
Vertical Distribution of Mercury in Forest Soils and Its Transfer to Edible Mushrooms in Relation to Tree Species

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
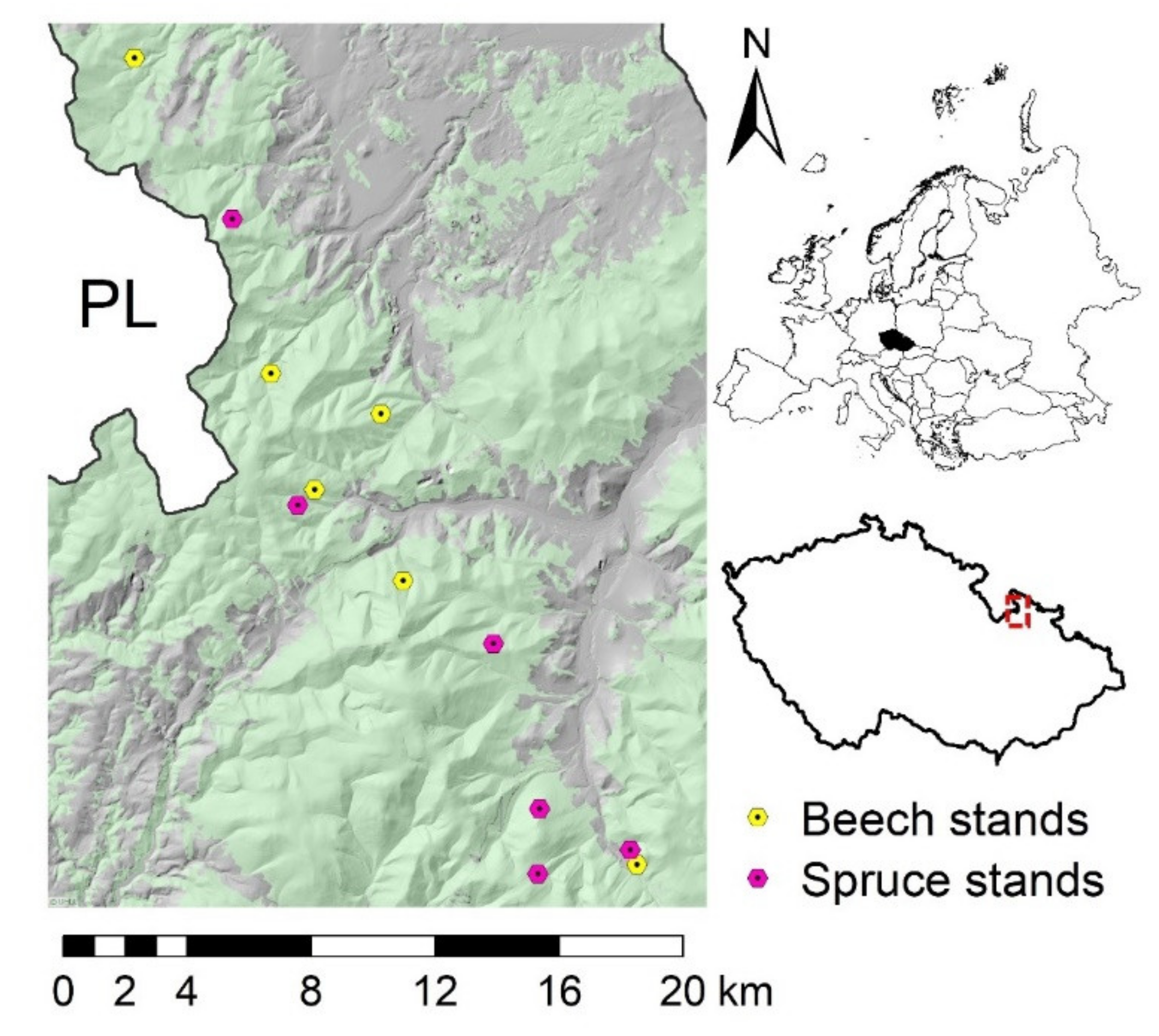
2.1. Study Area
2.2. Soil and Mushroom Sampling
2.3. Sample Preparation and Analysis
2.4. Mercury Accumulation in Mushrooms and Health Risk Assessment
2.5. Spatial and Statistical Analysis
3. Results
3.1. Effect of Soil Properties on Soil Profile Hg Distribution
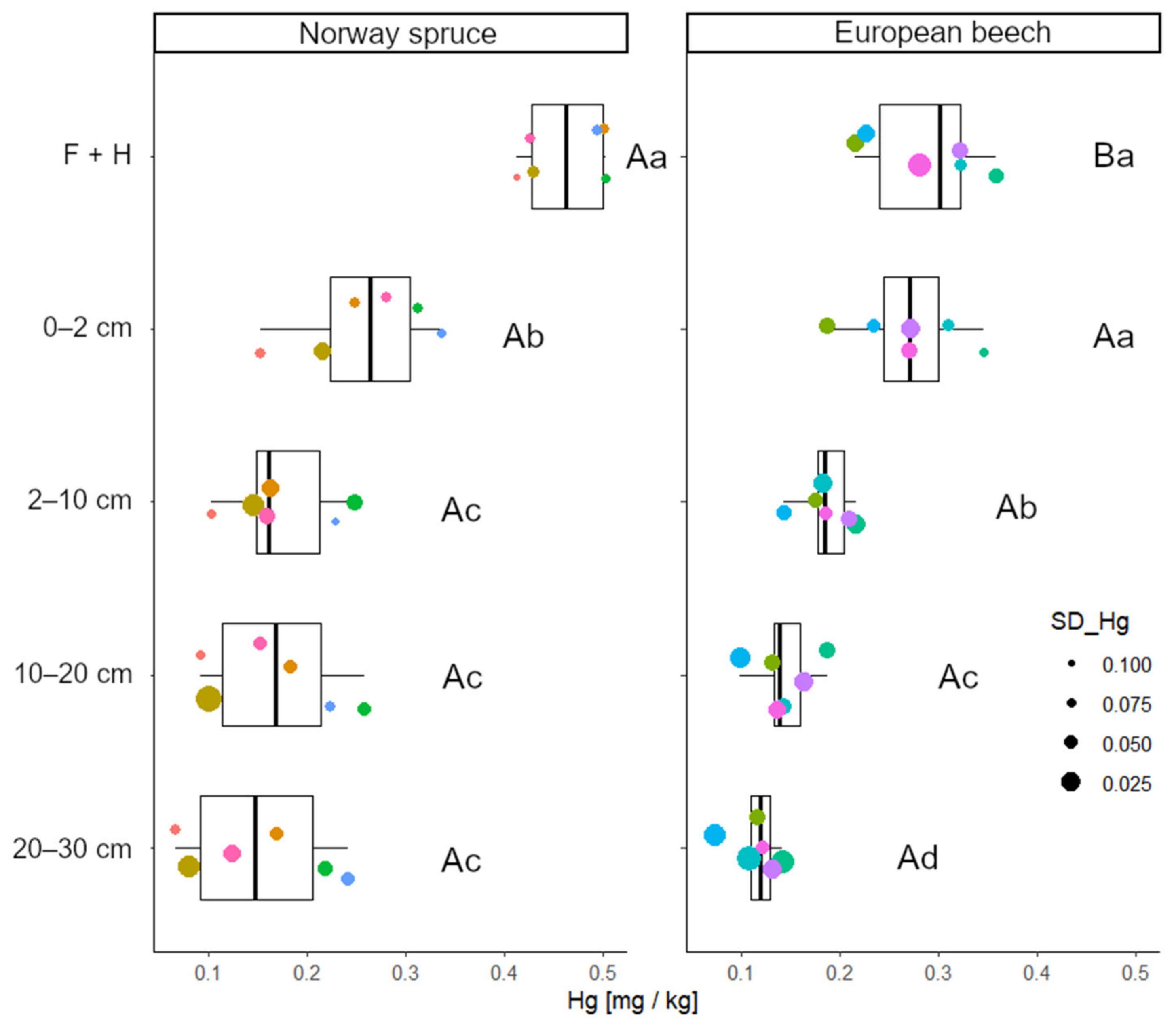
3.2. Tree-Species Effects on Hg Distribution in the Soil Profile
3.3. Effect of Tree Species on the Bioaccumulation of Hg in Mushrooms and the Related Health Risks
4. Discussion
4.1. Effect of Soil Properties on Soil Profile Hg Distribution
4.2. Effect of Tree Species on the Soil Profile Hg Distribution
4.3. Effect of Tree Species on the Hg Transfer to Mushrooms and Related Health Risks
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Variable | Species | F+H | 0–2 cm | 2–10 cm | 10–20 cm | 20–30 cm | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lower | Mean | Upper | a | Lower | Mean | Upper | a | Lower | Mean | Upper | a | Lower | Mean | Upper | a | Lower | Mean | Upper | a | ||
| Hg (mg/kg) | NS | 0.427 | 0.461 | 0.496 | *** | 0.230 | 0.264 | 0.298 | 0.157 | 0.180 | 0.204 | 0.127 | 0.156 | 0.185 | 0.102 | 0.133 | 0.164 | ||||
| EB | 0.188 | 0.287 | 0.387 | ||||||||||||||||||
| C (g/kg) | NS | 357 | 376 | 396 | ** | 109 | 120 | 131 | 57.3 | 64.9 | 72.5 | 39.2 | 47.9 | 56.6 | 30.1 | 39.8 | 49.6 | ||||
| EB | 266 | 316 | 366 | ||||||||||||||||||
| N (g/kg) | NS | 16.3 | 16.9 | 17.6 | 5.16 | 6.12 | 7.09 | * | 3.38 | 3.90 | 4.43 | 2.32 | 2.82 | 3.31 | 1.73 | 2.26 | 2.78 | ||||
| EB | 5.30 | 7.67 | 10.0 | ||||||||||||||||||
| S (g/kg) | NS | 2.05 | 2.17 | 2.28 | ** | 0.624 | 0.692 | 0.760 | 0.417 | 0.486 | 0.555 | 0.291 | 0.362 | 0.432 | 0.240 | 0.306 | 0.372 | ||||
| EB | 1.43 | 1.78 | 2.13 | ||||||||||||||||||
| C:N | NS | 20.9 | 21.7 | 22.5 | *** | 17.9 | 19.3 | 20.7 | ** | 16.4 | 19.0 | 21.5 | * | 16.6 | 19.6 | 22.6 | * | 17.6 | 20.7 | 23.9 | * |
| EB | 16.9 | 19.0 | 21.1 | 12.8 | 16.0 | 19.1 | 9.4 | 15.1 | 20.8 | 8.5 | 15.2 | 21.9 | 8.3 | 15.3 | 22.4 | ||||||
| pH | NS | 3.64 | 3.77 | 3.90 | *** | 3.76 | 3.89 | 4.01 | * | 4.03 | 4.11 | 4.20 | ** | 4.28 | 4.39 | 4.51 | ** | 4.49 | 4.59 | 4.70 | * |
| EB | 4.06 | 4.51 | 4.95 | 3.83 | 4.23 | 4.62 | 4.16 | 4.46 | 4.76 | 4.39 | 4.72 | 5.04 | 4.58 | 4.90 | 5.22 | ||||||
| Sand (%) | NS | n.a. | n.a. | n.a. | 34.6 | 38.7 | 42.8 | 36.2 | 40.8 | 45.5 | 37.8 | 42.6 | 47.4 | 41.1 | 45.2 | 49.4 | |||||
| EB | |||||||||||||||||||||
| Silt (%) | NS | n.a. | n.a. | n.a. | 50.1 | 54.2 | 58.3 | 48.1 | 53.2 | 58.2 | 46.8 | 52.0 | 57.2 | 45.3 | 49.7 | 54.2 | |||||
| EB | |||||||||||||||||||||
| Clay (%) | NS | n.a. | n.a. | n.a. | 6.23 | 7.09 | 7.95 | 5.16 | 6.05 | 6.93 | 4.38 | 5.40 | 6.42 | 4.00 | 5.02 | 6.03 | |||||
| EB | |||||||||||||||||||||
| Alo (g/kg) | NS | n.a. | n.a. | n.a. | 1.67 | 2.60 | 3.53 | ** | 2.22 | 3.81 | 5.40 | · | 4.86 | 6.57 | 8.28 | 5.56 | 7.67 | 9.78 | |||
| EB | 2.36 | 4.79 | 7.21 | 2.00 | 5.97 | 9.94 | |||||||||||||||
| Feo (g/kg) | NS | n.a. | n.a. | n.a. | 5.11 | 6.42 | 7.74 | 5.80 | 7.90 | 10.01 | 5.61 | 7.87 | 10.14 | 4.84 | 6.79 | 8.75 | |||||
| EB | |||||||||||||||||||||
References
- Wang, D.; Shi, X.; Wei, S. Accumulation and transformation of atmospheric mercury in soil. Sci. Total. Environ. 2003, 304, 209–214. [Google Scholar] [CrossRef]
- Kim, K.-H.; Kabir, E.; Jahan, S.A. A review on the distribution of Hg in the environment and its human health impacts. J. Hazard. Mater. 2016, 306, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Beckers, F.; Rinklebe, J. Cycling of mercury in the environment: Sources, fate, and human health implications: A review. Crit. Rev. Environ. Sci. Technol. 2017, 47, 693–794. [Google Scholar] [CrossRef]
- Frossard, A.; Hartmann, M.; Frey, B. Tolerance of the forest soil microbiome to increasing mercury concentrations. Soil Biol. Biochem. 2017, 105, 162–176. [Google Scholar] [CrossRef]
- Boszke, L.; Kowalski, A.; Astel, A.; Barański, A.; Gworek, B.; Siepak, J. Mercury mobility and bioavailability in soil from contaminated area. Environ. Earth Sci. 2007, 55, 1075–1087. [Google Scholar] [CrossRef]
- Liu, S.; Wang, X.; Guo, G.; Yan, Z. Status and environmental management of soil mercury pollution in China: A review. J. Environ. Manag. 2020, 277, 111442. [Google Scholar] [CrossRef] [PubMed]
- Lamborg, C.H.; Fitzgerald, W.F.; Damman, A.W.H.; Benoit, J.M.; Balcom, P.H.; Engstrom, D.R. Modern and historic atmospheric mercury fluxes in both hemispheres: Global and regional mercury cycling implications. Glob. Biogeochem. Cycles 2002, 16, 51-1. [Google Scholar] [CrossRef]
- Ballabio, C.; Jiskra, M.; Osterwalder, S.; Borrelli, P.; Montanarella, L.; Panagos, P. A spatial assessment of mercury content in the European Union topsoil. Sci. Total. Environ. 2021, 769, 144755. [Google Scholar] [CrossRef]
- Louis, V.L.S.; Rudd, J.W.M.; Kelly, C.A.; Hall, B.D.; Rolfhus, K.R.; Scott, K.J.; Lindberg, S.E.; Dong, W. Importance of the Forest Canopy to Fluxes of Methyl Mercury and Total Mercury to Boreal Ecosystems. Environ. Sci. Technol. 2001, 35, 3089–3098. [Google Scholar] [CrossRef]
- Grigal, D.F. Mercury Sequestration in Forests and Peatlands. J. Environ. Qual. 2003, 32, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Rea, A.W.; Keeler, G.J.; Scherbatskoy, T. The deposition of mercury in throughfall and litterfall in the lake champlain watershed: A short-term study. Atmos. Environ. 1996, 30, 3257–3263. [Google Scholar] [CrossRef]
- Falandysz, J.; Frankowska, A.; Mazur, A. Mercury and its bioconcentration factors in King Bolete (Boletus edulis) Bull. Fr. J. Environ. Sci. Health Part A 2007, 42, 2089–2095. [Google Scholar] [CrossRef] [PubMed]
- Melgar, M.; Alonso, J.; García, M. Mercury in edible mushrooms and underlying soil: Bioconcentration factors and toxicological risk. Sci. Total. Environ. 2009, 407, 5328–5334. [Google Scholar] [CrossRef]
- Xu, J.; Kleja, D.B.; Biester, H.; Lagerkvist, A.; Kumpiene, J. Influence of particle size distribution, organic carbon, pH and chlorides on washing of mercury contaminated soil. Chemosphere 2014, 109, 99–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, E. The behavior of mercury in the soil with special emphasis on complexation and adsorption processes—A review of the literature. Water Air Soil Pollut. 1991, 56, 667–680. [Google Scholar] [CrossRef]
- Skyllberg, U.; Qian, J.; Frech, W.; Xia, K.; Bleam, W.F. Distribution of mercury, methyl mercury and organic sulphur species in soil, soil solution and stream of a boreal forest catchment. Biogeochemistry 2003, 64, 53–76. [Google Scholar] [CrossRef]
- Biester, H.; Müller, G.; Schöler, H. Binding and mobility of mercury in soils contaminated by emissions from chlor-alkali plants. Sci. Total. Environ. 2002, 284, 191–203. [Google Scholar] [CrossRef]
- Gómez-Armesto, A.; Martínez-Cortizas, A.; Ferro-Vázquez, C.; Méndez-López, M.; Arias-Estévez, M.; Nóvoa-Muñoz, J.C. Modelling Hg mobility in podzols: Role of soil components and environmental implications. Environ. Pollut. 2020, 260, 114040. [Google Scholar] [CrossRef]
- Augusto, L.; Bonnaud, P.; Ranger, J. Impact of tree species on forest soil acidification. For. Ecol. Manag. 1998, 105, 67–78. [Google Scholar] [CrossRef]
- Berg, B.; Erhagen, B.; Johansson, M.-B.; Nilsson, M.; Stendahl, J.; Trum, F.; Vesterdal, L. Manganese in the litter fall-forest floor continuum of boreal and temperate pine and spruce forest ecosystems—A review. For. Ecol. Manag. 2015, 358, 248–260. [Google Scholar] [CrossRef]
- Gruba, P.; Socha, J.; Pietrzykowski, M.; Pasichnyk, D. Tree species affects the concentration of total mercury (Hg) in forest soils: Evidence from a forest soil inventory in Poland. Sci. Total. Environ. 2019, 647, 141–148. [Google Scholar] [CrossRef]
- Babur, E.; Dindaroğlu, T.; Solaiman, Z.M.; Battaglia, M.L. Microbial respiration, microbial biomass and activity are highly sensitive to forest tree species and seasonal patterns in the Eastern Mediterranean Karst Ecosystems. Sci. Total. Environ. 2021, 775, 145868. [Google Scholar] [CrossRef]
- Demers, J.D.; Driscoll, C.T.; Fahey, T.J.; Yavitt, J.B. Mercury cycling in litter and soil in different forest types in the adirondack region, New York, USA. Ecol. Appl. 2007, 17, 1341–1351. [Google Scholar] [CrossRef]
- Richardson, J.B.; Friedland, A.J. Mercury in coniferous and deciduous upland forests in northern New England, USA: Implications of climate change. Biogeosciences 2015, 12, 6737–6749. [Google Scholar] [CrossRef] [Green Version]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.-J.; Nabuurs, G.-J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2012, 3, 203–207. [Google Scholar] [CrossRef]
- Vitasse, Y.; Bottero, A.; Cailleret, M.; Bigler, C.; Fonti, P.; Gessler, A.; Lévesque, M.; Rohner, B.; Weber, P.; Rigling, A.; et al. Contrasting resistance and resilience to extreme drought and late spring frost in five major European tree species. Glob. Chang. Biol. 2019, 25, 3781–3792. [Google Scholar] [CrossRef]
- Berg, B.; Erhagen, B.; Johansson, M.-B.; Vesterdal, L.; Faituri, M.; Sanborn, P.; Nilsson, M. Manganese dynamics in decomposing needle and leaf litter—A synthesis. Can. J. For. Res. 2013, 43, 1127–1136. [Google Scholar] [CrossRef]
- Rothe, A.; Huber, C.; Kreutzer, K.; Weis, W. Deposition and soil leaching in stands of Norway spruce and European Beech: Results from the Höglwald research in comparison with other European case studies. Plant Soil 2002, 240, 33–45. [Google Scholar] [CrossRef]
- De Schrijver, A.; Geudens, G.; Augusto, L.; Staelens, J.; Mertens, J.; Wuyts, K.; Gielis, L.; Verheyen, K. The effect of forest type on throughfall deposition and seepage flux: A review. Oecologia 2007, 153, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Novák, M.; Emmanuel, S.; Vile, M.A.; Erel, Y.; Véron, A.; Paces, T.; Wieder, R.K.; Vaněcek, M.; Štěpánová, M.; Břízová, E.; et al. Origin of Lead in Eight Central European Peat Bogs Determined from Isotope Ratios, Strengths, and Operation Times of Regional Pollution Sources. Environ. Sci. Technol. 2002, 37, 437–445. [Google Scholar] [CrossRef]
- CHMI Denní Data Dle Zákona 123/1998 Sb. Available online: https://www.chmi.cz/historicka-data/pocasi/denni-data/Denni-data-dle-z.-123-1998-Sb (accessed on 21 January 2021).
- Burt, R. Soil Survey Investigation Report No. 42; USDA: Lincoln, NE, USA, 2004. [Google Scholar]
- Courchesne, F.; Turmel, M.C. Extractable Al, Fe, Mn and Si. In Soil Sampling and Methods of Analysis; Carter, M.R., Gregorich, E.G., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 307–315. [Google Scholar]
- Sysalová, J.; Kučera, J.; Fikrle, M.; Drtinová, B. Determination of the total mercury in contaminated soils by direct solid sampling atomic absorption spectrometry using an AMA-254 device and radiochemical neutron activation analysis. Microchem. J. 2013, 110, 691–694. [Google Scholar] [CrossRef]
- Alonso, J.; Melgar, M.J. The Concentrations and Bioconcentration Factors of Copper and Zinc in Edible Mushrooms. Arch. Environ. Contam. Toxicol. 2003, 44, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Komárek, M.; Chrastný, V.; Štíchová, J. Metal/metalloid contamination and isotopic composition of lead in edible mushrooms and forest soils originating from a smelting area. Environ. Int. 2007, 33, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Nováčková, J.; Fiala, P.; Chrastný, V.; Svoboda, L.; Kalač, P. Contents of Mercury, Cadmium and Lead in Edible Mushrooms and in Uderlying Substrates from a Rural Area with an Occurrence of Serpentines and Amphiboles. Ekológia 2007, 26, 322–329. [Google Scholar]
- Sarikurkcu, C.; Popović-Djordjević, J.; Solak, M.H. Wild edible mushrooms from Mediterranean region: Metal concentrations and health risk assessment. Ecotoxicol. Environ. Saf. 2020, 190, 110058. [Google Scholar] [CrossRef]
- Kojta, A.K.; Jarzyńska, G.; Falandysz, J. Mineral composition and heavy metal accumulation capacity of Bay Bolete (Xerocomus badius) fruiting bodies collected near a former gold and copper mining area. J. Geochem. Explor. 2012, 121, 76–82. [Google Scholar] [CrossRef]
- USEPA Regional Screening Level (RSL) Resident Soil Table (TR=1E-06, HQ=1) May 2020 (Corrected). Available online: https://semspub.epa.gov/work/HQ/200047.pdf (accessed on 27 February 2021).
- Kalač, P. Trace element contents in European species of wild growing edible mushrooms: A review for the period 2000–2009. Food Chem. 2010, 122, 2–15. [Google Scholar] [CrossRef]
- IHIS CR European Health Interview Survey in CR—EHIS CR (Body Mass Index, Physical Activity, Consumption of Fruits and Vegetables); Institute of Health Information and Statistics of the Czech Republic: Prague, Czech, 2010.
- ČÚZK WMS Service Sub-Layer: Gdmr5g: Grayscale Hillshade. Available online: https://ags.cuzk.cz/arcgis2/services/dmr5g/ImageServer/WMSServer? (accessed on 21 January 2021).
- FMI WMS Service: UHUL OPRL Web Map Service. Available online: http://gp2.uhul.cz/wms_oprl/service.svc/get? (accessed on 15 March 2021).
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Usinglme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Yin, Y.; Allen, H.E.; Li, Y.; Huang, C.P.; Sanders, P.F. Adsorption of Mercury(II) by Soil: Effects of pH, Chloride, and Organic Matter. J. Environ. Qual. 1996, 25, 837–844. [Google Scholar] [CrossRef]
- Desai, M.V.M.; Wilken, R.-D. The role of humic substances in the aqueous mobilization of mercury from contaminated floodplain soils. Water Air Soil Pollut. 1996, 90, 507–520. [Google Scholar] [CrossRef]
- Schwesig, D.; Ilgen, G.; Matzner, E. Mercury and Methylmercury in Upland and Wetland Acid Forest Soils of a Watershed in NE-Bavaria, Germany. Water Air Soil Pollut. 1999, 113, 141–154. [Google Scholar] [CrossRef]
- Bäckström, M.; Dario, M.; Karlsson, S.; Allard, B. Effects of a fulvic acid on the adsorption of mercury and cadmium on goethite. Sci. Total. Environ. 2003, 304, 257–268. [Google Scholar] [CrossRef]
- Lockwood, R.A.; Chen, K.Y. Adsorption of mercury(II) by hydrous manganese oxides. Environ. Sci. Technol. 1973, 7, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.P.; Dubey, S.S.; Tiwari, D. Inorganic particulates in removal of heavy metal toxic ions. J. Colloid Interface Sci. 2004, 279, 61–67. [Google Scholar] [CrossRef]
- Yang, Y.-K.; Zhang, C.; Shi, X.-J.; Lin, T.; Wang, D.-Y. Effect of organic matter and pH on mercury release from soils. J. Environ. Sci. 2007, 19, 1349–1354. [Google Scholar] [CrossRef]


| Layer/Depth (cm) | C | N | S | C:N | pH | Sand | Silt | Clay | Alo | Feo |
|---|---|---|---|---|---|---|---|---|---|---|
| F+H | 0.240 | 0.243 | 0.327 | n.s. | −0.801 | NA | NA | NA | NA | NA |
| 0–2 | 0.574 | 0.643 | 0.618 | n.s. | −0.392 | n.s. | n.s. | n.s. | n.s. | n.s. |
| 2–10 | 0.706 | 0.833 | 0.649 | n.s. | n.s. | −0.512 | 0.516 | n.s. | 0.616 | 0.561 |
| 10–20 | 0.785 | 0.705 | 0.614 | n.s. | n.s. | −0.289 | 0.362 | n.s. | 0.625 | 0.644 |
| 20–30 | 0.762 | 0.670 | 0.715 | n.s. | −0.294 | −0.200 | 0.238 | n.s. | 0.671 | 0.487 |
| Layer/Depth | Full Model | Reduced Model | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Covariate | Coefficient | Variance | R2 | Variance | R2 | ||||||
| (cm) | (Name) | Estimate | SE | Fixed | Random | Residual | Marginal | Conditional | Random | Residual | Conditional |
| F+H | pH | −0.801 | 0.075 | 0.641 | 0.041 | 0.258 | 0.682 | 0.726 | 0.695 | 0.293 | 0.704 |
| 0–2 | S | 0.618 | 0.075 | 0.382 | 0.172 | 0.338 | 0428 | 0.621 | 0.405 | 0.583 | 0.410 |
| 2–10 | N | 0.833 | 0.074 | 0.694 | 0.117 | 0.266 | 0.645 | 0.753 | 0.321 | 0.664 | 0.326 |
| 10–20 | C | 0.785 | 0.066 | 0.616 | 0.027 | 0.289 | 0.661 | 0.690 | 0.467 | 0.521 | 0.473 |
| 20–30 | C | 0.762 | 0.068 | 0.581 | 0.063 | 0.224 | 0.669 | 0.742 | 0.596 | 0.393 | 0.603 |
| Mushroom Species | Norway Spruce | European Beech | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Hg | BCF | HRI | Hg | BCF | HRI | ||||
| F+H | 0–2 cm | F+H | 0–2 cm | ||||||
| Xerocomellus chrysenteron (Bull.) Šutara (n = 6 + 6) | Average | 0.36a | 0.78b | 1.47a | 0.47a | 0.35a | 1.31a | 1.41a | 0.46a |
| S.D. | 0.07 | 0.17 | 0.52 | 0.09 | 0.06 | 0.40 | 0.48 | 0.07 | |
| Imleria badia (Fr.) Vizzini (n = 6 + 4) | Average | 0.54 | 1.12 | 1.54 | 0.71 | 0.29 | 0.97 | 1.03 | 0.38 |
| S.D. | 0.88 | 1.76 | 2.22 | 1.15 | 0.14 | 0.45 | 0.54 | 0.18 | |
| Xerocomellus pruinatus (Fr. & Hök) Šutara (n = 3 + 2) | Average | 0.38 | 0.79 | 1.68 | 0.50 | 0.35 | 1.39 | 1.44 | 0.46 |
| S.D. | 0.08 | 0.20 | 0.78 | 0.11 | 0.02 | 0.12 | 0.05 | 0.02 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pecina, V.; Valtera, M.; Trávníčková, G.; Komendová, R.; Novotný, R.; Brtnický, M.; Juřička, D. Vertical Distribution of Mercury in Forest Soils and Its Transfer to Edible Mushrooms in Relation to Tree Species. Forests 2021, 12, 539. https://doi.org/10.3390/f12050539
Pecina V, Valtera M, Trávníčková G, Komendová R, Novotný R, Brtnický M, Juřička D. Vertical Distribution of Mercury in Forest Soils and Its Transfer to Edible Mushrooms in Relation to Tree Species. Forests. 2021; 12(5):539. https://doi.org/10.3390/f12050539
Chicago/Turabian StylePecina, Václav, Martin Valtera, Gabriela Trávníčková, Renata Komendová, Radek Novotný, Martin Brtnický, and David Juřička. 2021. "Vertical Distribution of Mercury in Forest Soils and Its Transfer to Edible Mushrooms in Relation to Tree Species" Forests 12, no. 5: 539. https://doi.org/10.3390/f12050539
APA StylePecina, V., Valtera, M., Trávníčková, G., Komendová, R., Novotný, R., Brtnický, M., & Juřička, D. (2021). Vertical Distribution of Mercury in Forest Soils and Its Transfer to Edible Mushrooms in Relation to Tree Species. Forests, 12(5), 539. https://doi.org/10.3390/f12050539