
Biochemical and Gene Expression Analyses in Different Poplar Clones: The Selection Tools for Afforestation of Halomorphic Environments


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Biochemical Analyses
2.2.1. Extract Preparation for Biochemical Assays
2.2.2. Flavonoid Determination
2.2.3. Total Phenol Content (TPC) Assay
2.2.4. DPPH Assay
2.2.5. ABTS+ Assay
2.2.6. FRAP Assay
2.3. Biochemical Data Analysis
2.4. Gene Expression Analyses
2.4.1. RT-PCR and Dye-Based qPCR Analyses
2.4.2. Stress-Related Candidate Genes
2.4.3. Gene Expression Data Analyses
3. Results and Discussion
3.1. Biochemical Analyses
3.2. Comparative Genomic Studies
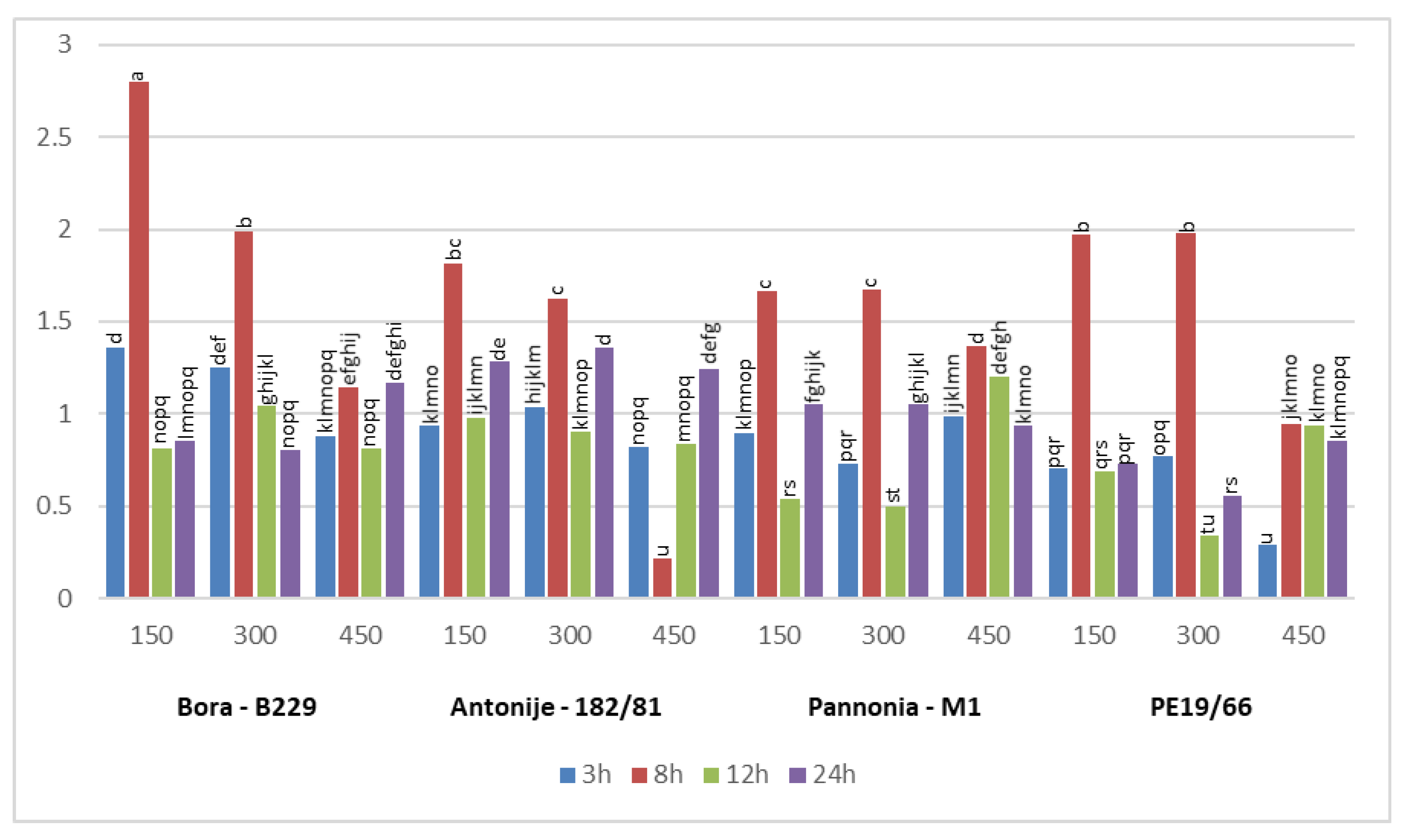
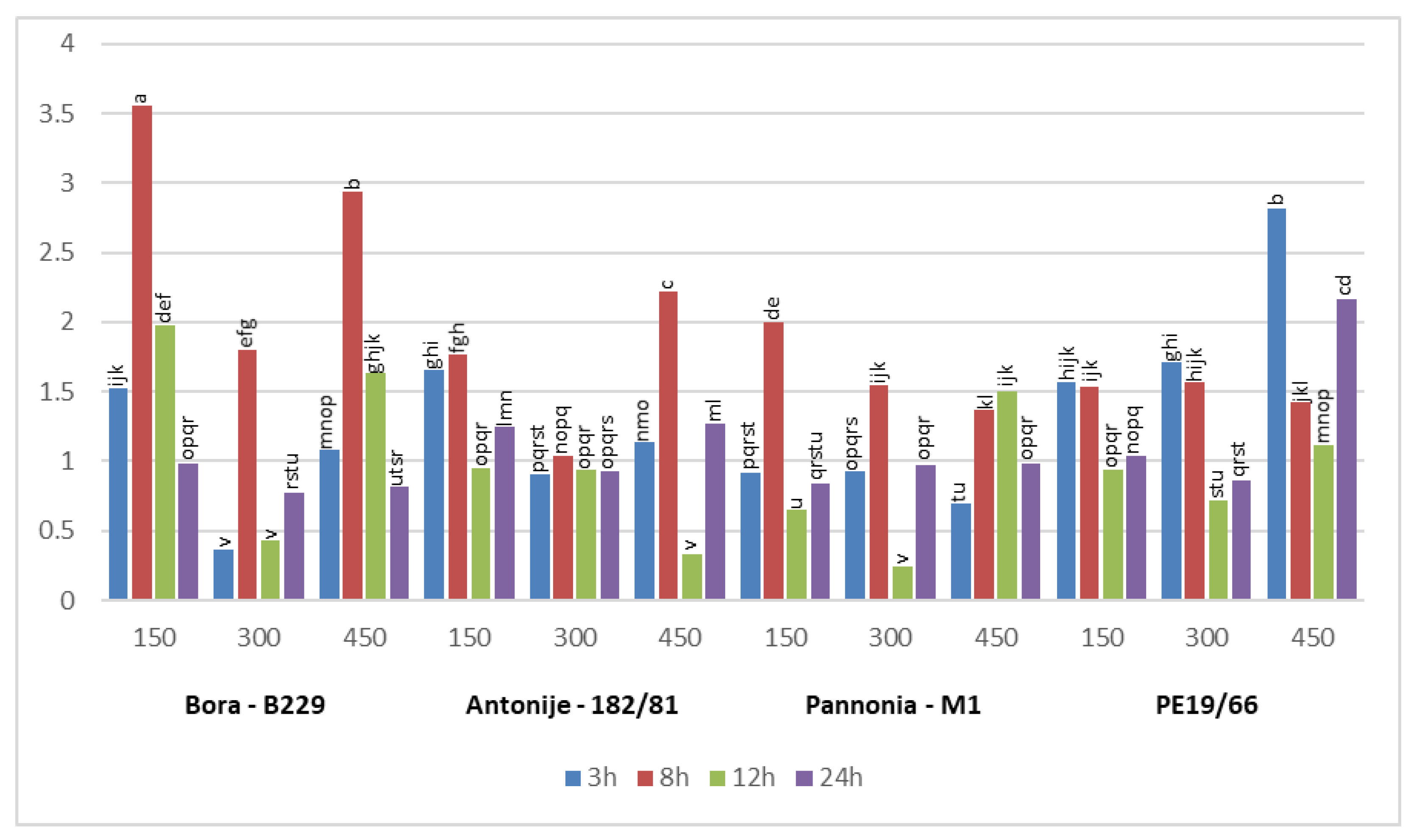
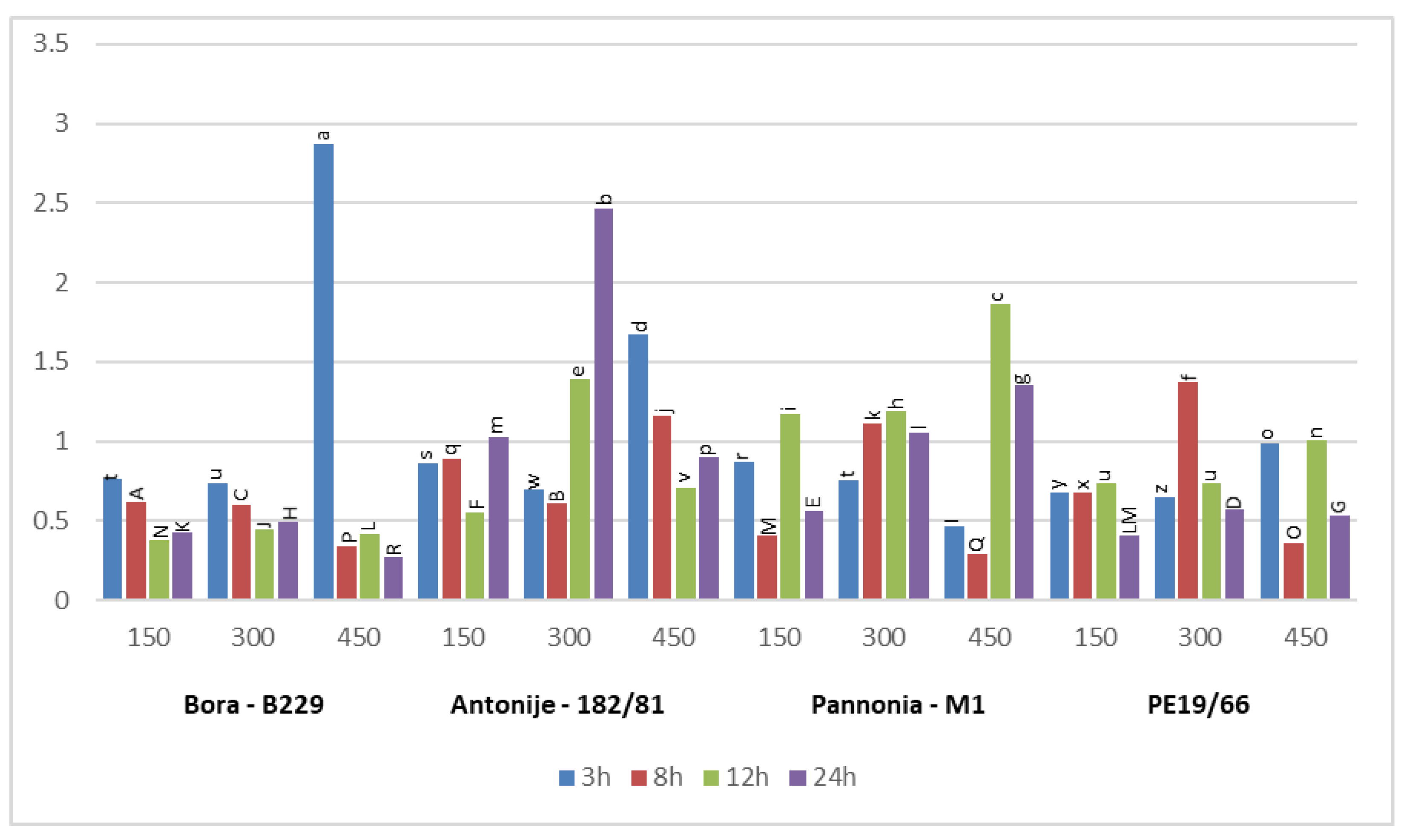
3.3. Differential Expression Pattern of Salt Stress-Related Candidate Genes
3.3.1. Salt Stress Responses of PtP5CS1
3.3.2. Salt Responses of PtSOS1
3.3.3. Salt Responses of PtDREB2
3.3.4. Salt Responses of PtGRAS16 and PtGRAS17
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shabala, S.; Bose, J.; Hedrich, R. Salt bladders: Do they matter? Trends Plant Sci. 2014, 9, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liu, C.; Tang, D.; Yan, L.; Wang, D.; Yang, Y.; Gui, J.; Zhao, X.-Y.; Li, L.; Tang, X.-D.; et al. The Receptor-like Cytoplasmic Kinase STRK1 Phosphorylates and Activates CatC, thereby Regulating H2O2 Homeostasis and Improving Salt Tolerance in Rice. Plant Cell 2018, 30, 1100–1118. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Polle, A. Salinity tolerance of Populus. Plant Biol. 2010, 12, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.C.; Marshal, C. The accumulation of zinc by subcellular fractions of roots of Agrostis tenuis Sibth. in relation to zinc tolerance. New Phytol. 1972, 71, 671–676. [Google Scholar] [CrossRef]
- Zorić, M.; Đukić, I.; Kljajić, L.; Karaklić, D.; Orlović, S. The possibilities for improvement of ecosystem services in Tara National Park. Topola 2019, 203, 53–63. [Google Scholar]
- Kesić, L.; Matović, B.; Stojnić, S.; Stjepanović, S.; Stojanović, D. Climate change as a factor reducing the growth of trees in the pure Norway spruce stand (Picea abies (L.) H. Karst.) in the national park “Kopaonik”. Topola 2016, 197/198, 25–34. [Google Scholar]
- Ivanišević, P.; Galić, Z.; Rončević, S.; Kovačević, B.; Marković, M. Significance of establishment of forest tree and shrub plantations for the stability and sustainable development of ecosystems in Vojvodina. Topola 2008, 181/182, 31–40. [Google Scholar]
- Ličina, V.; Nešić, L.; Belić, M.; Hadžić, V.; Sekulić, P.; Vasin, J.; Ninkov, J. The soils of Serbia and their degradation. Ratar. Povrt. 2011, 48, 285–290. [Google Scholar] [CrossRef]
- Wilkins, D.A. The measurement of tolerance to edaphic factors by means of root growth. New Phytol. 1978, 80, 623–633. [Google Scholar] [CrossRef]
- Ben Ahmed, C.; Ben Rouina, B.; Sensoy, S.; Boukhriss, M. Saline Water Irrigation Effects on Fruit Development, Quality, and Phenolic Composition of Virgin Olive Oils, Cv. Chemlali. J. Agric. Food Chem. 2009, 57, 2803–2811. [Google Scholar] [CrossRef]
- Petridis, A.; Therios, I.; Samouris, G.; Tananaki, C. Salinity-induced changes in phenolic compounds in leaves and roots of four olive cultivars (Olea europaea L.) and their relationship to antioxidant activity. Environ. Exp. Bot. 2012, 79, 37–43. [Google Scholar] [CrossRef]
- Taârit, M.B.; Msaada, K.; Hosni, K.; Marzouk, B. Physiological changes, phenolic content and antioxidant activity of Salvia officinalis L. grown under saline conditions. J. Sci. Food Agric. 2012, 92, 1614–1619. [Google Scholar] [CrossRef] [PubMed]
- Hura, T.; Hura, K.; Grzesiak, S. Contents of total phenolics and ferulic acid, and PAL activity during water potential changes in leaves of maize single-cross hybrids of different drought tolerance. J. Agron. Crop Sci. 2008, 194, 104–112. [Google Scholar] [CrossRef]
- Nguyen, K.K.; Cuellar, C.; Mavi, P.S.; LeDuc, D.; Bañuelos, G.; Sommerhalter, M. Two Poplar Hybrid Clones Differ in Phenolic Antioxidant Levels and Polyphenol Oxidase Activity in Response to High Salt and Boron Irrigation. J. Agric. Food Chem. 2018, 66, 7256–7264. [Google Scholar] [CrossRef] [PubMed]
- Popović, B.M.; Štajner, D.; Ždero-Pavlović, R.; Tumbas-Šaponjac, V.; Čanadanović-Brunet, J.; Orlović, S. Water stress induces changes in polyphenol profile and antioxidant capacity in poplar plants (Populus spp.). Plant Physiol. Biochem. 2016, 105, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Liu, S.; Lu, Z.; Pang, S.; Wang, L.; Wang, L.; Li, W. Gene Expression Profiles and Flavonoid Accumulation during Salt Stress in Ginkgo biloba Seedlings. Plants 2020, 9, 1162. [Google Scholar] [CrossRef]
- Valifard, M.; Mohsenzadeh, S.; Kholdebarin, B.; Rowshan, V. Effects of salt stress on volatile compounds, total phenolic content and antioxidant activities of Salvia mirzayanii. S. Afr. J. Bot. 2014, 93, 92–97. [Google Scholar] [CrossRef] [Green Version]
- Vuksanović, V.; Kovačević, B.; Kebert, M.; Milović, M.; Kesić, L.; Karaklić, V.; Orlović, S. In vitro modulation of antioxidant and physiological properties of white poplar induced by salinity. Bull. Fac. For. 2019, 120, 179–196. [Google Scholar] [CrossRef]
- Popović, B.M.; Štajner, D.; Ždero-Pavlović, R.; Tari, I.; Csiszár, J.; Gallé, Á.; Poór, P.; Galović, V.; Trudić, B.; Orlović, S. Biochemical response of hybrid black poplar tissue culture (Populus × canadensis) on water stress. J. Plant Res. 2017, 130, 559–570. [Google Scholar] [CrossRef] [Green Version]
- Niu, X.; Bressan, R.A.; Hasegawa, P.M.; Pardo, J.M. Ion Homeostasis in NaCl Stress Environments. Plant Physiol. 1995, 109, 735–742. [Google Scholar] [CrossRef] [Green Version]
- Štajner, D.; Orlović, S.; Popović, B.M.; Kebert, M.; Galić, Z. Screening of drought oxidative stress tolerance in Serbian melliferous plant species. Afr. J. Biotechnol. 2011, 10, 1609–1614. [Google Scholar] [CrossRef]
- Pessarakli, M.; Szabolcs, I. Soil salinity and sodicity as particular plant/crop stress factors. In Handbook of Plant and Crop Stress; Pessarakli, M., Ed.; Dekker: New York, NY, USA, 1999; pp. 1–16. ISBN 0824719484. [Google Scholar]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.-K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [Green Version]
- Tuskan, G.; DiFazio, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Jansson, S.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray ex Brayshaw). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef] [Green Version]
- Brosché, M.; Vinocur, B.; Alatalo, E.R.; Lamminmäki, A.; Teichmann, T.; Ottow, E.A.; Djilianov, D.; Afif, D.; Bogeat-Triboulot, M.B.; Altman, A.; et al. Gene expression and metabolite profiling of Populus euphratica growing in the Negev desert. Genome Biol. 2005, 6, R101. [Google Scholar] [CrossRef] [Green Version]
- Sairam, R.K.; Tyagi, A. Physiology and Molecular Biology of Salinity Stress Tolerance in Plants. Curr. Sci. 2004, 86, 407–421. [Google Scholar] [CrossRef] [Green Version]
- Giri, J. Glycinebetaine and abiotic stress tolerance in plants. Plant Signal. Behav. 2011, 6, 1746–1751. [Google Scholar] [CrossRef]
- Qureshi, M.; Abdin, M.; Ahmad, J.; Iqbal, M. Effect of long-term salinity on cellular antioxidants, compatible solute and fatty acid profile of sweet Annie (Artemisia annua L.). Phytochemistry 2013, 95, 215–223. [Google Scholar] [CrossRef]
- Kebert, M.; Rapparini, F.; Neri, L.; Bertazza, G.; Orlović, S.; Biondiet, S. Copper-induced responses in poplar clones are associated with genotype- and organ-specific changes in peroxidase activity and proline, polyamine, ABA, and IAA levels. J. Plant Growth Regul. 2017, 36, 131–147. [Google Scholar] [CrossRef]
- Ashrafa, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Székely, G.; Abrahám, E.; Cséplo, A.; Rigó, G.; Zsigmond, L.; Csiszár, J.; Ayaydin, F.; Strizhov, N.; Jásik, J.; Schmelzer, E.; et al. Duplicated P5CS genes of Arabidopsis play distinct roles in stress regulation and developmental control of proline biosynthesis. Plant J. 2008, 53, 11–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabro, G.; Kovács, I.; Pavet, V.; Szabados, L.; Alvarez, M.E. Proline accumulation and AtP5CS2 gene activation are induced by plant-pathogen incompatible interactions in Arabidopsis. Mol. Plant Microbe Interact. 2004, 17, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Gharsallah, C.; Fakhfakh, H.; Grubb, D.; Gorsane, F. Effect of salt stress on ion concentration, proline content, antioxidant enzyme activities and gene expression in tomato cultivars. AoB Plants 2016, 8, plw055. [Google Scholar] [CrossRef] [Green Version]
- Pottosin, I.; Velarde-Buendía, A.M.; Bose, J.; Zepeda-Jazo, I.; Shabala, S.; Dobrovinskaya, O. Cross-talk between reactive oxygen species and polyamines in regulation of ion transport across the plasma membrane: Implications for plant adaptive responses. J. Exp. Bot. 2014, 65, 1271–1283. [Google Scholar] [CrossRef] [Green Version]
- Kavi Kishor, P.B.; Sangam, S.; Amrutha, R.N.; Laxmi, P.S.; Naidu, K.R.; Rao, K.R.S.S.; Rao, S.; Theriappan, P.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr. Sci. 2005, 88, 424–438. [Google Scholar]
- Bray, E.A.; Bailey-Serres, J.; Weretilnyk, E. Responses to abiotic stress. In Biochemistry & Molecular Biology of Plants; Gruissem, W., Jones, R., Eds.; American Society of Plant Physiologists: Rockville, MA, USA, 2000; pp. 1158–1249. ISBN 0943088399. [Google Scholar]
- Cushman, J.C.; Bohnert, H.J. Genomic approaches to plant stress tolerance. Curr. Opin. Plant Biol. 2000, 3, 117–124. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When Defense Pathways Collide. The Response of Arabidopsis to a Combination of Drought and Heat Stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.S.; Zhang, J.; Li, M.-X. Metabolomics Analysis Reveals the Salt-Tolerant Mechanism in Glycine soja. J. Plant Growth Regul. 2017, 36, 460–471. [Google Scholar] [CrossRef]
- Rhodes, D.; Hanson, A.D. Quaternary ammonium and tertiary sulfonium compounds in higher-plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1993, 44, 357–384. [Google Scholar] [CrossRef]
- Seki, M.; Narusaka, M.; Ishida, J.; Nanjo, T.; Fujita, M.; Oono, Y.; Kamiya, A.; Nakajima, M.; Enju, A.; Sakurai, T.; et al. Monitoring the expression profiles of 7000 Arabidopsis genes under drought, cold and high salinity stresses using a full-length cDNA microarray. Plant J. 2002, 31, 279–292. [Google Scholar] [CrossRef]
- Wang, Q.; Guan, C.; Wang, P.; Ma, Q.; Bao, A.K.; Zhang, J.L.; Wang, S.M. The Effect of AtHKT1;1 or AtSOS1 Mutation on the Expressions of Na⁺ or K⁺ Transporter Genes and Ion Homeostasis in Arabidopsis thaliana under Salt Stress. Int. J. Mol. Sci. 2019, 20, 1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Q.-S.; Guo, Y.; Dietrich, M.A.; Schumaker, K.S.; Zhu, J.-K. Regulation of SOS1, a plasma membrane Na+/H+ exchanger in Arabidopsis thaliana, by SOS2 and SOS3. Proc. Natl. Acad. Sci. USA 2002, 99, 8436–8441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Ishitani, M.; Kim, C.; Zhu, J.-K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, H.; Shao, H.; Tang, X. Recent Advances in Utilizing Transcription Factors to Improve Plant Abiotic Stress Tolerance by Transgenic Technology. Front. Plant Sci. 2016, 7, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.S.; Liang, D.; Shuai, P.; Xia, X.L.; Yin, W.L. The salt- and drought-inducible poplar GRAS protein SCL7 confers salt and drought tolerance in Arabidopsis thaliana. J. Exp. Bot. 2010, 61, 4011–4019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubouzet, J.G.; Sakuma, Y.; Ito, Y.; Kasuga, M.; Dubouzet, E.G.; Miura, S.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and coldresponsive gene expression. Plant J. 2003, 33, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Magome, H.; Yamaguchi, S.; Hanada, A.; Kamiya, Y.; Oda, K. Dwarf and delayedflowering 1, a novel Arabidopsis mutant deficient in gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor. Plant J. 2004, 37, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinazaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an AP2/EREBP DNA-binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Organization of cis-acting regulatory elements in osmotic- and cold-stress responsive promoters. Trends Plant Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef]
- Chen, J.; Xia, X.; Yin, W. Expression profiling and functional characterization of a DREB2-type gene from Populus euphratica. Biochem. Biophys. Res. Commun. 2009, 378, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Galovic, V.; Orlovic, S.; Fladung, M. Characterization of two poplar homologs of the GRAS/SCL gene, which encodes a transcription factor putatively associated with salt tolerance. iForest Biogeosci. For. 2015, 8, 780–785. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.; Yang, M.; Wen, H.; Chern, J. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar] [CrossRef]
- Kim, D.O.; Jeong, S.W.; Lee, C.Y. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem. 2003, 81, 321–326. [Google Scholar] [CrossRef]
- Arnao, M.B. Some methodological problems in the determination of antioxidant activity using chromogen radicals: A practical case. Trends Food Sci. Technol. 2000, 11, 419–421. [Google Scholar] [CrossRef]
- Miller, N.; Rice-Evans, C. Factors influencing the antioxidant activity determined by the ABTS radical cation assay. Free Radic. Res. 1997, 26, 195–199. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- TIBCO Software Inc. Statistica (Data Analysis Software System). Version 13. 2017. Available online: https://docs.tibco.com/products/tibco-statistica-13-3-0 (accessed on 2 April 2021).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2(−delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Vuksanovic, V.; Kovacevic, B.; Kebert, M.; Katanic, M.; Pavlovic, L.; Kesic, L.; Orlovic, S. Clone specificity of white poplar (Populus alba L.) acidity tolerance in vitro. Fresenius Environ. Bull. 2019, 11, 8307–8313. [Google Scholar]
- Štajner, D.; Popović, B.M.; Ćalić-Dragosavac, D.; Malenčić, Đ.; Zdravković-Korać, S. Comparative Study on Allium schoenoprasum Cultivated Plant and Allium schoenoprasum Tissue Culture Organs Antioxidant Status. Phyther. Res. 2011, 25, 1618–1622. [Google Scholar] [CrossRef]
- Meloni, D.A.; Martínez, C.A. Glycinebetaine improves salt tolerance in vinal (Prosopis ruscifolia Griesbach) seedlings. Braz. J. Plant Physiol. 2009, 21, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Zuckerkandl, E.; Pauling, L. Evolutionary divergence and convergence in proteins. In Evolving Genes and Proteins; Bryson, V., Vogel, H.J., Eds.; Academic Press: New York, NY, USA, 1965; pp. 97–166. ISBN 9781483227344. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galović, V.; Orlović, O.; Prathiba, M.J.; Pekeč, S.; Vasić, V.; Vasić, S.; Szabados, L. Characterization of abiotic stress responsive RD29B and RD17 genes in different poplar clones. Topola 2020, 206, 13–20. [Google Scholar] [CrossRef]
- Shi, H.Z.; Quintero, F.J.; Pardo, J.M.; Zhu, J.K. The putative plasma membrane Na+/H+ antiporter SOS1 controls long-distance Na+ transport in plants. Plant Cell 2002, 14, 465–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, D.H.; Lee, S.Y.; Bressan, R.A.; Yun, D.J.; Bohnert, H.J. Intracellular consequences of SOS1 deficiency during salt stress. J. Exp. Bot. 2010, 61, 1205–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lata, C.; Prasad, M. Role of DREBs in regulation of abiotic stress responses in plants. J. Exp. Bot. 2011, 62, 4731–4748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fladung, M.; Buschbom, J. Identification of single nucleotide polymorphisms in different Populus species. Trees 2009, 23, 1199–1212. [Google Scholar] [CrossRef]
- Flowers, T.J.; Munns, R.; Colmer, T.D. Sodium chloride toxicity and the cellular basis of salt tolerance in halophytes. Ann. Bot. 2015, 115, 419–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janz, D.; Behnke, K.; Schnitzler, J.P.; Kanawati, B.; Schmitt-Kopplin, P.; Polle, A. Pathway analysis of the transcriptome and metabolome of salt sensitive and tolerant poplar species reveals evolutionary adaption of stress tolerance mechanisms. BMC Plant Biol. 2010, 10, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone Name | Clone Collection No. | Species |
|---|---|---|
| Bora | B229 | P. deltoides |
| In the process | PE19/66 | P. deltoides |
| Pannonia | M1 | P. x euramericana |
| Antonije | 182/81 | P. deltoides |
| Stress-Induced Genes | NCBI Accession Number | Sequences (5’→3’) | Tm (0C) | Product Size (bp) | |
|---|---|---|---|---|---|
| P5CS1 | EEF01373 | PtP5CS1-F | ggcgttctcctgattgttttt | 60.48 | 84 |
| PtP5CS1-R | gagtccattcccacttctgatt | 59.44 | |||
| SOS1 | EEF02008 | PtSOS1-F | ttgattggaaaaactcctgctc | 60.59 | 146 |
| PtSOS1-R | tcctgatggaatgacagcctac | 61.40 | |||
| DREB2 | XM_002315114 | PtDREB2-F | gattgttctcggggagttga | 60.05 | 78 |
| PtDREB2-R | ccacgaaggattttctgattga | 61.33 | |||
| GRAS17 (Scaffold_7) | XM_002310190 | PtGRAS17-F | cttaaaaatccctctctctctctcc | 59.46 | 118 |
| PtGRAS17-R | tctccagccaaccttcttactt | 59.41 | |||
| GRAS16 | XM_002327770 | PtGRAS16-F | actatttctttagacccaacgacgac | 62.25 | 66 |
| (Scaffold_5) | PtGRAS16-R | atcgcctccacaacagcc | 62.10 | ||
| β-Actin | XM_024591321 | PtActin_F | ggatattcagccccttgtctg | 60.90 | 141 |
| PtActin_R | ttctgccccattccaacc | 61.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galović, V.; Kebert, M.; Popović, B.M.; Kovačević, B.; Vasić, V.; Joseph, M.P.; Orlović, S.; Szabados, L. Biochemical and Gene Expression Analyses in Different Poplar Clones: The Selection Tools for Afforestation of Halomorphic Environments. Forests 2021, 12, 636. https://doi.org/10.3390/f12050636
Galović V, Kebert M, Popović BM, Kovačević B, Vasić V, Joseph MP, Orlović S, Szabados L. Biochemical and Gene Expression Analyses in Different Poplar Clones: The Selection Tools for Afforestation of Halomorphic Environments. Forests. 2021; 12(5):636. https://doi.org/10.3390/f12050636
Chicago/Turabian StyleGalović, Vladislava, Marko Kebert, Boris M. Popović, Branislav Kovačević, Verica Vasić, Mary Prathiba Joseph, Saša Orlović, and László Szabados. 2021. "Biochemical and Gene Expression Analyses in Different Poplar Clones: The Selection Tools for Afforestation of Halomorphic Environments" Forests 12, no. 5: 636. https://doi.org/10.3390/f12050636
APA StyleGalović, V., Kebert, M., Popović, B. M., Kovačević, B., Vasić, V., Joseph, M. P., Orlović, S., & Szabados, L. (2021). Biochemical and Gene Expression Analyses in Different Poplar Clones: The Selection Tools for Afforestation of Halomorphic Environments. Forests, 12(5), 636. https://doi.org/10.3390/f12050636