
Taxonomy, Ecology and Distribution of Juniperus oxycedrus L. Group in the Mediterranean Basin Using Bioclimatic, Phytochemical and Morphometric Approaches, with Special Reference to the Iberian Peninsula


 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
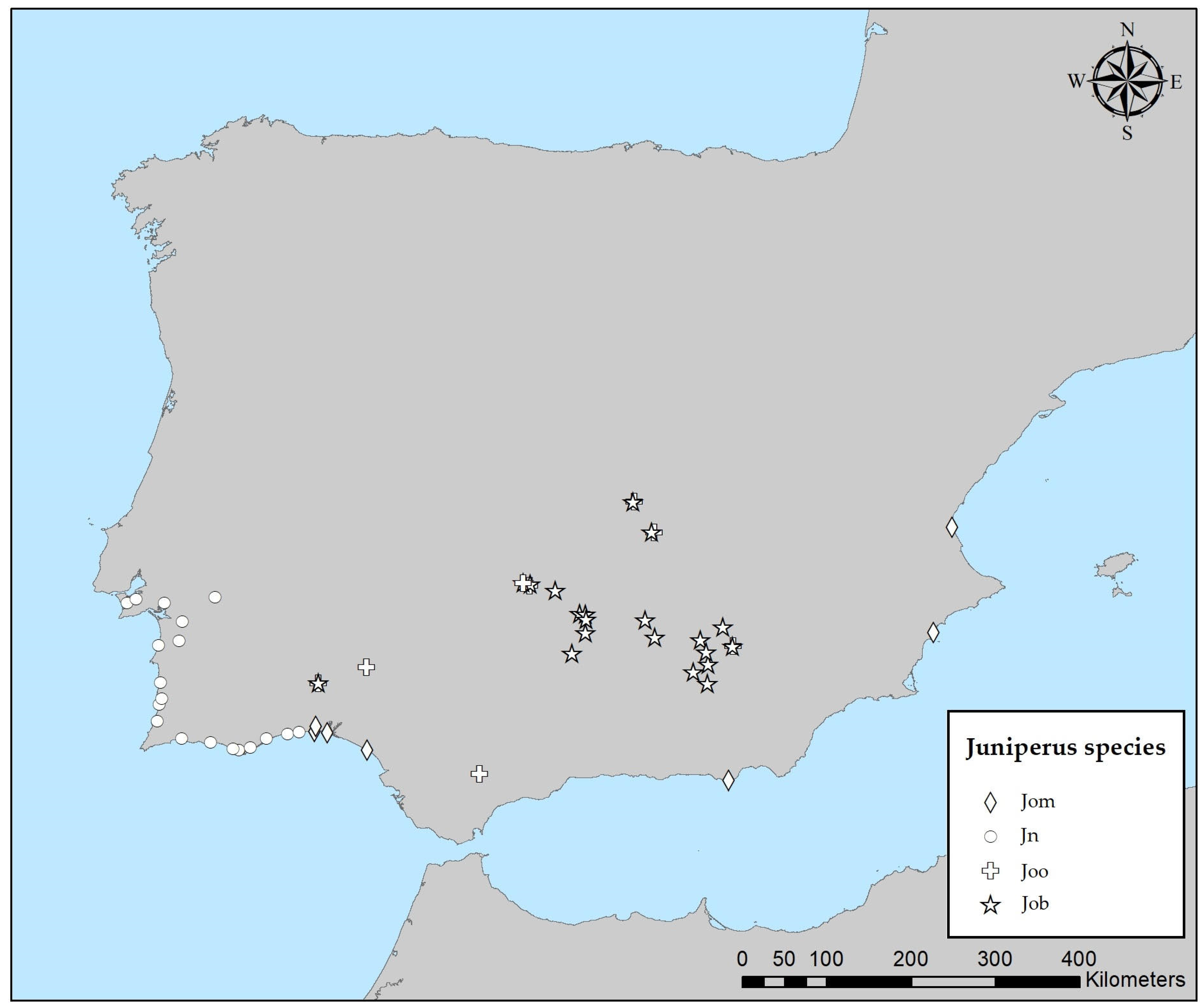
2.1. Sampling Design
2.2. Bioclimatic Analysis
2.3. Phytochemical Analysis
2.4. Morphometric Analysis
- Juniperus oxycedrus taxa studied group;
- Juniperus oxycedrus L. subsp. Oxycedrus;
- J. oxycedrus L. subsp. badia (H. Gay) Debeaux; Syn. J. oxycedrus L. var. badia H. Gay;
- J. oxycedrus L. subsp. macrocarpa (Sm.) Ball; Syn. J. macrocarpa Sm.;
- J. navicularis Gand; Syn. J. oxycedrus L. subsp. transtagana Franco;
3. Results and Discussion
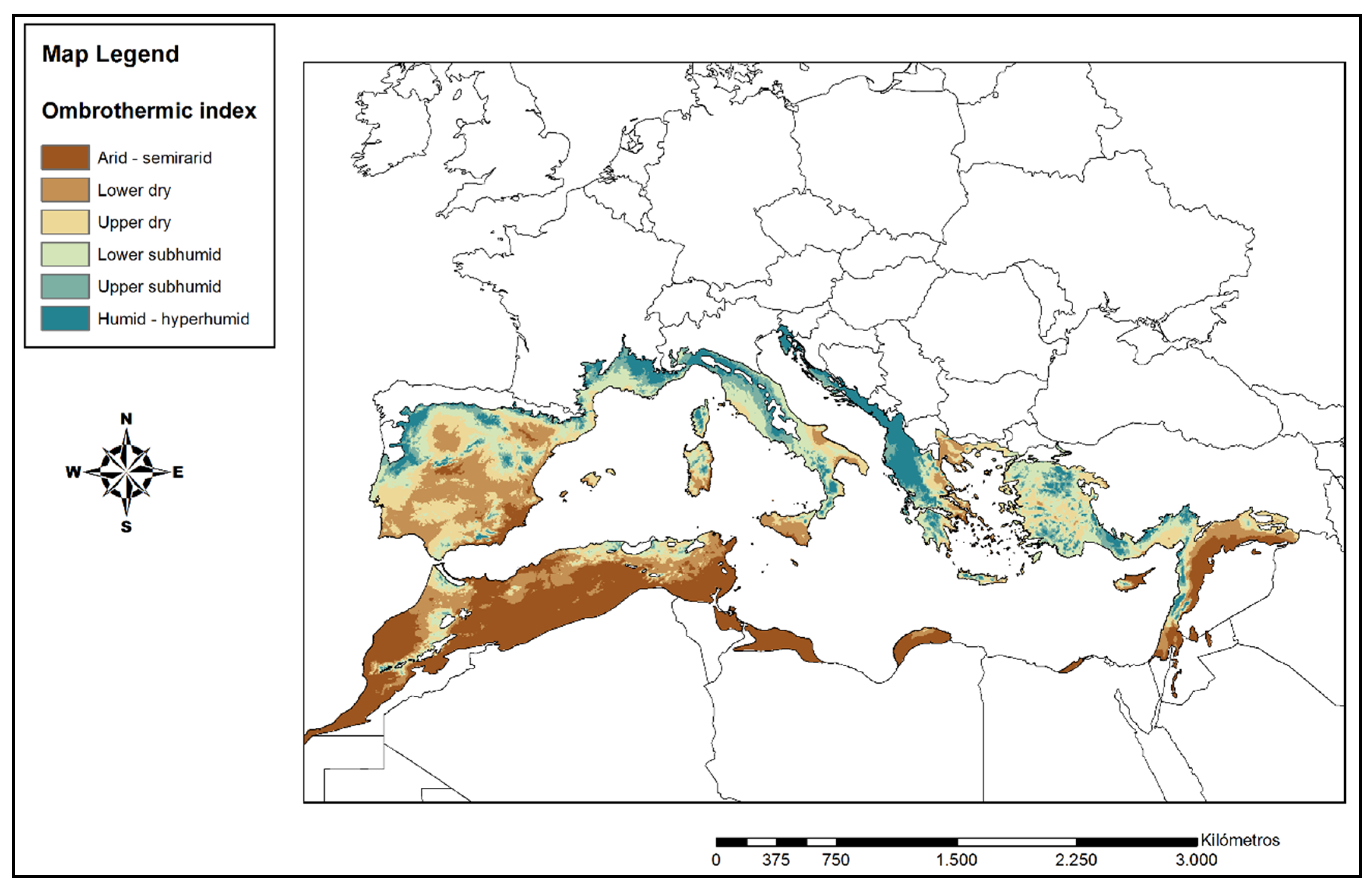
3.1. Bioclimatic Analysis
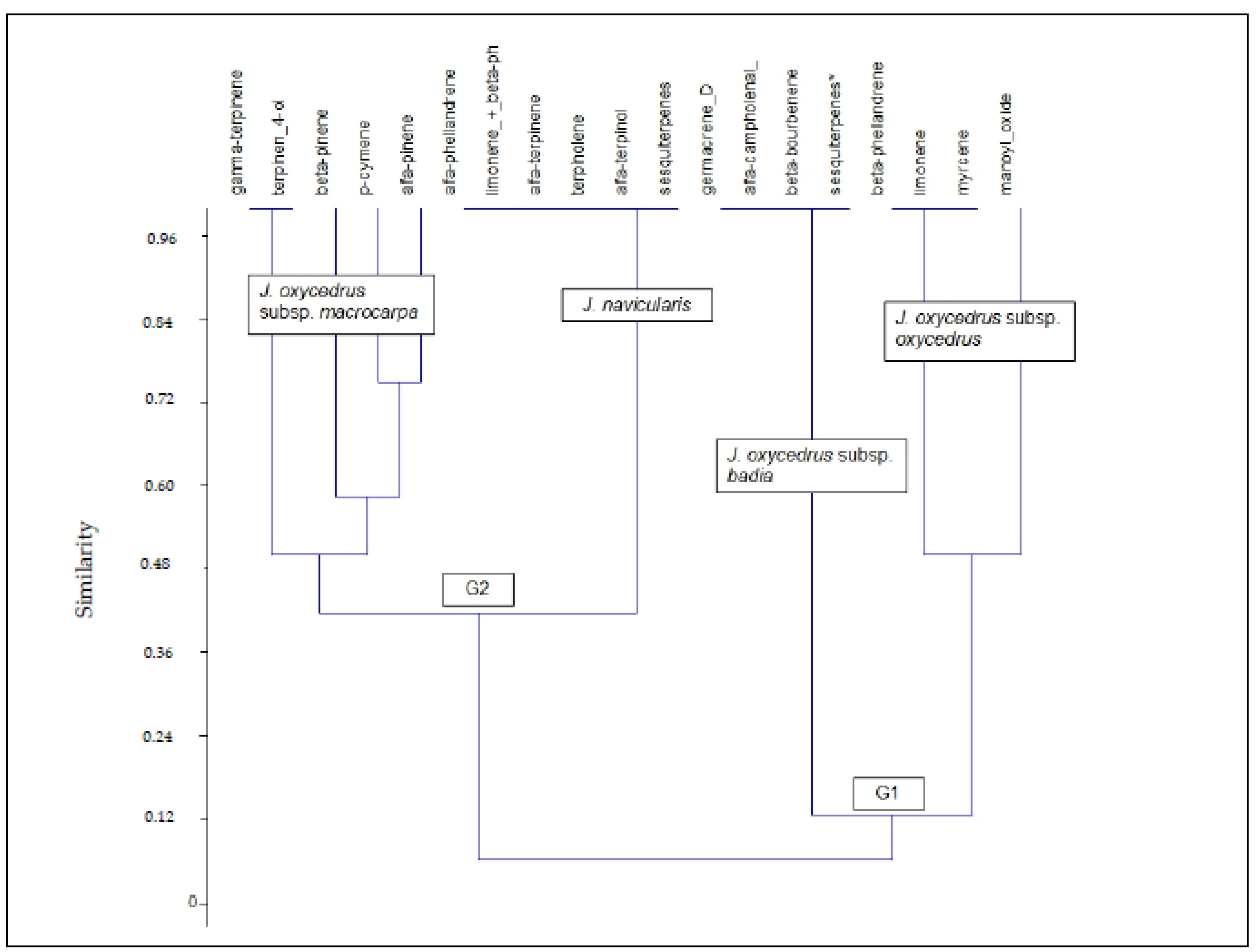
3.2. Phytochemical Analysis
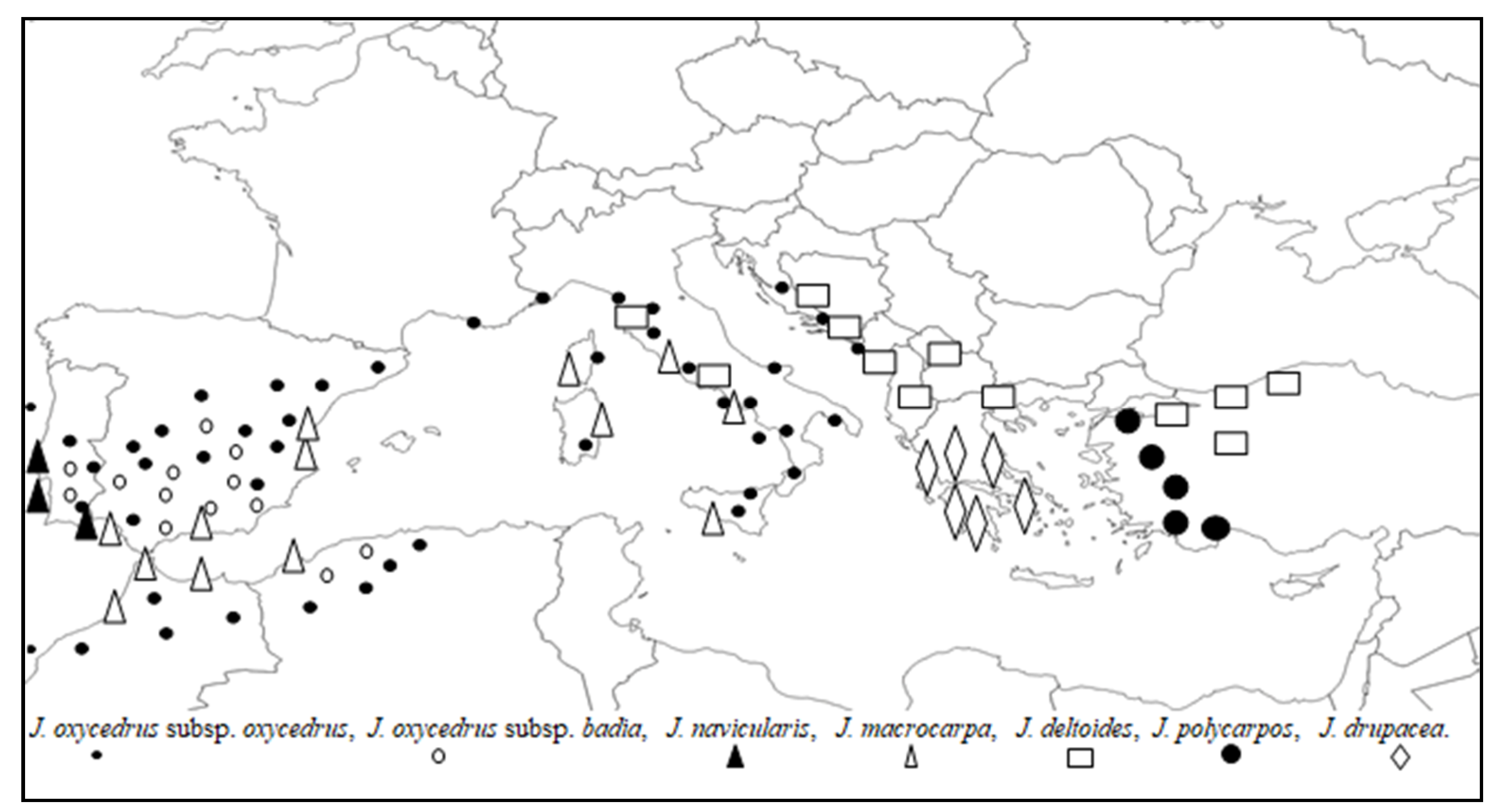
3.3. Morphometric Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vasic, P.S.; Dubak, D.V. Anatomical analysis of red Juniper leaf (Juniperus oxycedrus) taken from Kopaonik Mountain, Serbia. Turk. J. Bot. 2012, 36, 473–479. [Google Scholar]
- Amaral Franco, J. Juniperus. In Flora Ibérica; Real Jardin Botánico: Madrid, Spain, 1986; Volume 1, pp. 181–188. [Google Scholar]
- Burdet, H.M.; Greuter, W.; Long, G. Med-Checklist; Editions des Conservatoire et jardin botaniques de laville de Genève; OPTIMA: Genève, Swizterland, 1984. [Google Scholar]
- Pignatti, S. Juniperus. In Flora d’Italia; Edagricole: Bologna, Italy, 1982; Volume 1, p. 85. [Google Scholar]
- Brus, R.; Idžojtić, M.; Jarni, K. Morphologic variation in northern marginal Juniperus oxycedrus L. subsp. oxycedus population in Istria. Plant Biosyst. 2016, 150, 274–284. [Google Scholar] [CrossRef]
- Diez-Garretas, B.; Pereña, J.; Asensi, A. Morphological analysis, chorology and ecology of Juniperus navicularis Gand. in the Iberian Peninsula. Plant Biosyst. 2017, 151, 168–173. [Google Scholar] [CrossRef]
- Adams, R.P. Juniperus deltoides, a new species, and nomenclatural notes on Juniperus polycarpos and Juniperus turcomanica (Cupressaceae). Phytologia 2004, 86, 49–53. [Google Scholar]
- Adams, R.P. Morphological comparison and key to Juniperus deltoides and J. oxycedrus. Phytologia 2014, 96, 58–62. [Google Scholar]
- Roma-Marzio, F.; Najar, B.; Alessandri, J.; Pistelli, L.; Peruzzi, L. Taxonomy of prickly juniper (Juniperus oxycedrus group): A phytochemical-morphometric combined approach at the contact zone of two cryptospecies. Phytochemistry 2017, 141, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Piñar-Fuentes, J.C.; Cano Ortiz, A.; Musarella, C.M.; Pinto Gomes, C.J.; Spampinato, G.; Cano, E. Rupicolous habitats of interest for conservation in the central-southern Iberian Peninsula. Plant Sociol. 2017, 2, 29–42. [Google Scholar] [CrossRef]
- Perrino, E.V.; Tomaselli, V.; Costa, R.; Pavone, P. Conservation status of habitats (Directive 92/43 EEC) of coastal and low hill belts in a Mediterranean biodiversity hot spot (Gargano–Italy). Plant Biosyst. 2013, 147, 1006–1028. [Google Scholar] [CrossRef]
- Cano, E.; Musarella, C.M.; Cano-Ortiz, A.; Piñar Fuentes, J.C.; Rodríguez Torres, A.; Del Rio González, S.; Pinto Gomes, C.; Quinto-Canas, R.; Spampinato, G. Geobotanical Study of the Microforests of Juniperus oxycedrus subsp. badia in the Central and Southern Iberian Peninsula. Sustainability 2019, 11, 1111. [Google Scholar] [CrossRef] [Green Version]
- European Commission. The Habitats Directive. 2016. Available online: https://ec.europa.eu/environment/nature/legislation/habitatsdirective/index_en.htm (accessed on 26 May 2021).
- Melendo, M.; Giménez, E.; Cano, E.; Gómez Mercado, F.; Valle, F. The endemic flora in the south of the Iberian Peninsula: Taxonomic composition, biological spectrum, pollination, reproductive mode and dispersal. Flora 2003, 198, 260–276. [Google Scholar] [CrossRef] [Green Version]
- Cano, E.; Melendo, M.; Valle, F. The plant communities of the Asplenietea trichomanis in the SW Iberian Península. Folia Geobot. 1997, 32, 361–376. [Google Scholar]
- Cano Ortiz, A.; Piñar, J.C.; Pinto Gomes, C.J.; Musarella, C.M.; Cano, E. Expansion of the Juniperus genus due to anthropic activity. In Old-Growth Forest and Coniferous Forests; Weber, R.P., Ed.; Nova Science Publishers: New York, NY, USA, 2015; pp. 55–65. [Google Scholar]
- Cano, E.; Musarella, C.M.; Cano-Ortiz, A.; Piñar Fuentes, J.C.; Spampinato, G.; Pinto Gomes, C.J. Morphometric analysis and bioclimatic distribution of Glebionis coronaria s.l. (Asteraceae) in the Mediterranean area. PhytoKeys 2017, 81, 103–126. [Google Scholar] [CrossRef]
- Panuccio, M.R.; Fazio, A.; Musarella, C.M.; Mendoza-Fernández, A.J.; Mota, J.F.; Spampinato, G. Seed germination and antioxidant pattern in Lavandula multifida (Lamiaceae): A comparison between core and peripheral populations. Plant Biosyst. 2018, 152, 398–406. [Google Scholar] [CrossRef]
- Musarella, C.M.; Cano-Ortiz, A.; Piñar Fuentes, J.C.; Navas-Ureña, J.; Pinto Gomes, C.J.; Quinto-Canas, R.; Cano, E.; Spampinato, G. Similarity analysis between species of the genus Quercus L. (Fagaceae) in southern Italy based on the fractal dimension. PhytoKeys 2018, 113, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Piñar Fuentes, J.C.; Cano-Ortiz, A.; Musarella, C.M.; Quinto Canas, R.; Pinto Gomes, C.J.; Spampinato, G.; del Río, S.; Cano, E. Bioclimatology, Structure, and Conservation Perspectives of Quercus pyrenaica, Acer opalus subsp. granatensis, and Corylus avellana Deciduous Forests on Mediterranean Bioclimate in the South-Central Part of the Iberian Peninsula. Sustainability 2019, 11, 6500. [Google Scholar] [CrossRef] [Green Version]
- Spampinato, G.; Sciandrello, S.; del Galdo, G.; Puglisi, M.; Tomaselli, V.; Cannavò, S.; Musarella, C.M. Contribution to the knowledge of Mediterranean wetland biodiversity: Plant communities of the Aquila Lake (Calabria, Southern Italy). Plant Sociol. 2019, 56, 53–68. [Google Scholar] [CrossRef]
- Signorino, G.; Cannavò, S.; Crisafulli, A.; Musarella, C.M.; Spampinato, G. Fagonia cretica L. Inf. Bot. Italy 2011, 43, 397–399. [Google Scholar]
- Spampinato, G.; Musarella, C.M.; Cano-Ortiz, A.; Signorino, G. Habitat, occurrence and conservation status of the Saharo-Macaronesian and Southern-Mediterranean element Fagonia cretica L. (Zygophyllaceae) in Italy. J. Arid. Land 2018, 10, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Cano-Ortiz, A.; Musarella, C.M.; Piñar, J.C.; Spampinato, G.; Veloz, A.; Cano, E. Vegetation of the dry bioclimatic areas in the Dominican Republic. Plant Biosyst. 2015, 149, 451–472. [Google Scholar] [CrossRef]
- Vila-Viçosa, C.; Vázquez, F.M.; Mendes, P.; Del Rio, S.; Musarella, C.; Cano-Ortiz, A.; Meireles, C. Syntaxonomic update on the relict groves of Mirbeck’s oak (Quercus canariensis Willd. and Q. marianica C. Vicioso) in Southern Iberia. Plant Biosyst. 2015, 149, 512–526. [Google Scholar] [CrossRef]
- Mendes, P.; Meireles, C.; Vila-Viçosa, C.; Musarella, C.; Pinto-Gomes, C. Best management practices to face degraded territories occupied by Cistus ladanifer shrublands—Portugal case study. Plant Biosyst. 2015, 149, 494–502. [Google Scholar] [CrossRef] [Green Version]
- Quinto-Canas, R.; Mendes, P.; Cano-Ortiz, A.; Musarella, C.M.; Pinto-Gomes, C. Forest fringe communities of the southwestern Iberian Peninsula. Rev. Chapingo Ser. Cienc. For. Y Ambient. 2018, 24, 415–434. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Costa, M.; Castroviejo, S.; Valdés-Bermejo, E. La vegetación de Doñana. Lazaroa 1980, 2, 5–189. [Google Scholar]
- Rivas-Martínez, S.; Lousa, M.; Díaz, T.E.; Fernández-González, F.; Loidi, J. La vegetación del sur de Portugal (Sado, Alentejo y Algarve). Itiner. Geobot. 1990, 3, 5–126. [Google Scholar]
- Cano, E.; Rodríguez-Torres, A.; Gomes, C.P.; García-Fuentest, A.; Torres, J.A.; Salazar, C.; Ruiz-Valenzuela, L.; Cano-Ortiz, A.; Montilla, R.J. Analysis of the Juniperus oxycedrus L. in the centre and south of the Iberian Peninsula (Spain and Portugal). Acta Bot. Gallica 2007, 154, 79–99. [Google Scholar] [CrossRef]
- Adams, R.P.; Joaquin Altarejos, J.; Fernandez, C.; Camacho, A. The leaf essential oils and taxonomy of Juniperus oxycedrus L. subsp. oxycedrus, subsp. badia (H. Gay) Debeaux, and subsp. macrocarpa (Sibth. & Sm.) Ball. J. Essent. Oil Res. 1999, 11, 167–172. [Google Scholar] [CrossRef]
- Salido, S.; Altarejos, J.; Nogueras, M.; Sánchez, A.; Pannecouque, C.; Witvrouw, M.; De Clercq, E. Chemical studies of essential oils of Juniperus oxycedrus ssp. badia. J. Ethnopharmacol. 2002, 81, 129–134. [Google Scholar] [CrossRef]
- Velasco Negueruela, A.; Pérez Alonso, M.J.; Palá-Paúl, J.; Íñigo, A.; López, G. Leaf essential oils analysis of Juniperus navicularis Gandoger. Bot. Complut. 2002, 26, 85–91. [Google Scholar]
- Hayta, S.; Bagci, E. Essencial oil constituets of the leaves, bark and cones of Juniperus oxycedrus subsp. oxycedrus from Turkey. Acta Bot. Gallica 2014, 161, 201–207. [Google Scholar] [CrossRef]
- Montero Burgos, J.L.; González Rebollar, J.L. Diagramas Bioclimáticos; ICONA: Madrid, Spain, 1983. [Google Scholar]
- Rivas-Martínez, S.; Loidi, J. Bioclimatology of the Iberian Peninsula. Itiner. Geobot. 1999, 13, 41–47. [Google Scholar]
- Cano, E.; Cano-Orttiz, A.; Musarella, C.M.; Piñar Fuentes, J.C.; Ighbareyeh, J.M.H.; Leyva, F.; Del Río, S. Mitigating Climate Change through Bioclimatic Applications and Cultivation Techniques in Agriculture (Andalusia, Spain); Jhariya, M.K., Benerjee, A., Meena, R.S., Yadav, D.K., Eds.; Sustainable Agriculture, Forest and Environmental Management; Springer Nature: Singapore, 2019; pp. 31–69. [Google Scholar] [CrossRef]
- Del Río, S.; Cano-Ortiz, A.; Herrero, L.; Penas, A. Recent trends in mean maximum and minimum air temperatures over Spain (1961–2006). Theor. Appl. Climatol. 2012, 109, 605–626. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Penas, Á.; Díaz González, T.E.; Ladero Álvarez, M.; Asensi Marfil, A.; Díez Garretas, B.; Molero Mesa, J.; Valle Tendero, F.; Cano, E.; Costa Talens, M.; et al. Mapa de Series, Geoseries y Geopermaseries de Vegetación de Spain, Parte II. Itiner. Geobot. 2011, 18, 5–424. [Google Scholar]
- Rivas-Martínez, S. Mapa de Series, Geoseries y Geopermaseries de Vegetación de España, Parte I. Itiner. Geobot. 2007, 17, 5–436. [Google Scholar]
- Adams, R.P. Systematics of Juniperus section Juniperus based on leaf essential oils and random amplified polymorphic DNAs (RAPDs). Biochem. Syst. Ecol. 2000, 28, 515–528. [Google Scholar] [CrossRef]
- Anthos. Information System of the Plants of Spain. Real Jardín Botánico, CSIC—Fundación Biodiversidad. Electronic Resource. 2011. Available online: https://www.anthos.es (accessed on 26 May 2021).
- Castroviejo, S.; Aedo, C.; Cirujano, S.; Laínz, M.; Montserrat, P.; Morales, R.; Muñoz Garmendia, F.; Navarro, C.; Paiva, J.; Soriano, C. (Eds.) Flora Iberica 1; Real Jardín Botánico, CSIC: Madrid, Spain, 1986. [Google Scholar]
- Crespo, M.B.; Herbario ABH (Universidad de Alicante). Depto. de Ciencias Ambientales y Recursos Naturales, Universidad de Alicante. Occurrence Dataset. 2018. Available online: https://doi.org/10.15468/hs9hmr (accessed on 2 May 2021).
- Valdés, B.; Talavera, S.; Fernández Galiano, E. Flora Vascular de Andalucía Occidental; Ketres: Barcelona, Spain, 1987; Volume 1–3. [Google Scholar]
- Castroviejo, S. (coord. gen.); Flora Iberica; Real Jardín Botánico, CSIC: Madrid, Spain, 1986–2012. [Google Scholar]
- Tutin, T.; Heywood, V.H.; Burges, D.A.; Valentine, D.H.; Walter, S.M.; Webb, D.A. Flora Europaea; Cambridge University Press: London, UK, 1964–1980; Volumes I–V. [Google Scholar]
- Laguna, M. Flora Forestal Española de los Árboles, Arbustos y Matas. Parte Primera; Editor Máximo Laguna: Madrid, Spain, 1883; pp. 1–374. Available online: https://bdh-rd.bne.es/viewer.vm?id=0000042028&page=1 (accessed on 5 March 2020).
- Laguna, M. Flora Forestal Española de los Árboles, Arbustos y Matas. Parte Segunda; Ministry of Development: Madrid, Spain, 1890; pp. 375–839. Available online: https://bdh-rd.bne.es/viewer.vm?id=0000042028&page=1 (accessed on 5 March 2020).
- Fiori, A. Nuova Flora Analitica d’Italia Contenente la Descrizione Delle Piante Vascolari Indigene, Inselvatichite e Largamente Coltivate in Italia; Tipografia, M., Ed.; Ricci: Firenze, Italy, 1923; Volume 1, p. 279. [Google Scholar]
- Blanca, G.; Cabezudo, B.; Cueto, M.; Fernández López, C.; Morales Torres, C. Flora Vascular de Andalucía Oriental. Selaginellaceae-Ceratophyllaceae; Consejería de Medio Ambiente Junta de Andalucía: Sevilla, Spain, 2009; Volume 1. [Google Scholar]
- Linnaeus, C. Species Plantarum, Exhibentes Plantas Rite Cognitas, ad Genera Relatas, cum Differentiis Specificis, nominibus Trivialibus, Synonymis Selectis, Locis Natalibus, Secundum Systema Sexuale Digestas; Impensis Laurentii Salvii, L. Salvius: Stockholm, Sweden, 1753. [Google Scholar]
- Clusius, C. Rariorum Aliquot Stirpium per Hispanias Observatarum Historia; Antwerpen: Ex officina Christophori Plantini; Biblioteca del Real Jardín Botánico de Madrid: Madrid, Spain, 1576. [Google Scholar]
- Maire, R. Flore de l’Afrique du Nort; Paul Lechevalier: Paris, France, 1952; pp. 1–266. [Google Scholar]
- Bolòs, O.; Vigo, J. Flora dels Països Catalans; Barcino: Barcelona, Spain, 1984. [Google Scholar]
- Vicioso, C. Notas sobre la flora española. Madrid. Anal. Jar. Bot. Madrid 1946, 6, 5–89. [Google Scholar]
- Rivas-Martínez, S.; Díaz, T.E.; Fernández González, F.; Izco, J.; Loidi, J.; Lousa, M.; Penas, A. Vascular plant communities of Spain and Portugal. Itiner. Geobot. 2002, 15, 433–922. [Google Scholar]
- Musarella, C.M.; Cano-Ortiz, A.; Piñar Fuentes, J.C.; del Río, S.; Pinto Gomes, C.J.; Spampinato, G.; Cano, E. Analysis vegetation of Juniperus oxycedrus L. subsp. badia (H. Gay) Debeaux microforests in the Iberian Peninsula: Increasing their knowledge for a better restoration. Flora Medit. 2017, 27, 54–55. [Google Scholar]
- Meireles, C.; Goncalves, P.; Rego, F.; Silveira, S. Estudo da regeneração natural das espécies arbóreas autóctones na Reserva Natural da Serra da Malcata. Silva. Lusit. 2005, 13, 217–231. [Google Scholar]
- Spampinato, G.; Crisarà, R.; Cannavò, S.; Musarella, C.M. Phytotoponims of southern Calabria: A tool for the analysis of the landscape and its transformations. Atti Soc. Tosc. Sci. Nat. Mem. Ser. B 2017, 124, 61–72. [Google Scholar] [CrossRef]
- Boratyński, A.; Wachowiak, W.; Dering, M.; Krystyna, B.; Katarzyna, S.; Sobierajska, K.; Jasińska, A.K.; Klimko, M.; Montserrat, J.M.; Romo, A.; et al. The biogeography and genetic relationships of Juniperus oxycedrus and related taxa from the Mediterranean and Macaronesian regions. Bot. J. Linn. Soc. 2014, 174, 637–653. [Google Scholar] [CrossRef] [Green Version]
- Aggag, S.A. DNA Barcoding Identifies Juniperus oxycedrus subsp. macrocarpa in Derna Region, East Libya. J. Agric. Chem. Biotechnol. 2020, 11, 17–20. [Google Scholar]
- Juan, A.; Fay, M.F.; Pastor, J.; Juan, R.; Fernández, I.; Crespo, M.B. Genetic structure and phylogeography in Juniperus oxycedrus subsp. macrocarpa around the Mediterranean and Atlantic coasts of the Iberian Peninsula, based on AFLP and plastid markers. Eur. J. For. Res. 2012, 131, 845–856. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weather Station. 25–30 Years | Pp (mm) | Tp (°C) | Io | Ombrotype | ETP (mm) | e (mm) | Ioex1 | Ioex2 | Ioex3 | Ombro-Climatic Behaviour of the Locality |
|---|---|---|---|---|---|---|---|---|---|---|
| Almadén-Minas | 625.20 | 194.40 | 3.20 | dry | 808.54 | 161.70 | 0.59 | 1.19 | 1.78 | semiarid |
| Cabezas Rubias | 993.40 | 177.60 | 5.60 | subhumid | 702.14 | 140.42 | 1.20 | 2.40 | 3.60 | dry |
| Aracena (H) | 1025.80 | 175.20 | 5.90 | subhumid | 703.46 | 140.69 | 1.26 | 2.52 | 3.78 | dry |
| Santiago Pontones | 1148.70 | 164.40 | 7.00 | subhumid | 675.23 | 135.04 | 1.54 | 3.08 | 4.62 | dry |
| Vadillo Castril | 1182.20 | 140.40 | 8.40 | humid | 488.88 | 97.72 | 1.93 | 3.86 | 5.79 | subhumid |
| Grazalema | 1962.20 | 183.60 | 11.00 | humid | 726.22 | 145.24 | 2.47 | 4.94 | 7.42 | subhumid |
| Montoro | 522.40 | 210.00 | 2.50 | dry | 903.15 | 180.63 | 0.40 | 0.81 | 1.22 | arid |
| Pozoblanco | 514.40 | 193.20 | 2.70 | dry | 805.45 | 161.09 | 0.45 | 0.91 | 1.37 | arid |
| Villanueva del Arzobispo | 698.20 | 196.80 | 3.50 | dry | 915.70 | 183.14 | 0.65 | 1.30 | 1.96 | semiarid |
| Taxa | ||||
|---|---|---|---|---|
| J. oxycedrus subsp. oxycedrus | J. oxycedrus subsp. badia | J. oxycedrus subsp. macrocarpa | J. navicularis | |
| Essential oils | ||||
| alpha-pinene | 34 | 0.1 | 0.1 | 30.8 |
| limonene | 16.25 | 0 | 0 | 0 |
| beta-pinene | 0.1 | 0 | 0 | 3.3 |
| myrcene | 0.1 | 0 | 0 | 0 |
| p-cymene | 0.1 | 0 | 0.1 | 3 |
| beta-phellandrene | 0.1 | 0 | 0 | 0 |
| manoyl oxide | 0.1 | 0.1 | 0 | 0 |
| germacrene D | 0 | 0.1 | 0 | 0 |
| alpha-campholenal | 0 | 0.1 | 0 | 0 |
| beta-bourbenene | 0 | 0.1 | 0 | 0 |
| Sesquiterpenes/only for 1 species | 0 | 0.1 | 0 | 0 |
| gamma-terpinene | 0 | 0 | 0.1 | 0.5 |
| terpinen 4-ol | 0 | 0 | 0.1 | 0.9 |
| alpha-phellandrene | 0 | 0 | 0 | 11.1 |
| limonene + beta-phellandrene | 0 | 0 | 0 | 27.2 |
| alpha-terpinene | 0 | 0 | 0 | 0.5 |
| terpinolene | 0 | 0 | 0 | 3.4 |
| alpha-terpinol | 0 | 0 | 0 | 1.1 |
| Sesquiterpenes only for 1 species | 0 | 0 | 0 | 0.1 |
| Essential Oils | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 |
| 1 | 0.63 | 0.543 | 0.633 | 0.532 | 0.633 | 0.049 | −0.58 | −0.58 | −0.58 | −0.58 | 0.424 | 0.47 | 0.519 | 0.519 | 0.519 | 0.519 | 0.519 | 0.519 |
| - | 1 | −0.31 | 1 | −0.32 | 1 | 0.577 | −0.33 | −0.33 | −0.33 | −0.33 | −0.42 | −0.38 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 |
| - | - | 1 | −0.31 | 1 | −0.31 | −0.57 | −0.35 | −0.35 | −0.35 | −0.35 | 0.98 | 0.99 | 1 | 1 | 1 | 1 | 1 | 1 |
| - | - | - | 1 | −0.32 | 1 | 0.577 | −0.33 | −0.33 | −0.33 | −0.33 | −0.42 | −0.38 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 |
| - | - | - | - | 1 | −0.32 | −0.59 | −0.36 | −0.36 | −0.36 | −0.36 | 0.98 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| - | - | - | - | - | 1 | 0.577 | −0.33 | −0.33 | −0.33 | −0.33 | −0.42 | −0.38 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 |
| - | - | - | - | - | - | 1 | 0.577 | 0.577 | 0.577 | 0.577 | −0.73 | −0.66 | −0.58 | −0.58 | −0.58 | −0.58 | −0.58 | −0.58 |
| - | - | - | - | - | - | − | 1 | 1 | 1 | 1 | −0.42 | −0.38 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 |
| - | - | - | - | - | - | - | - | 1 | 1 | 1 | −0.42 | −0.38 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 |
| - | - | - | - | - | - | - | - | - | 1 | 1 | −0.42 | −0.38 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 |
| - | - | - | - | - | - | - | - | - | - | 1 | −0.42 | −0.38 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 | −0.33 |
| - | - | - | - | - | - | - | - | - | - | - | 1 | 1 | 0.98 | 0.98 | 0.98 | 0.98 | 0.98 | 0.98 |
| - | - | - | - | - | - | - | - | - | - | - | - | 1 | 0.99 | 0.99 | 0.99 | 0.99 | 0.99 | 0.99 |
| - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 1 | 1 | 1 | 1 | 1 |
| - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 1 | 1 | 1 | 1 |
| - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 1 | 1 | 1 |
| - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 1 | 1 |
| - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 1 |
| A1 | A2 | B1 | B2 | C1 | C2 | D1 | D2 | |
|---|---|---|---|---|---|---|---|---|
| leaf length (mm) | 8–15(25) | 12–18 | 12–20 | 8–20 | 20–25 | 12–20 | 4–12 | 6–10 |
| leaf width (mm) | 1–1.5 | <1.5 | 1.2–2 | >1.5 | 2–2.5 | <3 | 1–1.5 | <1.5 |
| galbuli (mm) | 8–10 | 6–12 | 10–13 | >10 | 12–15 | 12–15 | 7–10 | 5–10 |
| tree habit (m) | scrub < 4 | scrub <6 | tree < 15 | tree 7–12 | scrub < 3 | scrub <5 | scrub < 2 | scrub 1–2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cano Ortiz, A.; Spampinato, G.; Piñar Fuentes, J.C.; Pinto Gomes, C.J.; Quinto-Canas, R.; Cano, E. Taxonomy, Ecology and Distribution of Juniperus oxycedrus L. Group in the Mediterranean Basin Using Bioclimatic, Phytochemical and Morphometric Approaches, with Special Reference to the Iberian Peninsula. Forests 2021, 12, 703. https://doi.org/10.3390/f12060703
Cano Ortiz A, Spampinato G, Piñar Fuentes JC, Pinto Gomes CJ, Quinto-Canas R, Cano E. Taxonomy, Ecology and Distribution of Juniperus oxycedrus L. Group in the Mediterranean Basin Using Bioclimatic, Phytochemical and Morphometric Approaches, with Special Reference to the Iberian Peninsula. Forests. 2021; 12(6):703. https://doi.org/10.3390/f12060703
Chicago/Turabian StyleCano Ortiz, Ana, Giovanni Spampinato, José Carlos Piñar Fuentes, Carlos José Pinto Gomes, Ricardo Quinto-Canas, and Eusebio Cano. 2021. "Taxonomy, Ecology and Distribution of Juniperus oxycedrus L. Group in the Mediterranean Basin Using Bioclimatic, Phytochemical and Morphometric Approaches, with Special Reference to the Iberian Peninsula" Forests 12, no. 6: 703. https://doi.org/10.3390/f12060703
APA StyleCano Ortiz, A., Spampinato, G., Piñar Fuentes, J. C., Pinto Gomes, C. J., Quinto-Canas, R., & Cano, E. (2021). Taxonomy, Ecology and Distribution of Juniperus oxycedrus L. Group in the Mediterranean Basin Using Bioclimatic, Phytochemical and Morphometric Approaches, with Special Reference to the Iberian Peninsula. Forests, 12(6), 703. https://doi.org/10.3390/f12060703