
Mid-Scale Drivers of Variability in Dry Mixed-Conifer Forests of the Mogollon Rim, Arizona


Abstract
:1. Introduction
2. Materials and Methods
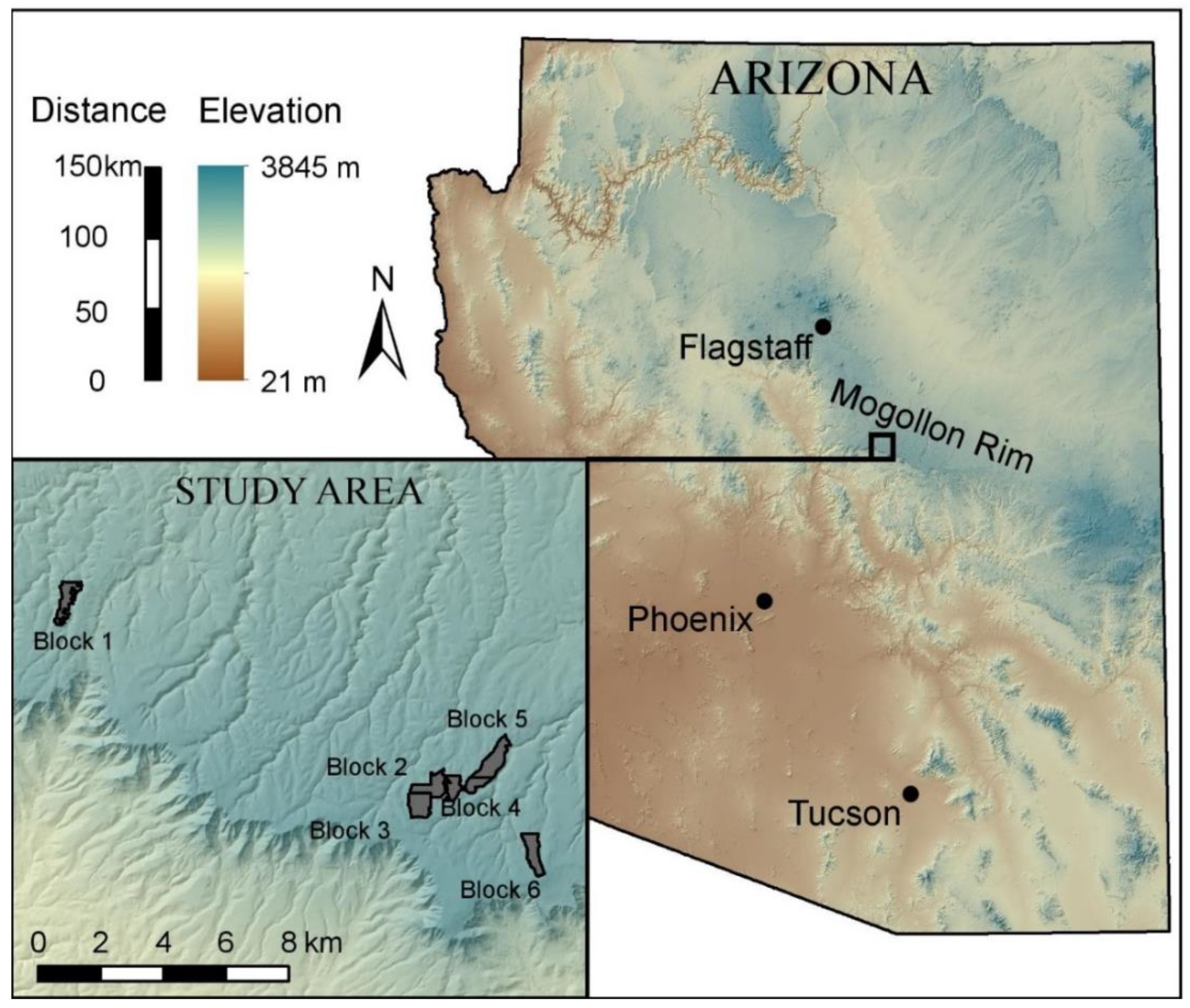
2.1. Study Design
2.2. Determining Historical Variability
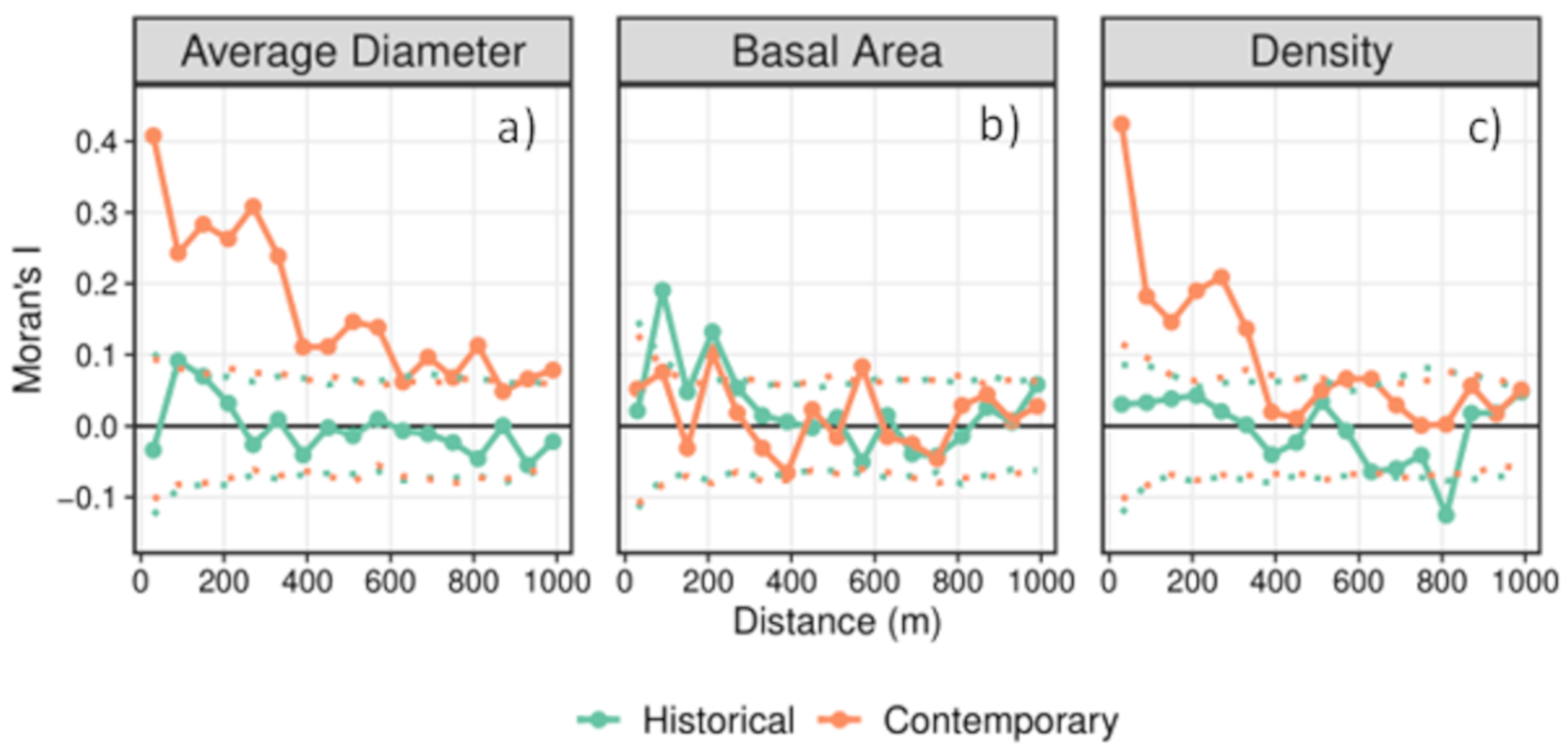
2.3. Measuring Spatial Variability
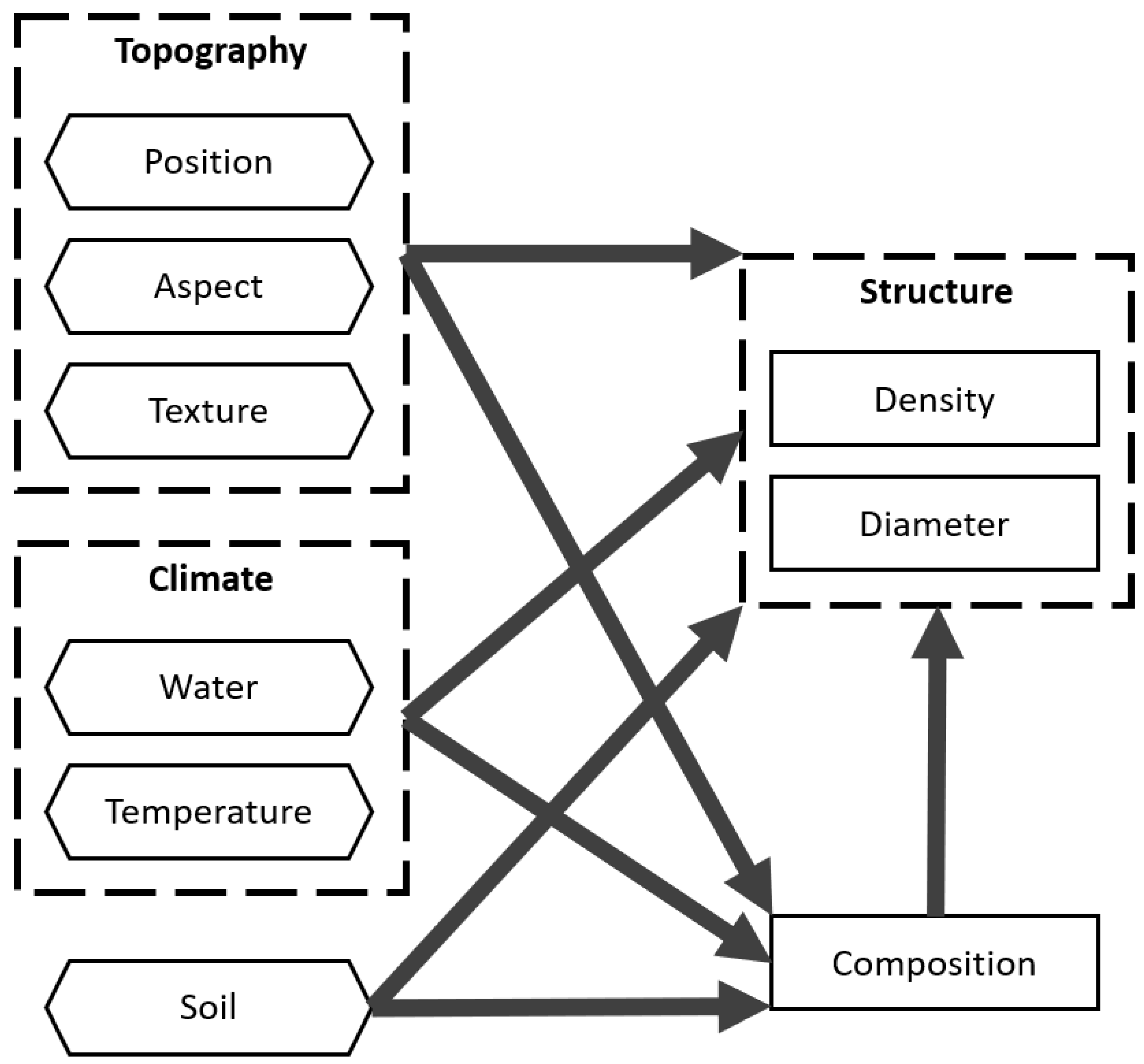
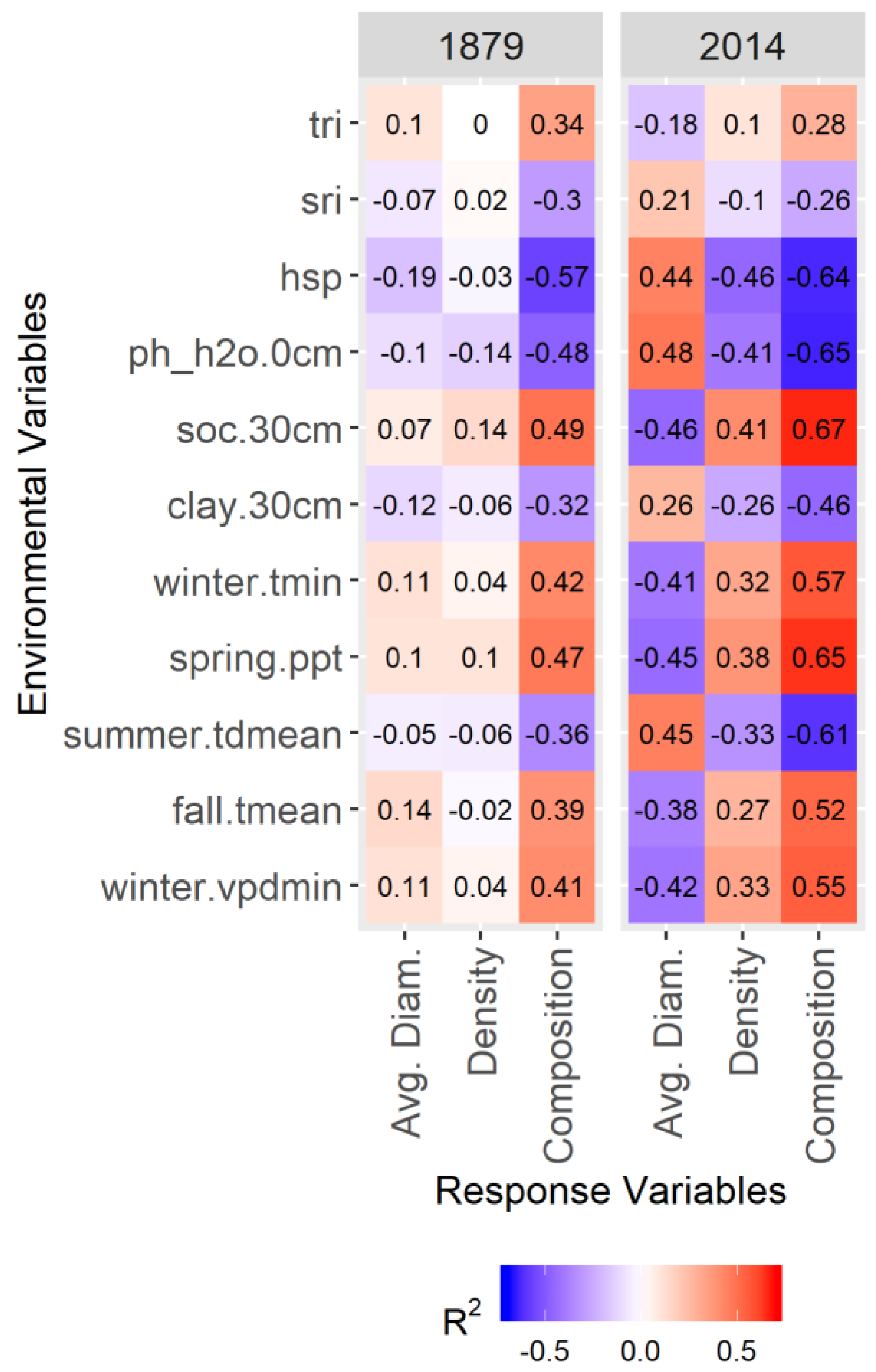
2.4. Identifying Drivers of Variability
3. Results
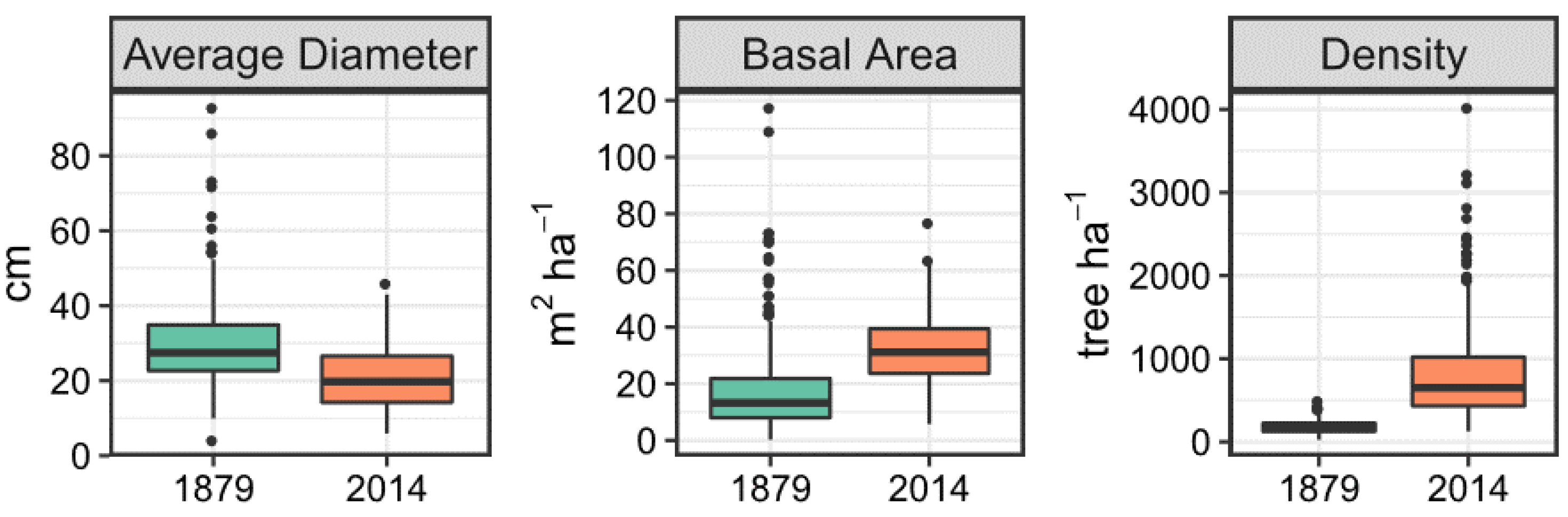
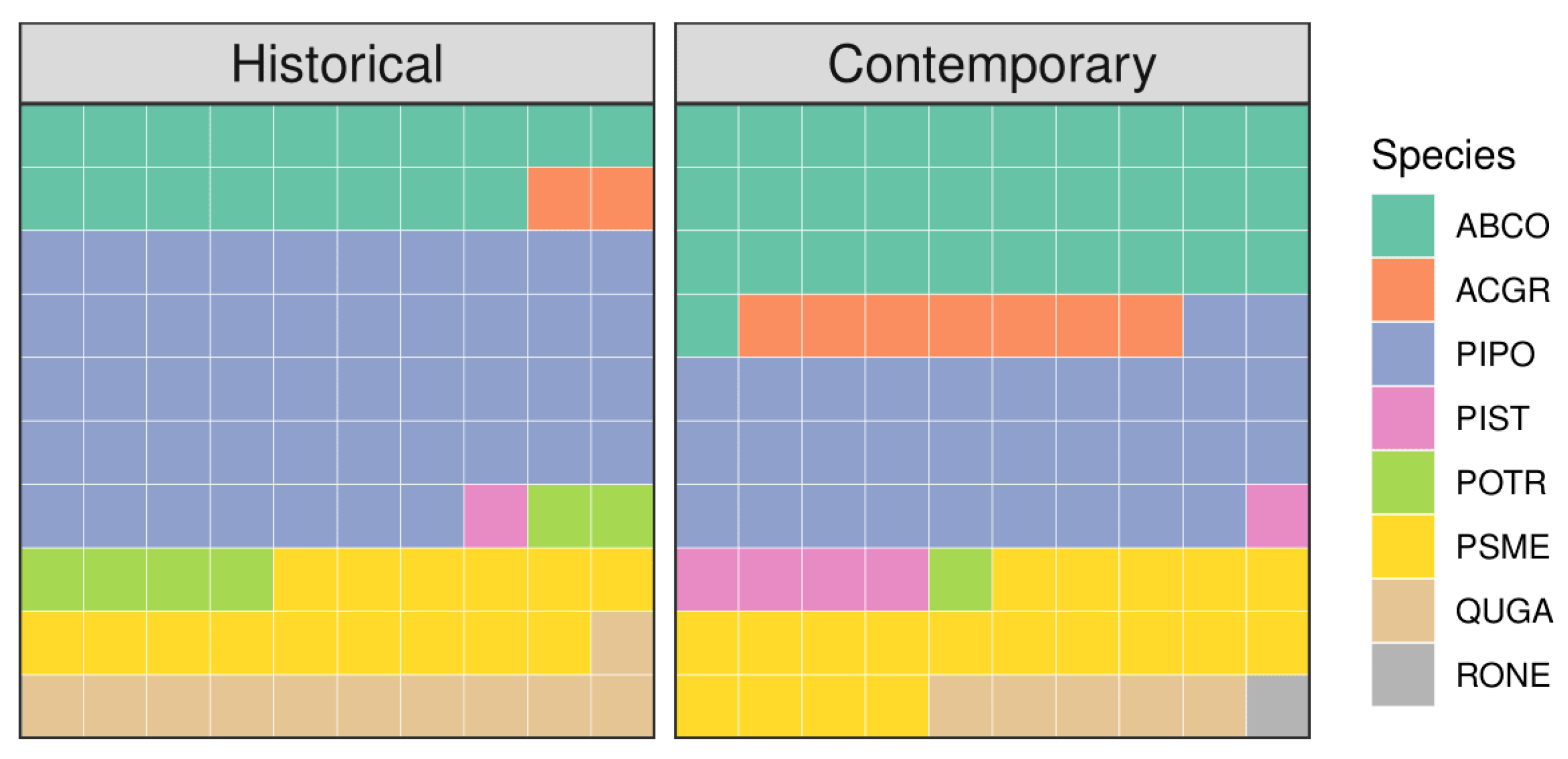
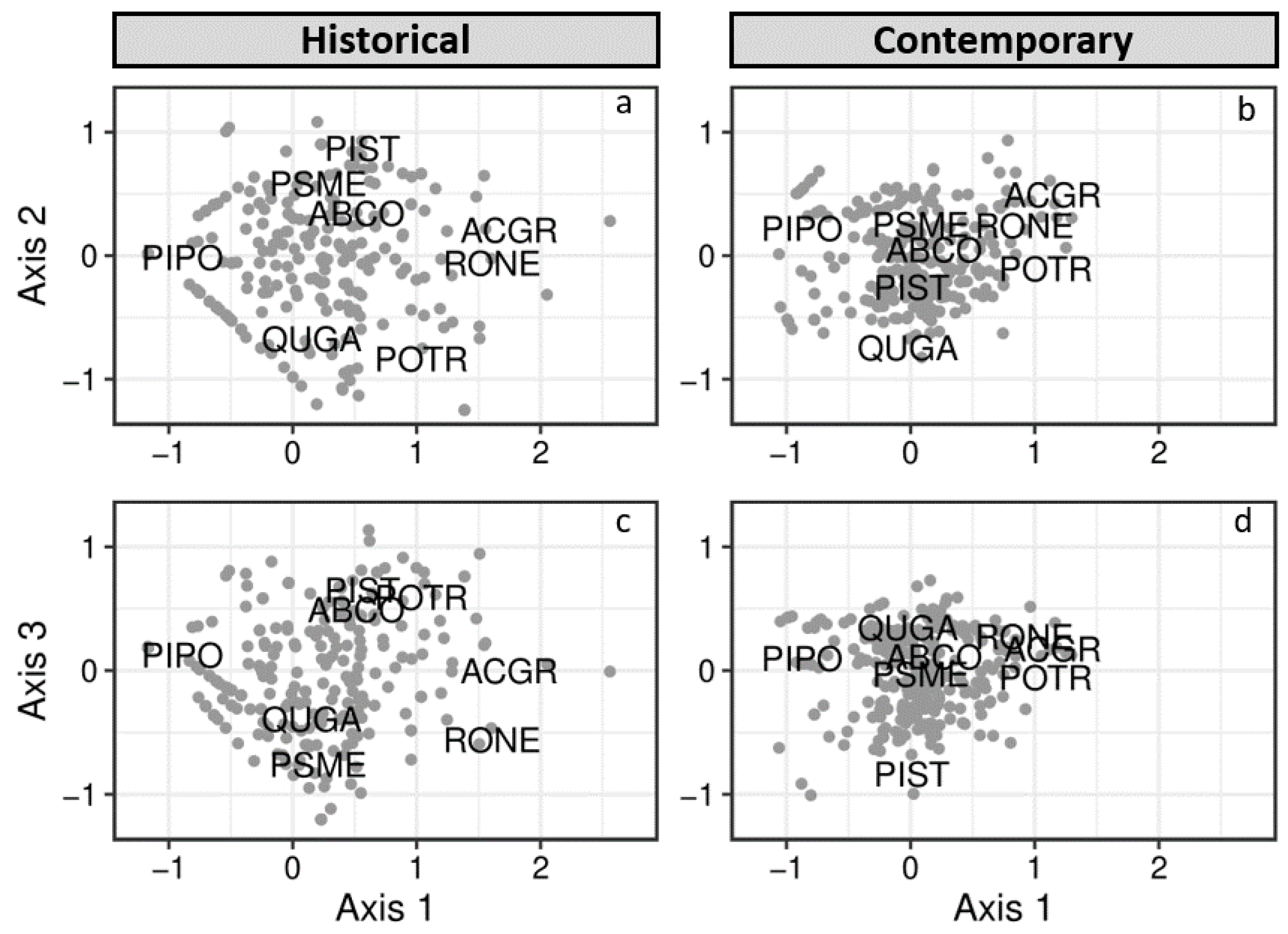
3.1. Historical and Contemporary Conditions
3.2. Spatial Variability
3.3. Drivers of Variability
4. Discussion
4.1. Historical Range of Variability
4.2. Changes to Spatial Patterns
4.3. Drivers of Variability
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Variable | Mean | Min | Max | SD | Description | Units | Source |
|---|---|---|---|---|---|---|---|---|
| Climate: Temperature | Annual tmax | 15.18 | 14.58 | 15.92 | 0.32 | Annual average of daily maximum temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Annual tmax | 15.55 | 14.95 | 16.31 | 0.33 | Annual average of daily maximum temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Spring tmax | 13.91 | 13.28 | 14.65 | 0.33 | Spring average of daily maximum temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Spring tmax | 14.44 | 13.80 | 15.22 | 0.34 | Spring average of daily maximum temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Summer tmax | 24.67 | 23.98 | 25.48 | 0.36 | Summer average of daily maximum temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Summer tmax | 25.44 | 24.75 | 26.27 | 0.36 | Summer average of daily maximum temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Fall tmax | 15.97 | 15.43 | 16.69 | 0.31 | Fall average of daily maximum temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Fall tmax | 16.27 | 15.72 | 17.01 | 0.31 | Fall average of daily maximum temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Winter tmax | 6.17 | 5.63 | 6.86 | 0.31 | Winter average of daily maximum temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Winter tmax | 6.07 | 5.54 | 6.76 | 0.30 | Winter average of daily maximum temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Annual tmean | 8.96 | 8.85 | 9.05 | 0.03 | Annual average of daily average temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Annual tmean | 9.31 | 9.20 | 9.40 | 0.04 | Annual average of daily average temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Spring tmean | 7.35 | 7.23 | 7.50 | 0.05 | Spring average of daily average temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Spring tmean | 7.84 | 7.73 | 8.00 | 0.05 | Spring average of daily average temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Summer tmean | 18.06 | 17.94 | 18.14 | 0.04 | Summer average of daily average temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Summer tmean | 18.53 | 18.40 | 18.63 | 0.05 | Summer average of daily average temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Fall tmean | 9.82 | 9.68 | 9.91 | 0.06 | Fall average of daily average temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Fall tmean | 10.16 | 9.99 | 10.29 | 0.08 | Fall average of daily average temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Winter tmean | 0.60 | 0.45 | 0.76 | 0.05 | Winter average of daily average temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Winter tmean | 0.71 | 0.63 | 0.84 | 0.04 | Winter average of daily average temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Annual tmin | 2.77 | 1.96 | 3.25 | 0.32 | Annual average of daily minimum temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Annual tmin | 3.07 | 2.25 | 3.61 | 0.33 | Annual average of daily minimum temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Spring tmin | 0.79 | 0.11 | 1.21 | 0.26 | Spring average of daily minimum temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Spring tmin | 1.24 | 0.57 | 1.68 | 0.26 | Spring average of daily minimum temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Summer tmin | 11.45 | 10.61 | 12.05 | 0.35 | Summer average of daily minimum temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Summer tmin | 11.62 | 10.79 | 12.26 | 0.36 | Summer average of daily minimum temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Fall tmin | 3.67 | 2.67 | 4.29 | 0.40 | Fall average of daily minimum temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Fall tmin | 4.04 | 3.00 | 4.76 | 0.44 | Fall average of daily minimum temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Temperature | Winter tmin | −4.98 | −5.70 | −4.69 | 0.26 | Winter average of daily minimum temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Temperature | Winter tmin | −4.64 | −5.34 | −4.25 | 0.27 | Winter average of daily minimum temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Water | Annual ppt | 921 | 760 | 975 | 71 | Annual total precipitation (1895 to 1924) | millimeters | PRISM |
| Climate: Water | Annual ppt | 887 | 752 | 932 | 59 | Annual total precipitation (1981 to 2010) | millimeters | PRISM |
| Climate: Water | Spring ppt | 169 | 131 | 187 | 17 | Spring total precipitation (1895 to 1924) | millimeters | PRISM |
| Climate: Water | Spring ppt | 169 | 136 | 184 | 15 | Spring total precipitation (1981 to 2010) | millimeters | PRISM |
| Climate: Water | Summer ppt | 267 | 203 | 286 | 29 | Summer total precipitation (1895 to 1924) | millimeters | PRISM |
| Climate: Water | Summer ppt | 242 | 182 | 259 | 27 | Summer total precipitation (1981 to 2010) | millimeters | PRISM |
| Climate: Water | Fall ppt | 195 | 179 | 200 | 7 | Fall total precipitation (1895 to 1924) | millimeters | PRISM |
| Climate: Water | Fall ppt | 190 | 177 | 194 | 5 | Fall total precipitation (1981 to 2010) | millimeters | PRISM |
| Climate: Water | Winter ppt | 289 | 246 | 316 | 19 | Winter total precipitation (1895 to 1924) | millimeters | PRISM |
| Climate: Water | Winter ppt | 286 | 255 | 306 | 14 | Winter total precipitation (1981 to 2010) | millimeters | PRISM |
| Climate: Water | Annual tdmean | −3.95 | −4.02 | −3.88 | 0.03 | Annual average of daily dewpoint temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Water | Annual tdmean | −2.29 | −2.38 | −2.21 | 0.04 | Annual average of daily dewpoint temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Water | Spring tdmean | −7.40 | −7.48 | −7.32 | 0.04 | Spring average of daily dewpoint temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Water | Spring tdmean | −5.45 | −5.54 | −5.30 | 0.06 | Spring average of daily dewpoint temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Water | Summer tdmean | 3.02 | 2.86 | 3.16 | 0.07 | Summer average of daily dewpoint temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Water | Summer tdmean | 4.41 | 4.28 | 4.53 | 0.07 | Summer average of daily dewpoint temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Water | Fall tdmean | −2.59 | −2.67 | −2.50 | 0.04 | Fall average of daily dewpoint temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Water | Fall tdmean | −0.78 | −0.89 | −0.65 | 0.05 | Fall average of daily dewpoint temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Water | Winter tdmean | −8.82 | −8.89 | −8.75 | 0.02 | Winter average of daily dewpoint temperature (1895 to 1924) | degrees Celsius | PRISM |
| Climate: Water | Winter tdmean | −7.36 | −7.41 | −7.26 | 0.03 | Winter average of daily dewpoint temperature (1981 to 2010) | degrees Celsius | PRISM |
| Climate: Water | Annual vpdmax | 14.65 | 13.94 | 15.51 | 0.37 | Annual average of daily maximum vapor pressure deficit (1895 to 1924) | hectopascals | PRISM |
| Climate: Water | Annual vpdmax | 14.94 | 14.19 | 15.75 | 0.35 | Annual average of daily maximum vapor pressure deficit (1981 to 2010) | hectopascals | PRISM |
| Climate: Water | Spring vpdmax | 13.51 | 12.87 | 14.24 | 0.32 | Spring average of daily maximum vapor pressure deficit (1895 to 1924) | hectopascals | PRISM |
| Climate: Water | Spring vpdmax | 13.73 | 13.11 | 14.40 | 0.29 | Spring average of daily maximum vapor pressure deficit (1981 to 2010) | hectopascals | PRISM |
| Climate: Water | Summer vpdmax | 24.22 | 23.03 | 25.61 | 0.61 | Summer average of daily maximum vapor pressure deficit (1895 to 1924) | hectopascals | PRISM |
| Climate: Water | Summer vpdmax | 25.03 | 23.56 | 26.49 | 0.66 | Summer average of daily maximum vapor pressure deficit (1981 to 2010) | hectopascals | PRISM |
| Climate: Water | Fall vpdmax | 14.35 | 13.72 | 15.21 | 0.35 | Fall average of daily maximum vapor pressure deficit (1895 to 1924) | hectopascals | PRISM |
| Climate: Water | Fall vpdmax | 14.38 | 13.73 | 15.16 | 0.32 | Fall average of daily maximum vapor pressure deficit (1981 to 2010) | hectopascals | PRISM |
| Climate: Water | Winter vpdmax | 6.53 | 6.15 | 6.99 | 0.20 | Winter average of daily maximum vapor pressure deficit (1895 to 1924) | hectopascals | PRISM |
| Climate: Water | Winter vpdmax | 6.61 | 6.36 | 6.93 | 0.13 | Winter average of daily maximum vapor pressure deficit (1981 to 2010) | hectopascals | PRISM |
| Climate: Water | Annual vpdmin | 3.26 | 2.84 | 3.56 | 0.18 | Annual average of daily minimum vapor pressure deficit (1895 to 1924) | hectopascals | PRISM |
| Climate: Water | Annual vpdmin | 3.09 | 2.92 | 3.27 | 0.09 | Annual average of daily minimum vapor pressure deficit (1981 to 2010) | hectopascals | PRISM |
| Climate: Water | Spring vpdmin | 3.10 | 2.78 | 3.32 | 0.13 | Spring average of daily minimum vapor pressure deficit (1895 to 1924) | hectopascals | PRISM |
| Climate: Water | Spring vpdmin | 3.15 | 2.97 | 3.30 | 0.07 | Spring average of daily minimum vapor pressure deficit (1981 to 2010) | hectopascals | PRISM |
| Climate: Water | Summer vpdmin | 5.30 | 4.61 | 5.84 | 0.31 | Summer average of daily minimum vapor pressure deficit (1895 to 1924) | hectopascals | PRISM |
| Climate: Water | Summer vpdmin | 5.13 | 4.88 | 5.46 | 0.15 | Summer average of daily minimum vapor pressure deficit (1981 to 2010) | hectopascals | PRISM |
| Climate: Water | Fall vpdmin | 3.16 | 2.66 | 3.53 | 0.22 | Fall average of daily minimum vapor pressure deficit (1895 to 1924) | hectopascals | PRISM |
| Climate: Water | Fall vpdmin | 2.86 | 2.64 | 3.11 | 0.12 | Fall average of daily minimum vapor pressure deficit (1981 to 2010) | hectopascals | PRISM |
| Climate: Water | Winter vpdmin | 1.48 | 1.30 | 1.55 | 0.06 | Winter average of daily minimum vapor pressure deficit (1895 to 1924) | hectopascals | PRISM |
| Climate: Water | Winter vpdmin | 1.20 | 1.15 | 1.23 | 0.02 | Winter average of daily minimum vapor pressure deficit (1981 to 2010) | hectopascals | PRISM |
| Soil | BD 0 cm | 666 | 527 | 948 | 67 | Bulk density of the fine earth fraction (<2 mm) at 0 cm soil depth | grams per cubic centimeter | Soil Grids |
| Soil | BD 5 cm | 891 | 766 | 1002 | 51 | Bulk density of the fine earth fraction (<2 mm) at 5 cm soil depth | grams per cubic centimeter | Soil Grids |
| Soil | BD 15 cm | 1131 | 1023 | 1212 | 36 | Bulk density of the fine earth fraction (<2 mm) at 15 cm soil depth | grams per cubic centimeter | Soil Grids |
| Soil | BD 30 cm | 1228 | 1162 | 1287 | 25 | Bulk density of the fine earth fraction (<2 mm) at 30 cm soil depth | grams per cubic centimeter | Soil Grids |
| Soil | BD 60 cm | 1336 | 1219 | 1418 | 44 | Bulk density of the fine earth fraction (<2 mm) at 60 cm soil depth | grams per cubic centimeter | Soil Grids |
| Soil | BD 100 cm | 1447 | 1364 | 1504 | 25 | Bulk density of the fine earth fraction (<2 mm) at 100 cm soil depth | grams per cubic centimeter | Soil Grids |
| Soil | BD 200 cm | 1439 | 1362 | 1507 | 30 | Bulk density of the fine earth fraction (<2 mm) at 200 cm soil depth | grams per cubic centimeter | Soil Grids |
| Soil | Clay 0 cm | 16 | 11 | 21 | 2 | Percent clay at 0 cm soil depth | percent by weight | Soil Grids |
| Soil | Clay 5 cm | 16 | 11 | 21 | 2 | Percent clay at 5 cm soil depth | percent by weight | Soil Grids |
| Soil | Clay 15 cm | 17 | 13 | 21 | 2 | Percent clay at 15 cm soil depth | percent by weight | Soil Grids |
| Soil | Clay 30 cm | 24 | 16 | 37 | 5 | Percent clay at 30 cm soil depth | percent by weight | Soil Grids |
| Soil | Clay 60 cm | 36 | 24 | 50 | 6 | Percent clay at 60 cm soil depth | percent by weight | Soil Grids |
| Soil | Clay 100 cm | 37 | 25 | 49 | 6 | Percent clay at 100 cm soil depth | percent by weight | Soil Grids |
| Soil | Clay 200 cm | 36 | 24 | 50 | 6 | Percent clay at 200 cm soil depth | percent by weight | Soil Grids |
| Soil | N Total 0 cm | 70 | 48 | 81 | 5 | Total Nitrogen at 0 cm soil depth | percent by weight | Soil Grids |
| Soil | N Total 5 cm | 37 | 29 | 43 | 3 | Total Nitrogen at 5 cm soil depth | percent by weight | Soil Grids |
| Soil | N Total 15 cm | 21 | 16 | 25 | 2 | Total Nitrogen at 15 cm soil depth | percent by weight | Soil Grids |
| Soil | N Total 30 cm | 13 | 8 | 18 | 2 | Total Nitrogen at 30 cm soil depth | percent by weight | Soil Grids |
| Soil | N Total 60 cm | 10 | 6 | 15 | 2 | Total Nitrogen at 60 cm soil depth | percent by weight | Soil Grids |
| Soil | N Total 100 cm | 9 | 4 | 14 | 2 | Total Nitrogen at 100 cm soil depth | percent by weight | Soil Grids |
| Soil | N Total 200 cm | 11 | 5 | 15 | 2 | Total Nitrogen at 200 cm soil depth | percent by weight | Soil Grids |
| Soil | pH 0 cm | 58.3 | 55.1 | 62.7 | 1.7 | pH in 1:1 soil–water solution at 0 cm soil depth | pH | Soil Grids |
| Soil | pH 5 cm | 56.8 | 53.7 | 61.0 | 1.6 | pH in 1:1 soil–water solution at 5 cm soil depth | pH | Soil Grids |
| Soil | pH 15 cm | 57.0 | 54.5 | 61.0 | 1.5 | pH in 1:1 soil–water solution at 15 cm soil depth | pH | Soil Grids |
| Soil | pH 30 cm | 57.0 | 55.0 | 60.3 | 1.3 | pH in 1:1 soil–water solution at 30 cm soil depth | pH | Soil Grids |
| Soil | pH 60 cm | 57.1 | 55.2 | 60.6 | 1.3 | pH in 1:1 soil–water solution at 60 cm soil depth | pH | Soil Grids |
| Soil | pH 100 cm | 57.2 | 55.1 | 61.6 | 1.5 | pH in 1:1 soil–water solution at 100 cm soil depth | pH | Soil Grids |
| Soil | pH 200 cm | 57.4 | 55.1 | 61.9 | 1.6 | pH in 1:1 soil–water solution at 200 cm soil depth | pH | Soil Grids |
| Soil | Sand 0 cm | 27 | 18 | 36 | 4 | Percent sand at 0 cm soil depth | percent by weight | Soil Grids |
| Soil | Sand 5 cm | 28 | 19 | 36 | 4 | Percent sand at 5 cm soil depth | percent by weight | Soil Grids |
| Soil | Sand 15 cm | 27 | 18 | 36 | 4 | Percent sand at 15 cm soil depth | percent by weight | Soil Grids |
| Soil | Sand 30 cm | 27 | 20 | 35 | 4 | Percent sand at 30 cm soil depth | percent by weight | Soil Grids |
| Soil | Sand 60 cm | 28 | 20 | 37 | 4 | Percent sand at 60 cm soil depth | percent by weight | Soil Grids |
| Soil | Sand 100 cm | 31 | 22 | 40 | 4 | Percent sand at 100 cm soil depth | percent by weight | Soil Grids |
| Soil | Sand 200 cm | 32 | 23 | 40 | 4 | Percent sand at 200 cm soil depth | percent by weight | Soil Grids |
| Soil | SOC 0 cm | 294 | 186 | 338 | 23 | Soil organic Carbon at 0 cm soil depth | percent by weight | Soil Grids |
| Soil | SOC 5 cm | 72 | 44 | 96 | 14 | Soil organic Carbon at 5 cm soil depth | percent by weight | Soil Grids |
| Soil | SOC 15 cm | 28 | 18 | 37 | 6 | Soil organic Carbon at 15 cm soil depth | percent by weight | Soil Grids |
| Soil | SOC 30 cm | 17 | 8 | 25 | 4 | Soil organic Carbon at 30 cm soil depth | percent by weight | Soil Grids |
| Soil | SOC 60 cm | 10 | 5 | 14 | 2 | Soil organic Carbon at 60 cm soil depth | percent by weight | Soil Grids |
| Soil | SOC 100 cm | 8 | 4 | 13 | 2 | Soil organic Carbon at 100 cm soil depth | percent by weight | Soil Grids |
| Soil | SOC 200 cm | 9 | 3 | 16 | 3 | Soil organic Carbon at 200 cm soil depth | percent by weight | Soil Grids |
| Topography: Aspect | Beer’s Aspect | 1.44 | 0.00 | 2.00 | 0.56 | Cosine transformed aspect | NA | DEM |
| Topography: Aspect | Heat Load Index (HLI) | 0.83 | 0.71 | 0.97 | 0.05 | Slope-aspect transformation | NA | DEM |
| Topography: Aspect | Solar Radiation Index (SRI) | 1,715,499 | 1,593,948 | 1,810,763 | 41,603 | Amount of incoming solar insolation | Watt hours per square meter | DEM |
| Topography: Position | Elevation | 2332 | 2223 | 2399 | 41 | Elevation above sea level | meters | DEM |
| Topography: Position | Hierarchical Slope Position (HSP) | 3295 | −10,371 | 14,386 | 5249 | Multi-scalar measure of topographic exposure | NA | DEM |
| Topography: Position | Topographic Position Index (TPI) | 0.55 | −11.53 | 9.75 | 3.61 | Local measure of topographic exposure | NA | DEM |
| Topography: Texture | Roughness | 17.1 | 1.4 | 31.2 | 5.8 | Maximum elevational difference | meters | DEM |
| Topography: Texture | Slope | 5.5 | 0.0 | 10.5 | 2.3 | Steepness of terrain | degrees | DEM |
| Topography: Texture | Terrain Ruggedness Index (TRI) | 19.3 | 2.0 | 42.0 | 7.2 | Average of elevational differences | meters | DEM |
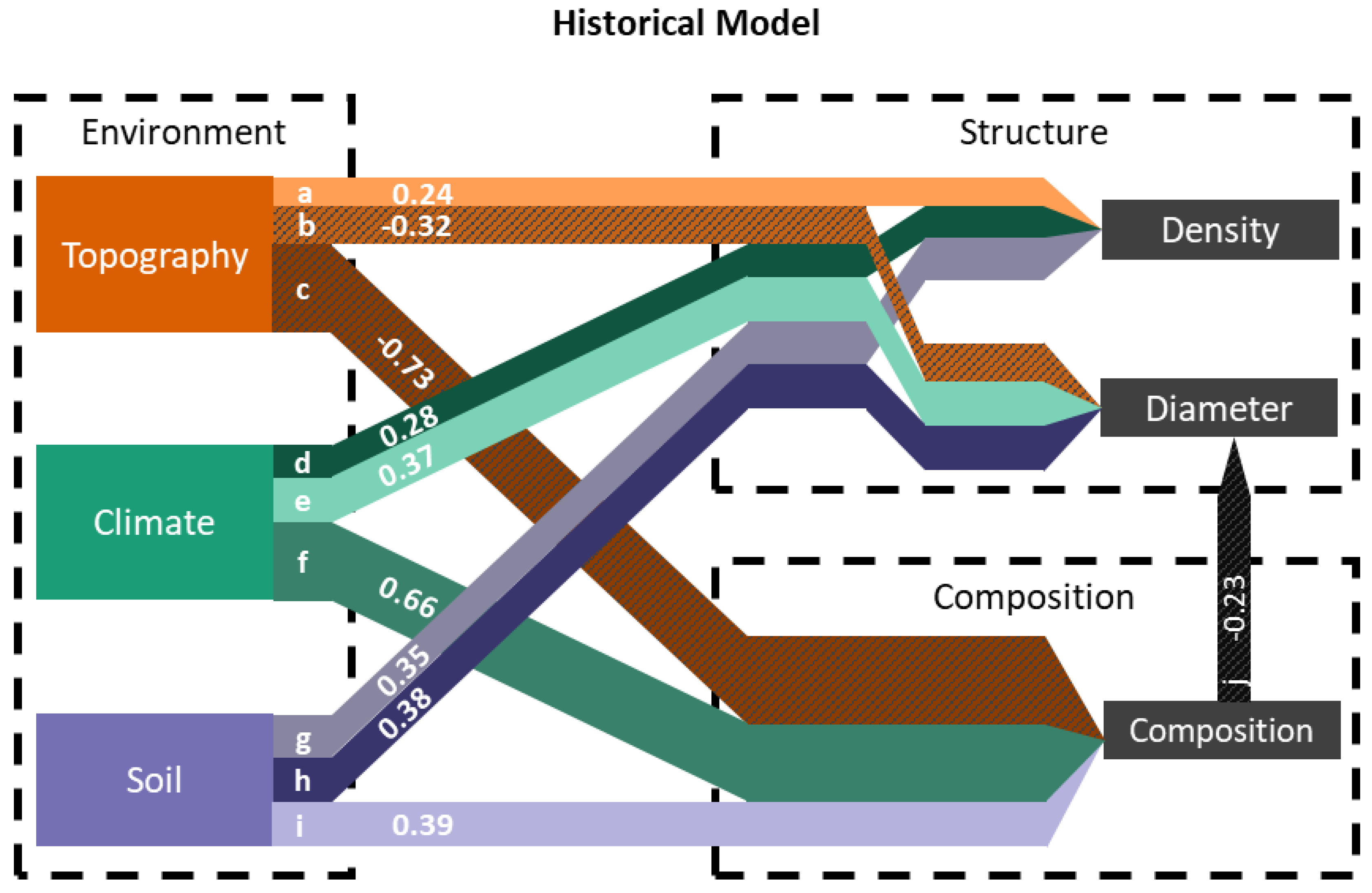
| Diagram Key | Pathway | Model | Coefficient | Components | |
|---|---|---|---|---|---|
| From | To | ||||
| a | Position | Density | Historical | 0.245 * | HSP |
| Contemporary | −0.358 * | HSP | |||
| b | Position | Diameter | Historical | −0.324 * | HSP |
| Contemporary | 0.321 * | HSP | |||
| c | Position | Composition | Historical | −0.371 * | HSP |
| Contemporary | −0.405 * | HSP | |||
| NA | Texture | Density | Historical | −0.065 | TRI |
| NA | Contemporary | −0.095 | TRI | ||
| NA | Texture | Diameter | Historical | 0.053 | TRI |
| NA | Contemporary | −0.020 | TRI | ||
| c | Texture | Composition | Historical | 0.126 * | TRI |
| NA | Contemporary | 0.068 | TRI | ||
| NA | Aspect | Density | Historical | 0.008 | SRI |
| NA | Contemporary | 0.010 | SRI | ||
| NA | Aspect | Diameter | Historical | −0.124 | SRI |
| NA | Contemporary | 0.072 | SRI | ||
| c | Aspect | Composition | Historical | −0.233 * | SRI |
| Contemporary | −0.184 * | SRI | |||
| d | Climate | Density | Historical | 0.276 * | Winter Tmin (−0.491); Winter Tmax (0.657); Fall Tmean (0.473); Spring Precip (3.546); Winter VPDmin (−3.462) |
| Contemporary | 0.212 * | Winter Tmin (−2.566); Winter Tmax (−3.615); Spring Precip (−1.742); Summer Tdmean (−0.035) | |||
| e | Climate | Diameter | Historical | 0.370 * | Winter Tmin (−2.052); Winter Tmax (−0.228); Fall Tmean (0.27); Spring Precip (0.773); Winter VPDmin (1.821) |
| Contemporary | 0.212 * | Winter Tmin (2.423); Winter Tmax (3.568); Spring Precip (2.202); Summer Tdmean (1.224) | |||
| f | Climate | Composition | Historical | 0.657 * | Winter Tmin (6.969); Winter Tmax (1.124); Fall Tmean (−0.769); Spring Precip (2.375); Winter VPDmin (−6.687) |
| Contemporary | 0.792 * | Winter Tmin (−0.855); Winter Tmax (0.185); Spring Precip (1.512); Summer Tdmean (−0.486) | |||
| g | Soil | Density | Historical | 0.352 * | Clay.30 cm (0.207); ph.0 cm (−0.726); SOC.30 cm (0.437) |
| Contemporary | 0.203 * | ph.0 cm (−0.793); SOC.30 cm (0.233) | |||
| h | Soil | Diameter | Historical | 0.383 * | Clay.30 cm (−0.482); pH.0 cm (0.441); SOC.30 cm (−0.863) |
| Contemporary | 0.259 * | pH.0 cm (0.723); SOC.30 cm (−0.309) | |||
| i | Soil | Composition | Historical | 0.389 * | Clay.30 cm (0.26); pH.0 cm (0.924); SOC.30 cm (0.116) |
| Contemporary | 0.345 * | pH.0 cm (0.661); SOC.30 cm (−0.373) | |||
| j | Composition | Diameter | Historical | −0.228 * | Axis 1 |
| NA | Contemporary | −0.058 | Axis 1 | ||
| NA | Composition | Density | Historical | 0.104 | Axis 1 |
| k | Contemporary | 0.255 * | Axis 1 | ||
Appendix B

References
- Dieterich, J.H. Fire History of Southwestern Mixed Conifer: A Case Study. For. Ecol. Manag. 1983, 6, 13–31. [Google Scholar] [CrossRef]
- Wan, H.Y.; McGarigal, K.; Ganey, J.L.; Lauret, V.; Timm, B.C.; Cushman, S.A. Meta-replication Reveals Nonstationarity in Multi-scale Habitat Selection of Mexican Spotted Owl. Condor 2017, 119, 641–658. [Google Scholar] [CrossRef]
- O’Donnell, F.C.; Flatley, W.T.; Springer, A.E.; Fulé, P.Z. Forest Restoration as a Strategy to Mitigate Climate Impacts on Wildfire, Vegetation, and Water in Semiarid Forests. Ecol. Appl. 2018, 28, 1459–1472. [Google Scholar] [CrossRef]
- Kalies, E.L.; Yocom Kent, L.L. Tamm Review: Are Fuel Treatments Effective at Achieving Ecological and Social Objectives? A Systematic Review. For. Ecol. Manag. 2016, 376, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Sánchez Meador, A.; Springer, J.D.; Huffman, D.W.; Bowker, M.A.; Crouse, J.E. Soil Functional Responses to Ecological Restoration Treatments in Frequent-fire Forests of the Western United States: A Systematic Review. Restor. Ecol. 2017, 25, 497–508. [Google Scholar] [CrossRef]
- Bahre, C.J. Late 19th Century Human Impacts on the Woodlands and Forests of Southeastern Arizona’s Sky Islands. Desert Plants 1998, 14, 8–21. [Google Scholar]
- Cooper, C.F. Changes in Vegetation, Structure, and Growth of South-western Pine Forests since White Settlement. Ecol. Monogr. 1960, 30, 129–164. [Google Scholar] [CrossRef]
- Savage, M.; Swetnam, T.W. Early 19th-Century Fire Decline Following Sheep Pasturing in a Navajo Ponderosa Pine Forest. Ecology 1990, 71, 2374–2378. [Google Scholar] [CrossRef]
- Reynolds, R.T.; Sánchez Meador, A.J.; Youtz, J.A.; Nicolet, T.; Matonis, M.S.; Jackson, P.L.; DeLorenzo, D.G.; Graves, A.D. Restoring Composition and Structure in Southwestern Frequent-Fire Forests: A Science-Based Framework for Improving Ecosystem Resiliency; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2013; p. 76.
- Bryant, T.; Waring, K.; Sánchez Meador, A.; Bradford, J.B. A Framework for Quantifying Resilience to Forest Disturbance. Front. For. Glob. Chang. 2019, 2, 56. [Google Scholar] [CrossRef]
- Society for Ecological Restoration International Science & Policy Working Group. The SER International Primer on Ecological Restoration; Society for Ecological Restoration International: Tucson, AZ, USA, 2004; p. 15. [Google Scholar]
- Fulé, P.Z.; Korb, J.E.; Wu, R. Changes in Forest Structure of a Mixed Conifer Forest, Southwestern Colorado, USA. For. Ecol. Manag. 2009, 258, 1200–1210. [Google Scholar] [CrossRef]
- Huffman, D.W.; Zegler, T.J.; Fulé, P.Z. Fire History of a Mixed Conifer Forest on the Mogollon Rim, Northern Arizona, USA. Int. J. Wildland Fire 2015, 24, 680–689. [Google Scholar] [CrossRef]
- Strahan, R.T.; Sánchez Meador, A.J.; Huffman, D.W.; Laughlin, D.C. Shifts in Community-level Traits and Functional Diversity in a Mixed Conifer Forest: A Legacy of Land-use Change. J. Appl. Ecol. 2016, 53, 1755–1765. [Google Scholar] [CrossRef] [Green Version]
- Landres, P.B.; Morgan, P.; Swanson, F.J. Overview of the Use of Natural Variability Concepts in Managing Ecological Systems. Ecol. Appl. 1999, 9, 1179. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate Change and Forests of the Future: Managing in the Face of Uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef]
- Walker, R.B.; Coop, J.D.; Parks, S.A.; Trader, L. Fire Regimes Approaching Historic Norms Reduce Wildfire-facilitated Conversion from Forest to Non-forest. Ecosphere 2018, 9. [Google Scholar] [CrossRef]
- Romme, W.; Floyd, M.; Hanna, D.; Bartlett, E.; Crist, M.; Green, D.; Grissino-Mayer, H.; Lindsey, J.; Mcgarigal, K.; Redders, J. Historical Range of Variability and Current Landscape Condition Analysis: South Central Highlands Section, Southwestern Colorado & Northwestern New Mexico; Colorado Forest Restoration Institute, Colorado State University: Fort Collins, CO, USA, 2009. [Google Scholar]
- Rodman, K.C.; Sánchez Meador, A.J.; Huffman, D.W.; Waring, K.M. Reference Conditions and Historical Fine-Scale Spatial Dynamics in a Dry Mixed-Conifer Forest, Arizona, USA. For. Sci. 2016, 62, 268–280. [Google Scholar] [CrossRef]
- Fulé, P.Z.; Crouse, J.E.; Heinlein, T.A.; Moore, M.M.; Covington, W.W.; Verkamp, G. Mixed-severity Fire Regime in a High-elevation Forest of Grand Canyon, Arizona, USA. Landsc. Ecol. 2003, 18, 465–486. [Google Scholar] [CrossRef]
- Cocke, A.E.; Fulé, P.Z.; Crouse, J.E. Forest Change on a Steep Mountain Gradient after Extended Fire Exclusion: San Francisco Peaks, Arizona, USA. J. Appl. Ecol. 2005, 42, 814–823. [Google Scholar] [CrossRef]
- Heinlein, T.A.; Moore, M.M.; Fulé, P.Z.; Covington, W.W. Fire History and Stand Structure of Two Ponderosa Pine-mixed Conifer Sites: San Francisco Peaks, Arizona, USA. Int. J. Wildland Fire 2005, 14, 307. [Google Scholar] [CrossRef]
- Rodman, K.C.; Sánchez Meador, A.J.; Moore, M.M.; Huffman, D.W. Reference Conditions are Influenced by the Physical Template and Vary by Forest Type: A Synthesis of Pinus ponderosa-Dominated Sites in the Southwestern United States. For. Ecol. Manag. 2017, 404, 316–329. [Google Scholar] [CrossRef]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and Earlier Spring Increase Western U.S. Forest Wildfire Activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef] [Green Version]
- Westerling, A.L.R. Increasing Western US Forest Wildfire Activity: Sensitivity to Changes in the Timing of Spring. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef]
- Margolis, E.Q.; Balmat, J. Fire History and Fire–climate Relationships along a Fire Regime Gradient in the Santa Fe Municipal Watershed, NM, USA. For. Ecol. Manag. 2009, 258, 2416–2430. [Google Scholar] [CrossRef]
- Urban, D.L.; Miller, C.; Halpin, P.N.; Stephenson, N.L. Forest Gradient Response in Sierran Landscapes: The Physical Template. Landscape Ecol. 2000, 15, 603–620. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Fulé, P.Z.; Huffman, D.W.; Crouse, J.; Laliberté, E. Climatic Constraints on Trait-based Forest Assembly. J. Ecol. 2011, 99, 1489–1499. [Google Scholar] [CrossRef]
- Swetnam, T.; Baisan, C. Historical Fire Regime Patterns in the Southwestern United States Since AD 1700. In Fire Effects in Southwestern Forests. In Proceedings of the Second La Mesa Fire Symposium, Los Alamos, NM, USA, 29–31 March 1994; U.S. Department of Agriculture, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1996; pp. 11–32. [Google Scholar]
- Brown, P.M.; Wu, R. Climate and Disturbance Forcing of Episodic Tree Recruitment in a Southwestern Ponderosa Pine Landscape. Ecology 2005, 86, 3030–3038. [Google Scholar] [CrossRef]
- League, K.; Veblen, T. Climatic Variability and Episodic Pinus ponderosa Establishment along the Forest-grassland Ecotones of Colorado. For. Ecol. Manag. 2006, 228, 98–107. [Google Scholar] [CrossRef]
- Puhlick, J.J.; Laughlin, D.C.; Moore, M.M. Factors Influencing Ponderosa Pine Regeneration in the Southwestern USA. For. Ecol. Manag. 2012, 264, 10–19. [Google Scholar] [CrossRef]
- Stephens, S.L.; Stevens, J.T.; Collins, B.M.; York, R.A.; Lydersen, J.M. Historical and Modern Landscape Forest Structure in Fir (Abies)-dominated Mixed Conifer Forests in the Northern Sierra Nevada, USA. Fire Ecol. 2018, 14, 7. [Google Scholar] [CrossRef] [Green Version]
- Abella, S.R.; Covington, W.W. Vegetation–environment Relationships and Ecological Species Groups of an Arizona Pinus Ponderosa Landscape, USA. Plant Ecol. 2006, 185, 255–268. [Google Scholar] [CrossRef]
- Abella, S.R.; Denton, C.W. Spatial Variation in Reference Conditions: Historical Tree Density and Pattern on a Pinus ponderosa Landscape. Can. J. For. Res. 2009, 39, 2391–2403. [Google Scholar] [CrossRef]
- Kimsey, M.J.; Shaw, T.M.; Coleman, M.D. Site Sensitive Maximum Stand Density Index Models for Mixed Conifer Stands Across the Inland Northwest, USA. For. Ecol. Manag. 2019, 433, 396–404. [Google Scholar] [CrossRef]
- Larson, A.J.; Churchill, D. Tree Spatial Patterns in Fire-frequent Forests of Western North America, Including Mechanisms of Pattern Formation and Implications for Designing Fuel Reduction and Restoration Treatments. For. Ecol. Manag. 2012, 267, 74–92. [Google Scholar] [CrossRef]
- Larson, A.J.; Stover, K.C.; Keyes, C.R. Effects of Restoration Thinning on Spatial Heterogeneity in Mixed-conifer Forest. Can. J. For. Res. 2012. [Google Scholar] [CrossRef]
- Hendricks, J.D.; Plescia, J.B. A Review of the Regional Geophysics of the Arizona Transition Zone. J. Geophys. Res. 1991, 96, 12351–12373. [Google Scholar] [CrossRef]
- Four Forest Restoration Initiative. Available online: https://www.fs.usda.gov/4fri (accessed on 20 April 2020).
- Cragin Watershed Protection Project. Available online: www.fs.usda.gov/detail/coconino/home/?cid=stelprd3850490 (accessed on 20 April 2020).
- Roccaforte, J.P.; Huffman, D.W.; Fulé, P.Z.; Covington, W.W.; Chancellor, W.W.; Stoddard, M.T.; Crouse, J.E. Forest Structure and Fuels Dynamics Following Ponderosa Pine Restoration Treatments, White Mountains, Arizona, USA. For. Ecol. Manag. 2015, 337, 174–185. [Google Scholar] [CrossRef]
- Myers, C.A. Estimating Past Diameters of Ponderosa Pines in Arizona and New Mexico; Rocky Mountain Forest and Range Experiment Station, Forest Service: Fort Collins, CO, USA; U.S. Department of Agriculture: Washington, DC, USA, 1963.
- Parker, J.L.; Thomas, J.W. Wildlife Habitats in Managed Forests: The Blue Mountains of Oregon and Washington; US Department of Agriculture: Washington, DC, USA, 1979.
- Sánchez Meador, A.J.; (Northern Arizona University School of Forestry, Flagstaff, AZ, USA). Personal communication, 2020.
- Bakker, J.D.; Sanchez Meador, A.J.; Fulé, P.Z.; Huffman, D.W.; Moore, M.M. “Growing Trees Backwards”: Description of a Stand Reconstruction Model; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2008; pp. 136–147.
- Moore, M.M.; Huffman, D.W.; Fulé, P.Z.; Covington, W.W.; Crouse, J.E. Comparison of Historical and Contemporary Forest Structure and Composition on Permanent Plots in Southwestern Ponderosa Pine Forests. For. Sci. 2004, 50, 162–176. [Google Scholar] [CrossRef]
- Curtis, J.T.; McIntosh, R.P. An Upland Forest Continuum in the Prairie-Forest Border Region of Wisconsin. Ecology 1951, 32, 476–496. [Google Scholar] [CrossRef]
- Bivand, R.S.; Pebesma, E.; Gómez-Rubio, V. Applied Spatial Data Analysis with R, 2nd ed.; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-7618-4. [Google Scholar]
- Bivand, R.S.; Wong, D.W.S. Comparing Implementations of Global and Local Indicators of Spatial Association. TEST 2018, 27, 716–748. [Google Scholar] [CrossRef]
- Moran, P.A.P. Notes on Continuous Stochastic Phenomena. Biometrika 1950, 37, 17. [Google Scholar] [CrossRef]
- Cliff, A.D.; Ord, J.K. Spatial Autocorrelation; Pion: London, UK, 1973. [Google Scholar]
- Mast, J.N.; Wolf, J.J. Spatial Patch Patterns and Altered Forest Structure in Middle Elevation versus upper Ecotonal Mixed-conifer Forests, Grand Canyon National Park, Arizona, USA. For. Ecol. Manag. 2006. [Google Scholar] [CrossRef]
- Arbuckle, J.L. Amos; IBM SPSS: Chicago, IL, USA, 2014. [Google Scholar]
- Grace, J.B. Structural Equation Modeling and Natural Systems; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Grace, J.B.; Bollen, K.A. Representing General Theoretical Concepts in Structural Equation Models: The Role of Composite Variables. Environ. Ecol. Stat. 2008, 15, 191–213. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Grace, J.B. A Multivariate Model of Plant Species Richness in Forested Systems: Old-growth Montane Forests with a Long History of Fire. Oikos 2006, 114, 60–70. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Abella, S.R.; Covington, W.W.; Grace, J.B. Species Richness and Soil Properties in Pinus ponderosa Forests: A Structural Equation Modeling Analysis. J. Veg. Sci. 2007, 18, 231–242. [Google Scholar] [CrossRef]
- Huffman, D.W.; Laughlin, D.C.; Pearson, K.W.; Pandey, S. Effects of Vertebrate Herbivores and Shrub Characteristics on Arthropod Assemblages in a Northern Arizona Forest Ecosystem. For. Ecol. Manag. 2009, 258, 616–625. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Bowker, M.A.; Grace, J.B.; Powell, J.R. From Patterns to Causal Understanding: Structural Equation Modeling (SEM) in Soil Ecology. Pedobiologia 2015, 58, 65–72. [Google Scholar] [CrossRef] [Green Version]
- ArcGIS Desktop: Release 10; Environmental Systems Research Institute (ESRI): Redlands, CA, USA, 2011.
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling; R Package, University of California: Davis, CA, USA, 2019. [Google Scholar]
- Evans, J.S. Spatialeco; R Package, University of Wyoming: Laramie, WY, USA, 2019. [Google Scholar]
- Beers, T.W.; Dress, P.E.; Wensel, L.C. Notes and Observations: Aspect Transformation in Site Productivity Research. J. For. 1966, 64, 691–692. [Google Scholar] [CrossRef]
- McCune, B.; Keon, D. Equations for Potential Annual Direct Incident Radiation and Heat Load. J. Veg. Sci. 2002, 13, 603–606. [Google Scholar] [CrossRef]
- Rich, P.M.; Dubayah, R.; Hetrick, W.A.; Saving, S.C. Using Viewshed Models to Calculate Intercepted Solar Radiation: Applications in Ecology; American Society for Photogrammetry and Remote Sensing: Bethesda, MD, USA, 1994; pp. 524–529. [Google Scholar]
- Riley, S.J.; DeGloria, S.D.; Elliot, R. Index That Quantifies Topographic Heterogeneity. Intermt. J. Sci. 1999, 5, 23–27. [Google Scholar]
- PRISM Climate Group. Climate Data; Oregon State University: Corvallis, OR, USA, 2019. [Google Scholar]
- Web Soil Survey. Available online: Websoilsurvey.nrcs.usda.gov/ (accessed on 20 April 2020).
- Ramcharan, A.; Hengl, T.; Nauman, T.; Brungard, C.; Waltman, S.; Wills, S.; Thompson, J. Soil Property and Class Maps of the Conterminous United States at 100-Meter Spatial Resolution. Soil Sci. Soc. Am. J. 2018, 82, 186–201. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; R Package, University of Helsinki: Helsinki, Norway, 2019. [Google Scholar]
- Wassermann, T.; Stoddard, M.T.; Waltz, A.E.M. Working Paper 42: A Summary of the Natural Range of Variability for Southwestern Frequent-Fire Forests; Ecological Restoration Institute, Northern Arizona University: Flagstaff, AZ, USA, 2019; p. 11. [Google Scholar]
- Williams, M.A.; Baker, W.L. Spatially Extensive Reconstructions Show Variable-severity Fire and Heterogeneous Structure in Historical Western United States Dry Forests. Glob. Ecol. Biogeogr. 2012, 21, 1042–1052. [Google Scholar] [CrossRef]
- Brown, P.M.; Battaglia, M.A.; Fornwalt, P.J.; Gannon, B.; Huckaby, L.S.; Julian, C.; Cheng, A.S. Historical (1860) Forest Structure in Ponderosa pine Forests of the Northern Front Range, Colorado. Can. J. For. Res. 2015, 45, 1462–1473. [Google Scholar] [CrossRef] [Green Version]
- Battaglia, M.A.; Gannon, B.; Brown, P.M.; Fornwalt, P.J.; Cheng, A.S.; Huckaby, L.S. Changes in Forest Structure since 1860 in Ponderosa Pine Dominated Forests in the Colorado and Wyoming Front Range, USA. For. Ecol. Manag. 2018, 422, 147–160. [Google Scholar] [CrossRef] [Green Version]
- Lydersen, J.M.; North, M.P.; Knapp, E.E.; Collins, B.M. Quantifying Spatial Patterns of Tree Groups and Gaps in Mixed-conifer Forests: Reference Conditions and Long-term Changes Following Fire Suppression and Logging. For. Ecol. Manag. 2013, 304, 370–382. [Google Scholar] [CrossRef]
- Collins, B.M.; Lydersen, J.M.; Everett, R.G.; Fry, D.L.; Stephens, S.L. Novel Characterization of Landscape-level Variability in Historical Vegetation Structure. Ecol. Appl. 2015, 25, 1167–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, S.L.; Lydersen, J.M.; Collins, B.M.; Fry, D.L.; Meyer, M.D. Historical and Current Landscape-scale Ponderosa Pine and Mixed Conifer Forest Structure in the Southern Sierra Nevada. Ecosphere 2015, 6. [Google Scholar] [CrossRef]
- Hagmann, R.K.; Franklin, J.F.; Johnson, K.N. Historical Structure and Composition of Ponderosa Pine and Mixed-conifer Forests in South-central Oregon. For. Ecol. Manag. 2013, 304, 492–504. [Google Scholar] [CrossRef] [Green Version]
- Hagmann, R.K.; Franklin, J.F.; Johnson, K.N. Historical Conditions in Mixed-conifer Forests on the Eastern Slopes of the Northern Oregon Cascade Range, USA. For. Ecol. Manag. 2014, 330, 158–170. [Google Scholar] [CrossRef]
- Hagmann, R.K.; Johnson, D.L.; Johnson, K.N. Historical and Current Forest Conditions in the Range of the Northern Spotted Owl in South Central Oregon, USA. For. Ecol. Manag. 2017, 389, 374–385. [Google Scholar] [CrossRef]
- Binkley, D.; Romme, B.; Cheng, T. Historical Forest Structure on the Uncompahgre Plateau: Informing Restoration Prescriptions for Mountainside Stewardship; Colorado Forest Restoration Initiative, Colorado State University: Fort Collins, CO, USA, 2008; p. 26. [Google Scholar]
- Korb, J.E.; Fulé, P.Z.; Wu, R. Variability of Warm/Dry Mixed Conifer Forests in Southwestern Colorado, USA: Implications for Ecological Restoration. For. Ecol. Manag. 2013, 304, 182–191. [Google Scholar] [CrossRef]
- Mueller, S.E.; Thode, A.E.; Margolis, E.Q.; Yocom, L.L.; Young, J.D.; Iniguez, J.M. Climate Relationships with Increasing Wildfire in the Southwestern US from 1984 to 2015. For. Ecol. Manag. 2020, 460. [Google Scholar] [CrossRef]
- Meunier, J.; Romme, W.H.; Brown, P.M. Climate and Land-use Effects on Wildfire in Northern Mexico, 1650–2010. For. Ecol. Manag. 2014, 325, 49–59. [Google Scholar] [CrossRef]
- Swetnam, T.W.; Farella, J.; Roos, C.I.; Liebmann, M.J.; Falk, D.A.; Allen, C.D. Multiscale Perspectives of Fire, Climate and Humans in Western North America and the Jemez Mountains, USA. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150168. [Google Scholar] [CrossRef] [PubMed]
- Fulé, P.Z.; Laughlin, D.C. Wildland Fire Effects on Forest Structure over an Altitudinal Gradient, Grand Canyon National Park, USA. J. Appl. Ecol. 2006, 44, 136–146. [Google Scholar] [CrossRef]
- Huffman, D.W.; Sánchez Meador, A.J.; Stoddard, M.T.; Crouse, J.E.; Roccaforte, J.P. Efficacy of Resource Objective Wildfires for Restoration of Ponderosa Pine (Pinus ponderosa) Forests in Northern Arizona. For. Ecol. Manag. 2017, 389. [Google Scholar] [CrossRef] [Green Version]
- Huffman, D.W.; Crouse, J.E.; Sánchez Meador, A.J.; Springer, J.D.; Stoddard, M.T. Restoration Benefits of Re-entry with Resource Objective Wildfire on a Ponderosa Pine Landscape in Northern Arizona, USA. For. Ecol. Manag. 2018, 408. [Google Scholar] [CrossRef]
- Stoddard, M.T.; Fulé, P.Z.; Huffman, D.W.; Sánchez Meador, A.J.; Roccaforte, J.P. Ecosystem Management Applications of Resource Objective Wildfires in Forests of the Grand Canyon National Park, USA. Int. J. Wildland Fire 2020, 29, 190. [Google Scholar] [CrossRef]









| Model Fit | Response Variable r2 | |||||
|---|---|---|---|---|---|---|
| Model | Chi2 (p-Value) | RMSEA (p-Value) | AGFI | Density | Average Diameter | Composition |
| Historical | 0.58 (0.447) | <0.001 (0.581) | 0.968 | 0.088 | 0.101 | 0.483 |
| Contemporary | 0.42 (0.515) | <0.001 (0.638) | 0.980 | 0.296 | 0.317 | 0.632 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaquette, M.; Sánchez Meador, A.J.; Huffman, D.W.; Bowker, M.A. Mid-Scale Drivers of Variability in Dry Mixed-Conifer Forests of the Mogollon Rim, Arizona. Forests 2021, 12, 622. https://doi.org/10.3390/f12050622
Jaquette M, Sánchez Meador AJ, Huffman DW, Bowker MA. Mid-Scale Drivers of Variability in Dry Mixed-Conifer Forests of the Mogollon Rim, Arizona. Forests. 2021; 12(5):622. https://doi.org/10.3390/f12050622
Chicago/Turabian StyleJaquette, Matthew, Andrew J. Sánchez Meador, David W. Huffman, and Matthew A. Bowker. 2021. "Mid-Scale Drivers of Variability in Dry Mixed-Conifer Forests of the Mogollon Rim, Arizona" Forests 12, no. 5: 622. https://doi.org/10.3390/f12050622
APA StyleJaquette, M., Sánchez Meador, A. J., Huffman, D. W., & Bowker, M. A. (2021). Mid-Scale Drivers of Variability in Dry Mixed-Conifer Forests of the Mogollon Rim, Arizona. Forests, 12(5), 622. https://doi.org/10.3390/f12050622