
Change of Potential Distribution Area of a Forest Tree Acer davidii in East Asia under the Context of Climate Oscillations

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sources
2.2. Environmental Factors and Pretreatment
2.3. Models and Methods
2.4. Model Result Evaluation
3. Results
3.1. Evaluation of the Accuracy of the Model
3.2. Importance of Variables and Climate Preference
3.3. Threshold Analysis of Dominant Environmental Variables
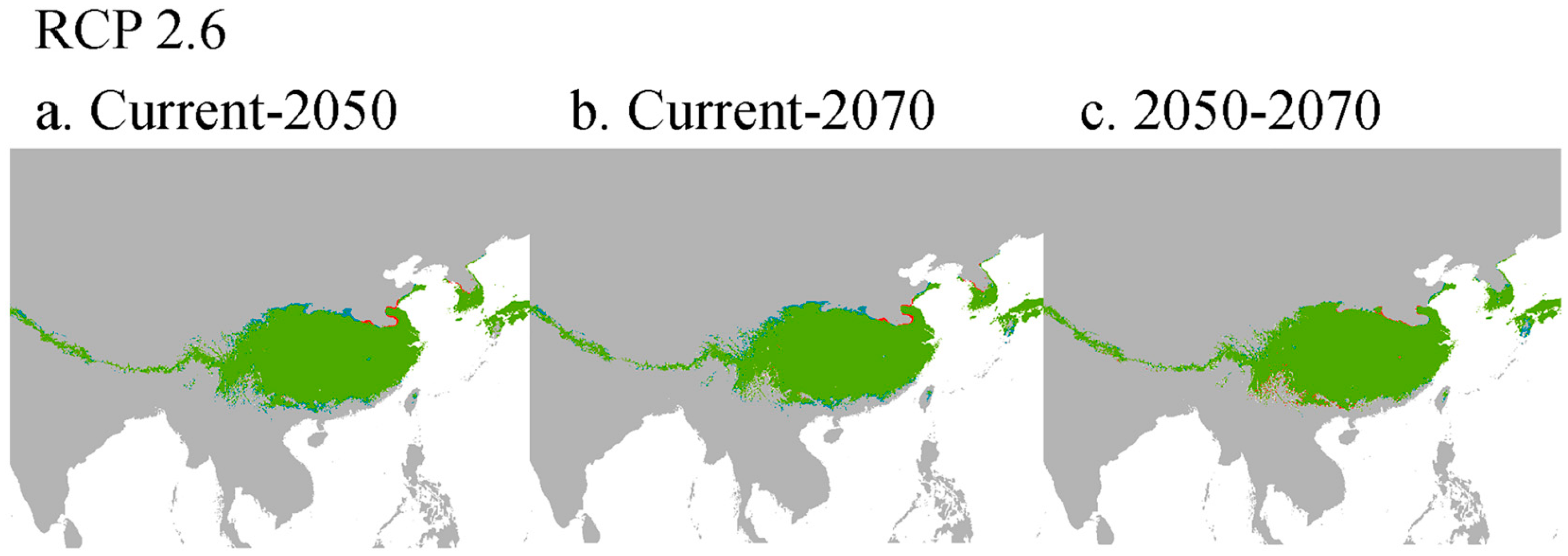
3.4. Changes of Potential Geographical Distribution of A. davidii in Asia under Different Climatic Scenarios
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Cong, M.; Xu, Y.; Tang, L.; Yang, W.; Jian, M. Predicting the dynamic distribution of Sphagnum bogs in China under climate change since the last interglacial period. PLoS ONE 2020, 15. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, M.C.; Gove, A.D.; Sanders, N.J.; Dunn, R.R. Climate change, plant migration, and range collapse in a global biodiversity hotspot: The Banksia (Proteaceae) of Western Australia. Glob. Chang. Biol. 2008, 14, 1337–1352. [Google Scholar] [CrossRef]
- Alberto, F.J.; Aitken, S.N.; Alia, R.; Gonzalez-Martinez, S.C.; Hanninen, H.; Kremer, A.; Lefevre, F.; Lenormand, T.; Yeaman, S.; Whetten, R.; et al. Potential for evolutionary responses to climate change evidence from tree populations. Glob. Chang. Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.-J.; Wang, X.-P.; Zhao, C.-Y.; Yang, X.-M. Diverse responses of vegetation growth to meteorological drought across climate zones and land biomes in northern China from 1981 to 2014. Agric. For. Meteorol. 2018, 262, 1–13. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, A.T.; Papes, M.; Eaton, M. Transferability and model evaluation in ecological niche modeling: A comparison of GARP and Maxent. Ecography 2007, 30, 550–560. [Google Scholar] [CrossRef]
- Li, Y.; Tang, Z.; Yan, Y.; Wang, K.; Cai, L.; He, J.; Gu, S.; Yao, Y. Incorporating species distribution model into the red list assessment and conservation of macrofungi: A case study with Ophiocordyceps sinensis. Biodivers. Sci. 2020, 28, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, G.E. Population studies—Animal ecology and demography—Concluding remarks. Cold Spring Harb. Symp. Quant. Biol. 1957, 22, 415–427. [Google Scholar] [CrossRef]
- Soberon, J. Grinnellian and Eltonian niches and geographic distributions of species. Ecol. Lett. 2007, 10, 1115–1123. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudik, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- David, R.B.; Stockwell, I.R.N. Induction of sets of rules from animal distribution data: A robust and informative method of data analysis. Math. Comput. Simul. 1992, 33, 385–390. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Zhang, Z.-Y.; Chen, L.-D.; Peng, Y.-S.; Wang, X. Changes in potential geographical distribution of Tsoongiodendron odorum since the Last Glacial Maximum. Chin. J. Plant Ecol. 2020, 44, 44–55. [Google Scholar] [CrossRef]
- Garza, G.; Rivera, A.; Venegas Barrera, C.S.; Guadalupe Martinez-Avalos, J.; Dale, J.; Arroyo, T.P.F. Potential Effects of Climate Change on the Geographic Distribution of the Endangered Plant Species Manihot walkerae. Forests 2020, 11, 689. [Google Scholar] [CrossRef]
- Costa, G.C.; Nogueira, C.; Machado, R.B.; Colli, G.R. Sampling bias and the use of ecological niche modeling in conservation planning: A field evaluation in a biodiversity hotspot. Biodivers. Conserv. 2010, 19, 883–899. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Leng, W.; He, H.S.; Bu, R.; Dai, L.; Hu, Y.; Wang, X. Predicting the distributions of suitable habitat for three larch species under climate warming in Northeastern China. For. Ecol. Manag. 2008, 254, 420–428. [Google Scholar] [CrossRef]
- Zhang, K.; Sun, L.; Tao, J. Impact of Climate Change on the Distribution of Euscaphis japonica (Staphyleaceae) Trees. Forests 2020, 11, 525. [Google Scholar] [CrossRef]
- Lin, L.; He, J.; Xie, L.; Cui, G. Prediction of the Suitable Area of the Chinese White Pines (Pinus subsect. Strobus) under Climate Changes and Implications for Their Conservation. Forests 2020, 11, 996. [Google Scholar] [CrossRef]
- Zhao, R.; Chu, X.; He, Q.; Tang, Y.; Song, M.; Zhu, Z. Modeling Current and Future Potential Geographical Distribution of Carpinus tientaiensis, a Critically Endangered Species from China. Forests 2020, 11, 774. [Google Scholar] [CrossRef]
- Velasquez-Tibata, J.; Salaman, P.; Graham, C.H. Effects of climate change on species distribution, community structure, and conservation of birds in protected areas in Colombia. Reg. Environ. Chang. 2013, 13, 235–248. [Google Scholar] [CrossRef]
- Valle, M.; Chust, G.; del Campo, A.; Wisz, M.S.; Olsen, S.M.; Garmendia, J.M.; Borja, A. Projecting future distribution of the seagrass Zostera noltii under global warming and sea level rise. Biol. Conserv. 2014, 170, 74–85. [Google Scholar] [CrossRef]
- Editorial Committee of Chinese Flora. Flora of China; Beijing Science Press: Beijing, China, 1993. [Google Scholar]
- Guo, X.; Luo, Y.J.; Xu, Z.W.; Li, M.Y.; Guo, W.H. Response strategies of Acer davidii to varying light regimes under different water conditions. Flora 2019, 257. [Google Scholar] [CrossRef]
- Zuo, W.; Liu, Q.; Lin, B.; He, H. Physiological responses of 2-year-old Acer davidii seedlings to short-term enhanced UV-B radiation. J. Appl. Ecol. 2005, 16, 1682–1686. [Google Scholar]
- Hanba, Y.T.; Kogami, H.; Terashima, I. The effect of growth irradiance on leaf anatomy and photosynthesis in Acer species differing in light demand. Plant Cell Environ. 2002, 25, 1021–1030. [Google Scholar] [CrossRef]
- Coudun, C.; Gegout, J.-C.; Piedallu, C.; Rameau, J.-C. Soil nutritional factors improve models of plant species distribution: An illustration with Acer campestre (L.) in France. J. Biogeogr. 2006, 33, 1750–1763. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, B.C.; Kim, J.H.; Sim, G.S.; Lee, D.H.; Lee, K.E.; Yun, Y.P.; Pyo, H.B. The isolation and antioxidative effects of vitexin from Acer palmatum. Arch. Pharm. Res. 2005, 28, 195–202. [Google Scholar] [CrossRef]
- Tsuda, M.; Tyree, M.T. Whole-plant hydraulic resistance and vulnerability segmentation in Acer saccharinum. Tree Physiol. 1997, 17, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Royer, D.L.; McElwain, J.C.; Adams, J.M.; Wilf, P. Sensitivity of leaf size and shape to climate within Acer rubrum and Quercus Kelloggii. New Phytol. 2008, 179, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Heikkinen, R.K.; Luoto, M.; Araujo, M.B.; Virkkala, R.; Thuiller, W.; Sykes, M.T. Methods and uncertainties in bioclimatic envelope modelling under climate change. Prog. Phys. Geogr. 2006, 30, 751–777. [Google Scholar] [CrossRef] [Green Version]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Rong, Z.; Zhao, C.; Liu, J.; Gao, Y.; Zang, F.; Guo, Z.; Mao, Y.; Wang, L. Modeling the Effect of Climate Change on the Potential Distribution of Qinghai Spruce (Picea crassifolia Kom.) in Qilian Mountains. Forests 2019, 10, 62. [Google Scholar] [CrossRef] [Green Version]
- Nozawa, T.; Ogura, T.; Yokohata, T.; Okada, N.; Shiogama, H. Climate Change Simulations with a Coupled Ocean-Atmosphere GCM Called the Model for Interdisciplinary Research on Climate: MIROC; National Institute for Environmental Studies: Tsukuba, Japan, 2007. [Google Scholar]
- Wang, J.; Feng, L.; Tang, X.; Bentley, Y.; Hook, M. The implications of fossil fuel supply constraints on climate change projections: A supply-side analysis. Futures 2017, 86, 58–72. [Google Scholar] [CrossRef]
- Rutledge, D. Estimating long-term world coal production with logit and probit transforms. Int. J. Coal Geol. 2011, 85, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Ford, J.D.; Vanderbilt, W.; Berrang-Ford, L. Authorship in IPCC AR5 and its implications for content: Climate change and Indigenous populations in WGII. Clim. Chang. 2012, 113, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Dyderski, M.K.; Paz, S.; Frelich, L.E.; Jagodzinski, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef] [PubMed]
- Puchalka, R.; Dyderski, M.K.; Vitkova, M.; Sadlo, J.; Klisz, M.; Netsvetov, M.; Prokopuk, Y.; Matisons, R.; Mionskowski, M.; Wojda, T.; et al. Black locust (Robinia pseudoacacia L.) range contraction and expansion in Europe under changing climate. Glob. Chang. Biol. 2021, 27, 1587–1600. [Google Scholar] [CrossRef] [PubMed]
- Bush, A.; Mokany, K.; Catullo, R.; Hoffmann, A.; Kellermann, V.; Sgro, C.; McEvey, S.; Ferrier, S. Incorporating evolutionary adaptation in species distribution modelling reduces projected vulnerability to climate change. Ecol. Lett. 2016, 19, 1468–1478. [Google Scholar] [CrossRef]
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef]
- Stewart, J.R.; Lister, A.M.; Barnes, I.; Dalen, L. Refugia revisited: Individualistic responses of species in space and time. Proc. R. Soc. B Biol. Sci. 2010, 277, 661–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jianmeng, F. Spatial patterns of species diversity of seed plants in China and their climatic explanation. Biodivers. Sci. 2008, 16, 470–476. [Google Scholar] [CrossRef]
- Liyan, Z.H.I.; Tianbing, W.U.; Yunying, L.; Lin, Y.U.; Qi, Y.O.U.; Fangjian, C.; Songzhu, H.U. Leaf physiological characteristics of Euscaphis konishii seedlings in drought stress. J. Fujian Coll. For. 2008, 28, 190–192. [Google Scholar]
- Jia, X.; Ma, F.; Zhou, W.; Zhou, L.; Yu, D.; Qin, J.; Dai, L. Impacts of climate change on the potential geographical distribution of broadleaved Korean pine (Pinus koraiensis) forests. Acta Ecol. Sin. 2017, 37, 464–473. [Google Scholar]
- Jian, G.; Hui, Z. Evaluating the spatial distribution of Acer palmatum species based on MaxEnt model. J. Northwest For. Univ. 2021, 36, 163–167. [Google Scholar]
- LI-li, L. Study on the Phenotypic Differences of Acer tawny Population and Prediction of Potential Distribution Areas. Master’s Thesis, Shanxi Normal University, Xi’an, China, 2015. [Google Scholar]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef]
- Jin, J.-X.; Jiang, H.; Peng, W.; Zhang, L.-T.; Lu, X.-H.; Xu, J.-H.; Zhang, X.-Y.; Wang, Y. Evaluating the impact of soil factors on the potential distribution of Phyllostachys edulis (bamboo) in China based on the species distribution model. Chin. J. Plant Ecol. 2013, 37, 631–640. [Google Scholar] [CrossRef]
- Jian-Fu, J.; Fan, X.-C. Simulation of the geographical distribution of three endangered Vitis L. plants in China. Chin. J. Ecol. 2014, 33, 1615–1622. [Google Scholar]
- Bai, Y.; Wei, X.; Li, X. Distributional dynamics of a vulnerable species in response to past and future climate change: A window for conservation prospects. PeerJ 2018, 6. [Google Scholar] [CrossRef]
- Fordham, D.A.; Akcakaya, H.R.; Araujo, M.B.; Elith, J.; Keith, D.A.; Pearson, R.; Auld, T.D.; Mellin, C.; Morgan, J.W.; Regan, T.J.; et al. Plant extinction risk under climate change: Are forecast range shifts alone a good indicator of species vulnerability to global warming? Glob. Chang. Biol. 2012, 18, 1357–1371. [Google Scholar] [CrossRef]
- Vaclavik, T.; Meentemeyer, R.K. Invasive species distribution modeling (iSDM): Are absence data and dispersal constraints needed to predict actual distributions? Ecol. Model. 2009, 220, 3248–3258. [Google Scholar] [CrossRef]
- Qiao, H.; Hu, J.; Huang, J. Theoretical Basis, Future Directions, and Challenges for Ecological Niche Models. Sci. Sin. Vitae 2013, 43, 915–927. [Google Scholar] [CrossRef]
- Hallfors, M.H.; Aikio, S.; Fronzek, S.; Hellmann, J.J.; Ryttari, T.; Heikkinen, R.K. Assessing the need and potential of assisted migration using species distribution models. Biol. Conserv. 2016, 196, 60–68. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Variable | Code |
|---|---|---|
| Climate | SD of temperature change (standard deviation) | bio4 |
| Mean temperature of the coldest quarter/°C | bio11 | |
| Annual precipitation/mm | bio12 | |
| Precipitation of wettest month/mm | bio13 | |
| Precipitation of wettest quarter/mm | bio16 | |
| Precipitation of the warmest quarter/mm | bio18 |
| bio4 | bio11 | bio12 | bio13 | bio16 | bio18 | ||
|---|---|---|---|---|---|---|---|
| Percent contribution | LGM (MIROC) | 10.2 | 40.5 | 10.5 | 0.7 | 0.1 | 38.0 |
| LGM (CCSM) | 10.4 | 41.7 | 11.6 | 0.7 | 0.4 | 35.2 | |
| LIG | 10.9 | 41.6 | 10.1 | 0.7 | 0.2 | 36.5 | |
| Current | 10.2 | 41.9 | 10.8 | 0.5 | 0.3 | 36.3 | |
| 2050 | 8.1 | 76.4 | 0.1 | 13.5 | 1.3 | 0.7 | |
| 2070 | 11.2 | 40.0 | 15.3 | 0.7 | 0.3 | 37.5 | |
| Permutation importance | LGM (MIROC) | 7.7 | 51.3 | 20.8 | 11.2 | 3.8 | 5.1 |
| LGM (CCSM) | 10.1 | 55.8 | 10.7 | 7.9 | 11.6 | 3.8 | |
| LIG | 8.1 | 56.1 | 16.3 | 7.8 | 6.9 | 4.7 | |
| Current | 9.6 | 55.8 | 13.7 | 9.0 | 8.3 | 3.7 | |
| 2050 | 16.1 | 69.1 | 0.2 | 9.9 | 3.4 | 1.2 | |
| 2070 | 8.4 | 53.9 | 15.3 | 8.9 | 7.2 | 6.3 |
| Area of Each Suitable Region (The Percentage Change in Area Compared with Current, %) | ||||
|---|---|---|---|---|
| Period | Marginally Suitable Region | Moderately Suitable Region | Highly Suitable Region | Total Suitable Region |
| LGM (CCSM) | 4.99 (−1.22) | 3.58 (−2.82) | 6.59 (+2.02) | 15.16(−2.03) |
| LGM (MIROC) | 5.24 (−0.97) | 6.18 (−0.22) | 5.09 (+0.52) | 16.51 (−0.68) |
| LIG | 6.64 (+0.43) | 3.48 (−2.92) | 2.52 (−2.05) | 12.64 (−4.55) |
| Current | 6.21 (0.00) | 6.40 (0.00) | 4.57 (0.00) | 17.19 (0.00) |
| 2050 | 22.51 (+16.3) | 37.88 (+31.48) | 9.56 (+4.99) | 69.95 (+52.76) |
| 2070 | 19.19 (+12.98) | 8.64 (+2.24) | 9.32 (+4.75) | 37.16 (+19.97) |
| Area of Each Suitable Region (The Change in Area Compared with Current, km2) | ||||
|---|---|---|---|---|
| Period | Marginally Suitable Region | Moderately Suitable Region | Highly Suitable Region | Unsuitable Region |
| LGM (CCSM) | 111.5191 (−11.3368) | 79.8524 (−46.8716) | 147.1458 (+56.6354) | 1894.2540 (+255.6780) |
| LGM (MIROC) | 117.0781 (−5.7778) | 137.9948 (−11.2708) | 113.6233 (+23.1129) | 1864.0750 (+225.4990) |
| LIG | 129.9705 (+7.1146) | 68.2274 (−58.4966) | 49.3559 (−41.1545) | 1711.1840 (−72.6080) |
| Current | 122.8559 (0.00) | 126.7240 (0.00) | 90.5104 (0.00) | 1638.5760 (0.00) |
| 2050 | 449.9479 (+327.0920) | 757.2726 (+630.5486) | 191.1059 (+100.5955) | 600.7188 (−1037.8572) |
| 2070 | 383.6684 (+260.8125) | 172.7830 (+46.0590) | 186.3767 (+95.8663) | 1256.217 (−382.3590) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Z.; Huang, X.; Zhong, Q.; Liu, M.; Song, X.; Liu, J.; Fu, A.; Tan, J.; Kou, Y.; Li, Z. Change of Potential Distribution Area of a Forest Tree Acer davidii in East Asia under the Context of Climate Oscillations. Forests 2021, 12, 689. https://doi.org/10.3390/f12060689
Su Z, Huang X, Zhong Q, Liu M, Song X, Liu J, Fu A, Tan J, Kou Y, Li Z. Change of Potential Distribution Area of a Forest Tree Acer davidii in East Asia under the Context of Climate Oscillations. Forests. 2021; 12(6):689. https://doi.org/10.3390/f12060689
Chicago/Turabian StyleSu, Zidong, Xiaojuan Huang, Qiuyi Zhong, Mili Liu, Xiaoyu Song, Jianni Liu, Aigen Fu, Jiangli Tan, Yixuan Kou, and Zhonghu Li. 2021. "Change of Potential Distribution Area of a Forest Tree Acer davidii in East Asia under the Context of Climate Oscillations" Forests 12, no. 6: 689. https://doi.org/10.3390/f12060689
APA StyleSu, Z., Huang, X., Zhong, Q., Liu, M., Song, X., Liu, J., Fu, A., Tan, J., Kou, Y., & Li, Z. (2021). Change of Potential Distribution Area of a Forest Tree Acer davidii in East Asia under the Context of Climate Oscillations. Forests, 12(6), 689. https://doi.org/10.3390/f12060689