 | 60 | 90 | 1 | 22 | |
| Forest Stand/Area Number | Content of Nutrients (g·kg−1) and Soil Abundance | C:N | |||
|---|---|---|---|---|---|
| Ntotal | P | K | Corganic | ||
| 1 | 0.36 insufficient | 0.004 insufficient | 0.021 insufficient | 2.09 | 5.81 |
| 2 | 0.38 insufficient | 0.005 insufficient | 0.027 insufficient | 2.59 | 6.82 |
| 3 | 0.24 insufficient | 0.004 insufficient | 0.024 insufficient | 2.21 | 9.21 |
| 4 | 0.57 insufficient | 0.004 insufficient | 0.023 insufficient | 2.71 | 4.75 |
| 5 | 0.41 insufficient | 0.005 insufficient | 0.024 insufficient | 2.90 | 7.07 |
| 6 | 0.77 medium | 0.054 medium | 0.142 good | 6.60 | 8.57 |
| 7 | 0.84 medium | 0.004 insufficient | 0.030 insufficient | 4.32 | 5.14 |
| 8 | 0.78 medium | 0.007 insufficient | 0.025 insufficient | 6.05 | 7.76 |
| 9 | x | x | x | x | x |
| 10 | 1.23 medium | 0.015 medium | 0.028 insufficient | 6.79 | 5.52 |
| 11 | 0.43 insufficient | 0.059 medium | 0.063 medium | 3.62 | 8.42 |
| 12 | 1.95 good | 0.176 good | 0.260 good | 10.37 | 5.32 |
| 13 | 1.34 good | 0.101 good | 0.126 good | 8.74 | 6.52 |
| 14 | 0.57 insufficient | 0.109 good | 0.087 medium | 4.18 | 7.33 |
| Forest Stand/Area Number | Age (Years) | Number of Trees (Pcs. ha−1) | Average Height (m) | Average Diameter at Breast Height (cm) | |||
|---|---|---|---|---|---|---|---|
| Main | Secondary | Main | Secondary | Main | Secondary | ||
| 1 | 35 | 919 | 588 | 16.5 | 12.5 | 19.0 | 9.5 |
| 2 | 35 | 1029 | 882 | 18.0 | 13.0 | 19.5 | 10.0 |
| 3 | 35 | 750 | 1525 | 15.5 | 11.0 | 17.5 | 8.5 |
| 4 | 35 | 580 | 1440 | 17.0 | 11.5 | 23.5 | 8.5 |
| 5 | 35 | 943 | 1057 | 15.5 | 10.0 | 16.0 | 8.5 |
| 6 | 35 | 650 | 575 | 19.0 | 12.0 | 21.0 | 9.0 |
| 7 | 35 | 870 | 1111 | 18.0 | 11.5 | 18.0 | 9.0 |
| 8 | 41 | 410 | - | 24.5 | - | 26.5 | - |
| 9 | 33 | 1840 | - | 11.5 | - | 11.0 | - |
| 10 | 64 | 320 | - | 24.0 | - | 38.5 | - |
| 11 | 4 | 1720 | - | 2.0 | - | 4.5 | - |
| 12 | 8 | 1905 | - | 7.5 | - | 12.0 | - |
| 13 | 8 | 1828 | - | 8.0 | - | 11.5 | - |
| 14 | 8 | 1715 | - | 7.5 | - | 12.0 | - |
| Forest Stand/Area Number | Age (Years) | Soil | Trendline Equation | Coefficient of Determination R2 |
|---|---|---|---|---|
| 1 | 35 | sand | y = −6.2696x2 + 55.296x − 57.73 | 0.4264 |
| 2 | 35 | y = −6.6857x2 + 62.937x − 66.98 | 0.3563 | |
| 3 | 35 | y = −3.7768x2 + 33.055x − 34.01 | 0.4516 | |
| 4 | 35 | y = −1.9679x2 + 21.644x − 14.04 | 0.8575 | |
| 5 | 35 | y = −3.7571x2 + 32.223x − 26.68 | 0.6532 | |
| 6 | 35 | clay | y = -3.5107x2 + 37.035x − 39.66 | 0.3206 |
| 7 | 35 | y = −3.3x2 + 31.66x − 21.04 | 0.5409 | |
| 8 | 41 | y = −2.8286x2 + 31.457x − 32.743 | 0.4643 | |
| 9 | 33 | dust | y = −8.575x2 + 53.505x − 44.925 | 0.8645 |
| 10 | 64 | y = −1.9674x2 + 28.404x − 26.117 | 0.3982 | |
| 11 | 4 | y = −11.45x2 + 46.75x − 32.1 | 1 | |
| 12 | 8 | y = −2.5x2 + 18.4x + 9.4 | 0.775 | |
| 13 | 8 | y = 3.425x2 − 3.635x + 19.325 | 0.4789 | |
| 14 | 8 | y = −0.225x2 + 11.215x + 6.825 | 0.9203 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kraszkiewicz, A. Productivity of Black Locust (Robinia pseudoacacia L.) Grown on a Varying Habitats in Southeastern Poland. Forests 2021, 12, 470. https://doi.org/10.3390/f12040470
Kraszkiewicz A. Productivity of Black Locust (Robinia pseudoacacia L.) Grown on a Varying Habitats in Southeastern Poland. Forests. 2021; 12(4):470. https://doi.org/10.3390/f12040470
Chicago/Turabian StyleKraszkiewicz, Artur. 2021. "Productivity of Black Locust (Robinia pseudoacacia L.) Grown on a Varying Habitats in Southeastern Poland" Forests 12, no. 4: 470. https://doi.org/10.3390/f12040470
APA StyleKraszkiewicz, A. (2021). Productivity of Black Locust (Robinia pseudoacacia L.) Grown on a Varying Habitats in Southeastern Poland. Forests, 12(4), 470. https://doi.org/10.3390/f12040470