
Landscape-Scale Drivers of Resistance and Resilience to Bark Beetles: A Conceptual Susceptibility Model


Abstract
:1. Introduction
2. Ecology of Bark Beetles
3. History and Ecological Underpinnings of Stand-Level Hazard Models
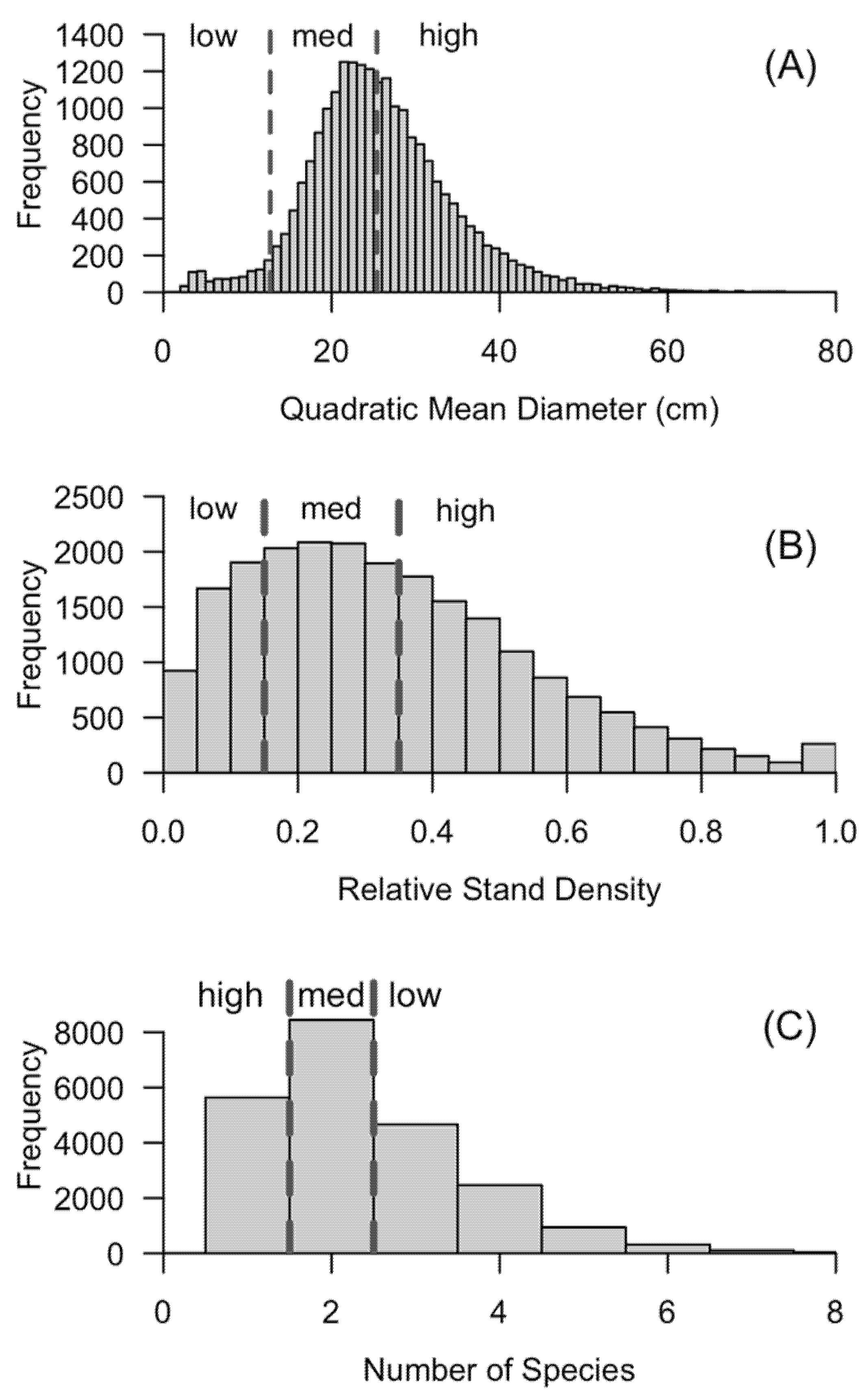
4. Landscape Susceptibility Index: Conceptual Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Susceptibility | Stocking (Relative Stand Density Index) * | Structural Susceptibility (QMD cm) | Tree Species Richness (Count of Species) |
|---|---|---|---|
| High (3) | >35% | >25.4 cm DBH | 1 |
| Medium (2) | 15.1–35% | 12.71–25.4 cm DBH | 2 |
| Low (1) | 0–15% | 2.54–12.7 cm DBH | >3 |
5. Material and Methods
5.1. Testing the Conceptual Model
5.2. FIA Dataset
5.3. Conceptual Model Testing
| Percentage of Plots | Component | Low | Medium | High |
| Stocking (relative stand density index) | 14.9% | 34.8% | 48.6% | |
| Structural Susceptibility (QMD) | 6.7% | 52.3% | 40.9% | |
| Richness (count of tree species) | 38.5% | 39.3% | 22.2% | |
| Scoring of the LSI | ||||
| Stocking (relative stand density index) | (0.149 × 1) = 0.149 | (0.348 × 2) = 0.696 | (0.486 × 3) = 1.458 | |
| Structural Susceptibility (QMD) | (0.067 × 1) = 0.067 | (0.523 × 2) = 1.046 | (0.409 × 3) = 1.227 | |
| Richness (count of tree species) | (0.385 × 1) = 0.385 | (0.393 × 2) = 0.786 | (0.222 × 3) = 0.666 | |
| Total LSI | (0.149 + 0.696 +1.458 + 0.067 + 1.046 + 1.227 +0.385 +0.786 +0.666) = | 6.48 | ||
6. Results
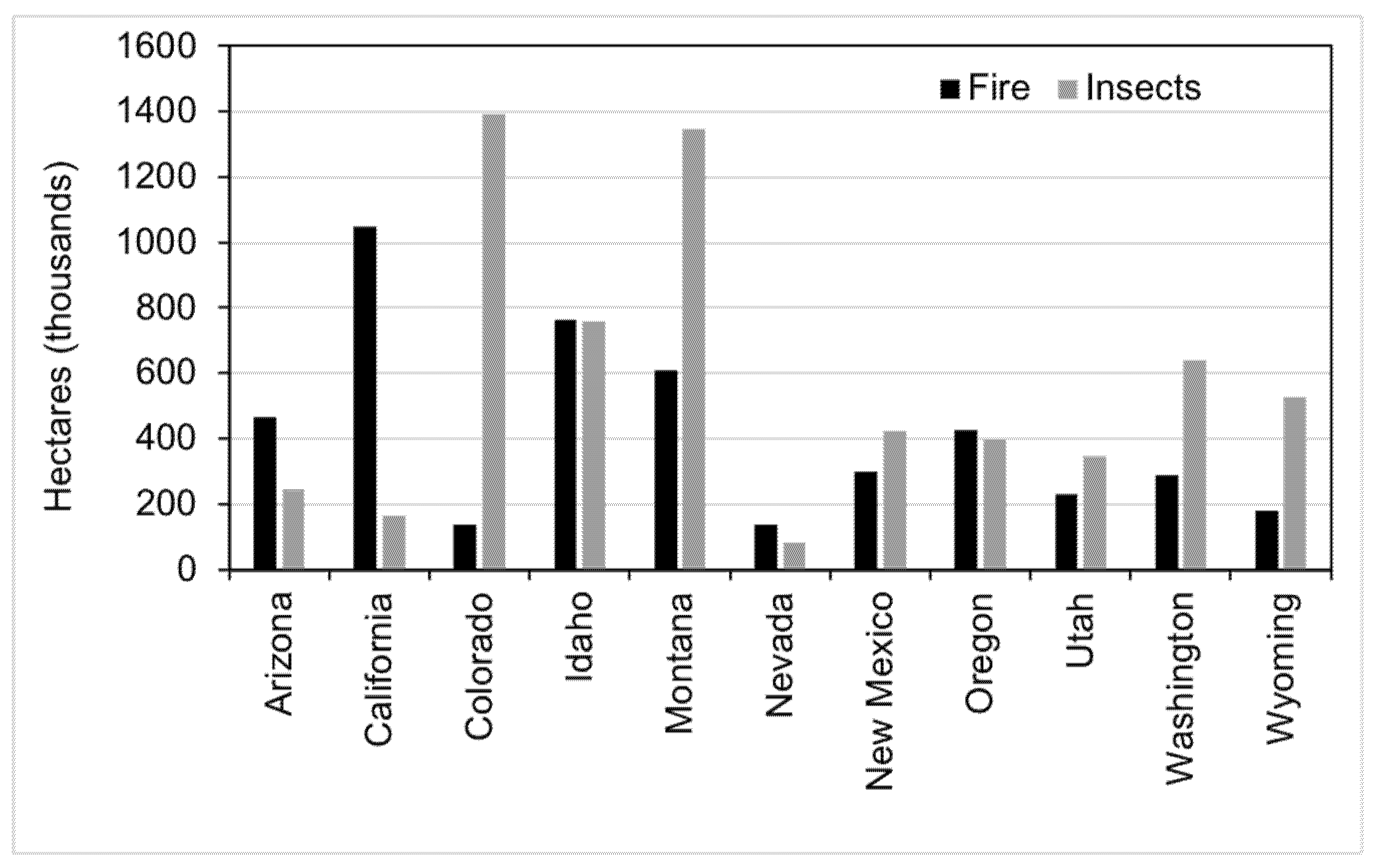
6.1. Interior Western States
6.2. Colorado
6.3. Arizona
| Arizona (n = 2986) LSI = 6.65 | |||
| Component | Low | Medium | High |
| Relative stand density index | 26.8% | 33.6% | 36.9% |
| Quadratic mean diameter | 2.0% | 37.6% | 60.3% |
| Richness (number species) | 31.2% | 35.8% | 33.1% |
| Four Forest Restoration Initiative (4FRI) (n = 1109) LSI = 6.86 | |||
| Component | Low | Medium | High |
| Relative stand density index | 16.8% | 34.9% | 46.5% |
| Quadratic mean diameter | 2.7% | 42.8% | 54.4% |
| Richness (number species) | 40.9% | 34.1% | 25.0% |
| Apache-Sitgreaves National Forest (n = 336) LSI = 6.67 | |||
| Component | Low | Medium | High |
| Relative stand density index | 14.3% | 33.6% | 50.3% |
| Quadratic mean diameter | 3.0% | 44.0% | 52.7% |
| Richness (number species) | 42.6% | 29.5% | 28.0% |
| Coconino National Forest (n = 268) LSI = 6.72 | |||
| Component | Low | Medium | High |
| Relative stand density index | 22.4% | 34.0% | 41.8% |
| Quadratic mean diameter | 3.4% | 38.8% | 57.8% |
| Richness (number species) | 31.7% | 35.1% | 33.2% |
| Kaibab National Forest (n = 267) LSI = 6.79 | |||
| Component | Low | Medium | High |
| Relative stand density index | 13.5% | 37.8% | 47.6% |
| Quadratic mean diameter | 1.9% | 43.4% | 54.7% |
| Richness (number species) | 42.3% | 38.2% | 19.5% |
| Tonto National Forest (n = 238) LSI = 6.42 | |||
| Component | Low | Medium | High |
| Relative stand density index | 17.6% | 34.5% | 45.4% |
| Quadratic mean diameter | 2.5% | 45.0% | 52.5% |
| Richness (number species) | 47.5% | 34.9% | 17.6% |
7. Discussion
8. Implications for Management
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bentz, B.J.; Kegley, S.; Gibson, K.; Thier, R. A test of high-dose verbenone for stand-level protection of lodgepole and whitebark pine from mountain pine beetle (Coleoptera: Curculionidae: Scolytinae) attacks. J. Econ. Entomol. 2005, 98, 1614–1621. [Google Scholar] [CrossRef]
- Brunelle, A.; Rehfeldt, G.E.; Bentz, B.J.; Munson, A.S. Holocene records of Dendroctonus bark beetles in high elevation pine forests of Idaho and Montana, USA. For. Ecol. Manag. 2008, 255, 836–846. [Google Scholar] [CrossRef]
- Morris, J.L.; Derose, R.J.; Brunelle, A.R. Long-term landscape changes in a subalpine spruce-fir forest in central Utah, USA. For. Ecosyst. 2015, 2, 35. [Google Scholar] [CrossRef] [Green Version]
- Thybony, S.; Rosenberg, R.G.; Rosenbergy, E.M. The Medicine Bows: Wyoming’s Mountain Country; Caxton Press: Caldwell, ID, USA, 1985; pp. 1–173. [Google Scholar]
- Keane, R.E.; Ryan, K.C.; Veblen, T.T.; Allen, C.D.; Logan, J.A.; Hawkes, B.; Barron, J. The cascading effects of fire exclu-sion in Rocky Mountain ecosystems. Rocky Mountain futures: An ecological perspective. In Rocky Mountain Futures: An Ecological Perspective; Barron, J., Ed.; Island Press: Washington, DC, USA, 2002; pp. 133–153. [Google Scholar]
- Stephens, S.L.; Burrows, N.; Buyantuyev, A.; Gray, R.W.; Keane, R.E.; Kubian, R.; Liu, S.; Seijo, F.; Shu, L.; Tolhurst, K.G.; et al. Temperate and boreal forest mega-fires: Characteristics and challenges. Front. Ecol. Environ. 2014, 12, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale Drivers of Natural Disturbances Prone to Anthropogenic Amplification: The Dynamics of Bark Beetle Eruptions. Bioscience 2008, 58, 501–517. [Google Scholar] [CrossRef] [Green Version]
- Fettig, C.J.; Hilszczański, J. Management strategies for bark beetles in conifer forests. In Bark Beetles; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 555–584. [Google Scholar]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain pine beetle and forest carbon feedback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Derose, R.J.; Long, J.N. Resistance and Resilience: A Conceptual Framework for Silviculture. For. Sci. 2014, 60, 1205–1212. [Google Scholar] [CrossRef]
- Witt, C.; Shaw, J.D.; Menlove, J.; Goeking, S.A.; DeRose, R.J.; Pelz, K.A.; Morgan, T.A.; Hayes, S.W. Montana’s forest re-sources, 2006–2015. In Resour. Bull. RMRS-RB-30; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2019; p. 30. [Google Scholar]
- Thompson, M.T.; Shaw, J.D.; Witt, C.; Werstak, C.E.; Amacher, M.C.; Goeking, S.A.; DeRose, R.J.; Morgan, T.A.; Sorenson, C.B.; Hayes, S.W.; et al. Colorado’s forest resources, 2004–2013. In Resour. Bull. RMRS-RB-23; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2017. [Google Scholar]
- Woudenberg, S.W.; Conkling, B.L.; O’Connell, B.M.; LaPoint, E.B.; Turner, J.A.; Waddell, K.L. The Forest Inventory and Analysis Database: Database Description and Users Manual Version 4.0 for Phase 2 (No. RMRS-GTR-245); U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2010.
- Hopkins, A.D. The Black Hills Beetle. United States Bureau of Entomology, Series 56. 1905. Available online: https://ir.library.oregonstate.edu/concern/defaults/8623j0070 (accessed on 10 June 2021).
- Chamberlin, W.J. Bark-Beetles Infesting the Douglas fir (Vol. 147); Oregon Agricultural College Experiment Station: Corvallis, OR, USA, 1918. [Google Scholar]
- Aukema, B.H.; McKee, F.R.; Wytrykush, D.L.; Carroll, A.L. Population dynamics and epidemiology of four species of Dendroctonus (Coleoptera: Curculionidae): 100 Years since J.M. Swaine. Can. EÈntomol. 2016, 148, S82–S110. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.C.; Carder, D.R. Sustained Yield of What? J. For. 1977, 75, 722–723. [Google Scholar]
- Keen, F.P. Relative Susceptibility of Ponderosa Pines to Bark-Beetle Attack. J. For. 1936, 34, 919–927. [Google Scholar] [CrossRef]
- Keen, F.P. Ponderosa pine tree classes redefined. J. For. 1943, 41, 249–253. [Google Scholar] [CrossRef]
- Miller, J.M.; Keen, F.P. Biology and Control of the Western Pine Beetle: A Summary of the First Fifty Years of Research (No. 800); US Department of Agriculture: Washington, DC, USA, 1960.
- Steele, R. Stand Hazard Rating for Central Idaho Forests; US Department of Agriculture, Forest Service, Inter-mountain Research Station: Ogden, UT, USA, 1996; Volume 332.
- Daniels, R.F.; Leuschner, W.A.; Zarnoch, S.J.; Burkhart, H.E.; Hicks, R.R. A method for estimating the probability of southern pine beetle outbreaks. For. Sci. 1979, 25, 265–269. [Google Scholar] [CrossRef]
- Hicks, R.R.; Howard, J.E.; Watterston, K.G.; Coster, J.E. Rating forest stand susceptibility to southern pine beetle in East Texas. For. Ecol. Manag. 1979, 2, 269–283. [Google Scholar] [CrossRef]
- Mason, G.N.; Bryant, C.M.V. Establishing southern pine beetle hazard from aerial stand data and historical records. For. Sci. 1984, 30, 375–382. [Google Scholar] [CrossRef]
- Hedden, R.L.; Belanger, R.P. Predicting susceptibility to southern pine beetle attack in the Coastal Plain, Piedmont, and Southern Appalachians. In Integrated Pest Management Research Symposium: The Proceedings; Branhman, S.J., Thatcher, R.C., Eds.; USDA Forest Service, Southern Forest Experiment Station: Ashevhille, NC, USA, 1985; pp. 233–238. [Google Scholar]
- Cook, S.; Cherry, S.; Humes, K.; Guldin, J.; Williams, C. Development of a Satellite-Based Hazard Rating System for Dendroctonus frontallis (Coleoptera: Scolytidae) in the Ouachita Mountains of Arkansas. J. Econ. EÈntomol. 2014, 100, 381–388. [Google Scholar] [CrossRef]
- Eaton, C.B. Insect-Caused Mortality in Relation to Methods of Cutting in Ponderosa Pine on the Blacks Mountain Experi-Mental Forest. Tech. Pap. 43; Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1959; p. 33.
- Sartwell, C.; Stevens, R.E. Mountain pine beetle in ponderosa pine–prospects for silvicultural control in second-growth stands. J. For. 1975, 73, 136–140. [Google Scholar] [CrossRef]
- Amman, G.D.; McGregor, M.D.; Cahill, D.B.; Klein, W.H. Guidelines for Reducing Losses of Lodgepole Pine to the Mountain Pine Beetle in Unmanaged Stands in the Rocky Mountains; General Technical Report, INT-GTR-262; US Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1977; p. 19.
- Stevens, R.E.; McCambridge, W.F.; Edminster, C.B. Risk Rating Guide for Mountain Pine Beetle in Black Hills Ponderosa Pine; Research Note RM-385; US Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1980; p. 2.
- McGregor, M.D.; Amman, G.D.; Cole, W.E. Hazard-rating lodgepole pine for susceptibility to mountain pine beetle in-festation. In Hazard Rating Systems in Forest Insect Pest Management: Symposium Proceedings; Hedden, R.L., Barras, S.J., Coster, J.E., Eds.; Tech. coord. USDA For. Serv. Gen.Tech.Rep.WO-27.; USDA Forest Service: Athens, GA, USA, 1981; pp. 99–104. [Google Scholar]
- Hazard-Rating Lodgepole Pine for Susceptibility to Mountain Pine Beetle Infestation; General Technical Report; USDA Forest Service: Washington, DC, USA, 1981; pp. 99–104.
- Schmid, J.M.; Mata, S.A. Stand Density and Mountain Pine Beetle-Caused Tree Mortality in Ponderosa Pine Stands; USDA, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1992.
- Bentz, B.J.; Amman, G.D.; Logan, J.A. A critical assessment of risk classification systems for the mountain pine beetle. For. Ecol. Manag. 1993, 61, 349–366. [Google Scholar] [CrossRef]
- Stuart, J.D. Hazard rating of lodgepole pine stands to mountain pine beetle outbreaks in southcentral Oregon. Can. J. For. Res. 1984, 14, 666–671. [Google Scholar] [CrossRef]
- Olsen, W.K.; Schmid, J.M.; Mata, S.A. Stand characteristics associated with mountain pine beetle infestations in Ponderosa pine. For. Sci. 1996, 42, 310–327. [Google Scholar] [CrossRef]
- Powell, J.; Kennedy, B.; White, P.; Bentz, B.; Logan, J.; Roberts, D. Mathematical Elements of Attack Risk Analysis for Mountain Pine Beetles. J. Theor. Biol. 2000, 204, 601–620. [Google Scholar] [CrossRef] [Green Version]
- Shore, T.L.; Safranyik, L.; Lemieux, J.P. Susceptibility of lodgepole pine stands to the mountain pine beetle: Testing of a rating system. Can. J. For. Res. 2000, 30, 44–49. [Google Scholar] [CrossRef]
- Parkins, J.R.; MacKendrick, N.A. Assessing community vulnerability: A study of the mountain pine beetle outbreak in British Columbia, Canada. Glob. Environ. Chang. 2007, 17, 460–471. [Google Scholar] [CrossRef]
- Hicke, J.A.; Jenkins, J.C. Mapping lodgepole pine stand structure susceptibility to mountain pine beetle attack across the western United States. For. Ecol. Manag. 2008, 255, 1536–1547. [Google Scholar] [CrossRef]
- Robertson, C.; Wulder, M.A.; Nelson, T.A.; White, J.C. Risk rating for mountain pine beetle infestation of lodgepole pine forests over large areas with ordinal regression modelling. For. Ecol. Manag. 2008, 256, 900–912. [Google Scholar] [CrossRef]
- Negrón, J.F.; Popp, J.B. Probability of ponderosa pine infestation by mountain pine beetle in the Colorado Front Range. For. Ecol. Manag. 2004, 191, 17–27. [Google Scholar] [CrossRef]
- Weatherby, J.C.; Thier, R.W. A preliminary Validation of Douglas-Fir Beetle Hazard Rating System; Mountain Home Ranger District, Boise National Forest; 1993; Forest Pest Management Report No. R4-93-05; U.S. Department of Agriculture, Forest Service: Boise, ID, USA, 1993; p. 7.
- Negron, J.F. Probability of infestation and extent of mortality associated with the Douglas-fir beetle in the Colorado Front Range. For. Ecol. Manag. 1998, 107, 71–85. [Google Scholar] [CrossRef]
- Shore, T.L.; Safranyik, L.; Riel, W.G.; Ferguson, M.; Castonguay, J. Evaluation of factors affecting tree and stand susceptibility to the douglas-fir beetle (coleoptera: Scolytidae). Can. EÈntomol. 1999, 131, 831–839. [Google Scholar] [CrossRef]
- Garrison-Johnston, M.T.; Moore, J.A.; Cook, S.P.; Niehoff, G.J. Douglas-Fir Beetle Infestations Are Associated with Certain Rock and Stand Types in the Inland Northwestern United States. Environ. EÈntomol. 2003, 32, 1354–1363. [Google Scholar] [CrossRef]
- Dodds, K.J.; Ross, D.W.; Randall, C.; Daterman, G.E. Landscape Level Validation of a Douglas-Fir Beetle Stand Hazard-Rating System Using Geographical Information Systems. West. J. Appl. For. 2004, 19, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Dodds, K.J.; Garman, S.L.; Ross, D.W. Risk Rating Systems for the Douglas-Fir Beetle in the Interior Western United States. West. J. Appl. For. 2006, 21, 173–177. [Google Scholar] [CrossRef] [Green Version]
- Schmid, J.M.; Frye, R.H. Stand Ratings for Spruce Beetles; USDA FS RN-RM-309 4p, USDA Forest Service Research Note; Rocky Mountain Forest and Range Experiment Station: Fort Collings, CO, USA, 1976.
- Reynolds, K.; Holsten, E. Relative importance of risk factors for spruce beetle outbreaks. Can. J. For. Res. 1994, 24, 2089–2095. [Google Scholar] [CrossRef]
- Hansen, E.M.; Negron, J.F.; Munson, A.S.; Anhold, J.A. A Retrospective Assessment of Partial Cutting to Reduce Spruce Beetle-Caused Mortality in the Southern Rocky Mountains. West. J. Appl. For. 2006, 25, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Crocker, S.J.; Liknes, G.C.; McKee, F.R.; Albers, J.S.; Aukema, B.H. Stand-level factors associated with resurging mortality from eastern larch beetle (Dendroctonus simplex LeConte). For. Ecol. Manag. 2016, 375, 27–34. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef] [PubMed]
- McKenney, D.W.; Pedlar, J.H.; Lawrence, K.; Campbell, K.; Hutchinson, M.F. Potential Impacts of Climate Change on the Distribution of North American Trees. Bioscience 2007, 57, 939–948. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Crookston, N.L.; Warwell, M.V.; Evans, J. Empirical Analyses of Plant-Climate Relationships for the Western United States. Int. J. Plant Sci. 2006, 167, 1123–1150. [Google Scholar] [CrossRef]
- O’Hara, K.L.; Nagel, L.M. The Stand: Revisiting a Central Concept in Forestry. J. For. 2013, 111, 335–340. [Google Scholar] [CrossRef] [Green Version]
- Royama, T. Analytical Population Dynamics; Chapman & Hall: London, UK, 1992. [Google Scholar]
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate Change and Bark Beetles of the Western United States and Canada: Direct and Indirect Effects. Bioscience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Hansen, E.M.; Bentz, B.J. Comparison of reproductive capacity among univoltine, semivoltine, and re-emerged parent spruce beetles (Coleoptera: Scolytidae). Can. EÈntomol. 2003, 135, 697–712. [Google Scholar] [CrossRef] [Green Version]
- Mitton, J.B.; Ferrenberg, S. Mountain Pine Beetle Develops an Unprecedented Summer Generation in Response to Climate Warming. Am. Nat. 2012, 179, E163–E171. [Google Scholar] [CrossRef] [Green Version]
- Derose, R.J.; Bekker, M.F.; Long, J.N. Traumatic resin ducts as indicators of bark beetle outbreaks. Can. J. For. Res. 2017, 47, 1168–1174. [Google Scholar] [CrossRef]
- Raffa, K.F. Mixed messages across multiple trophic levels: The ecology of bark beetle chemical communication systems. Chemoecology 2001, 11, 49–65. [Google Scholar] [CrossRef]
- Amman, G.D.; McGregor, M.D.; Dolph, R.E., Jr. Mountain Pine Beetle; USDA-Forest Service Forest Insect & Disease Leaflet 2; USDA-Forest Service: Portland, OR, USA, 1990.
- Six, D.L.; Bracewell, R. Dendroctonus. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: London, UK, 2015; pp. 305–350. [Google Scholar]
- Safranyik, L.; Shore, T.L.; Moeck, H.A.; Whitney, H.S. Dendroctonus Ponderosae Hopkins, Mountain Pine Beetle (Coleoptera: Scolytidae); CABI Publishing: Wallingford, UK, 2001; pp. 104–109. [Google Scholar]
- Amman, G.D. Prey Consumption and Development of Thanasimus undatulus, 1 a Predator of the Mountain Pine Beetle 2. Environ. EÈntomol. 1972, 1, 528–530. [Google Scholar] [CrossRef]
- Poland, T.M.; Borden, J.H. Attraction of a bark beetle predator, Thanasimus undatulus (Coleoptera: Cleridae), to pheromones of the spruce beetle and two secondary bark beetles (Coleoptera: Scolytidae). J. Entomol. Soc. Br. Columbia 1997, 94, 35–42. [Google Scholar]
- Ross, D.W.; Daterman, G.E. Using pheromone-baited traps to control the amount and distribution of tree mortality during outbreaks of the Douglas-fir beetle. For. Sci. 1997, 43, 65–70. [Google Scholar]
- Weed, A.S.; Bentz, B.J.; Ayres, M.P.; Holmes, T.P. Geographically variable response of Dendroctonus ponderosae to winter warming in the western United States. Landsc. Ecol. 2015, 30, 1075–1093. [Google Scholar] [CrossRef]
- Fettig, C.J.; Klepzig, K.D.; Billings, R.F.; Munson, A.S.; Nebeker, T.E.; Negrón, J.F.; Nowak, J.T. The effectiveness of vegetation management practices for prevention and control of bark beetle infestations in coniferous forests of the western and southern United States. For. Ecol. Manag. 2007, 238, 24–53. [Google Scholar] [CrossRef]
- Jenkins, M.J.; Hebertson, E.G.; Munson, A.S. Spruce Beetle Biology, Ecology and Management in the Rocky Mountains: An Addendum to Spruce Beetle in the Rockies. Forest 2014, 5, 21–71. [Google Scholar] [CrossRef] [Green Version]
- Randall, C.; Steed, B.; Bush, R. Revised R1 Forest Insect Hazard Rating System User Guide for Use with Inventory Data Stored in FSVeg and/or Analyzed with the Forest Vegetation Simulation. USDA Forest Health Protection Report 11-06. 2011. Available online: https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5346736.pdf (accessed on 10 June 2021).
- Reed, D.D.; Burkhart, H.E.; Leuschner, W.A.; Hedden, R.L. A severity model for southern pine beetle infestations. For. Sci. 1981, 27, 290–296. [Google Scholar] [CrossRef]
- Burns, R.M.; Honkala, B.H. (Eds.) Silvics of North America: Volume 1. Conifers, Agric. Handb. 654; USDA/FS: Washington, DC, USA, 1990.
- Conner, L.G.; Bunnell, M.C.; Gill, R.A. Forest diversity as a factor influencing Engelmann spruce resistance to beetle outbreaks. Can. J. For. Res. 2014, 44, 1369–1375. [Google Scholar] [CrossRef]
- Mitchell, R.G.; Waring, R.H.; Pitman, G.B. Thinning lodgepole pine increases tree vigor and resistance to mountain pine beetle. For. Sci. 1983, 29, 204–211. [Google Scholar] [CrossRef]
- Long, N.J.; Windmuller-Campione, M.A.; DeRose, J.R. Building Resistance and Resilience: Regeneration Should Not be Left to Chance. Forests 2018, 9, 270. [Google Scholar] [CrossRef] [Green Version]
- Bentz, B.J.; Jönsson, A.M. Modeling Bark Beetle Responses to Climate Change. In Bark Beetles; Elsevier BV: Amsterdam, The Netherlands, 2015; pp. 533–553. [Google Scholar]
- Shaw, J.D.; Menlove, J.; Witt, C.; Morgan, T.A.; Amacher, M.C.; Goeking, S.A.; Werstak, C.E. Arizona’s Forest Resources, 2001–2014. Resour. Bull. RMRS-RB-25; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2018; p. 126.
- Audley, J.P.; Fettig, C.J.; Munson, A.S.; Runyon, J.B.; Mortenson, L.A.; Steed, B.E.; Gibson, K.E.; Jørgensen, C.L.; McKelvey, S.R.; McMillin, J.D.; et al. Impacts of mountain pine beetle outbreaks on lodgepole pine forests in the Inter-mountain West, US, 2004–2019. For. Ecol. Manag. 2020, 475, 118403. [Google Scholar] [CrossRef]
- Wurtzebach, Z.; DeRose, R.J.; Bush, R.R.; Goeking, S.A.; Healey, S.; Menlove, J.; Pelz, K.A.; Schultz, C.; Shaw, J.D.; Witt, C. Supporting National Forest System Planning with Forest Inventory and Analysis Data. J. For. 2020, 118, 289–306. [Google Scholar] [CrossRef]
- Underhill, J.L.; Dickinson, Y.; Rudney, A.; Thinnes, J. Silviculture of the Colorado Front Range Landscape Restoration Initiative. J. For. 2014, 112, 484–493. [Google Scholar] [CrossRef]
- Dickinson, Y. Landscape restoration of a forest with a historically mixed-severity fire regime: What was the historical landscape pattern of forest and openings? For. Ecol. Manag. 2014, 331, 264–271. [Google Scholar] [CrossRef]
- Shaw, J.D. Application of Stand Density Index to Irregularly Structured Stands. West. J. Appl. For. 2000, 15, 40–42. [Google Scholar] [CrossRef] [Green Version]
- Long, J.N. A Practical Approach to Density Management. For. Chron. 1985, 61, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urgenson, L.S.; Ryan, C.M.; Halpern, C.B.; Bakker, J.D.; Belote, R.T.; Franklin, J.F.; Haugo, R.D.; Nelson, C.R.; Waltz, A.E. Visions of Restoration in Fire-Adapted Forest Landscapes: Lessons from the Collaborative Forest Landscape Restoration Program. Environ. Manag. 2017, 59, 338–353. [Google Scholar] [CrossRef]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Kolb, T.E. Responses of tree-killing bark beetles to a changing climate. In Climate Change and Insect Pests; Björkman, C., Niemelä, P., Eds.; CAB International: Oxfordshire, UK, 2015; pp. 173–201. [Google Scholar]
- Arno, S.F.; Harrington, M.G.; Fiedler, C.E.; Carlson, C.E. Restoring fire-dependent ponderosa pine forests in western Montana. Restor. Manag. Notes 1995, 13, 32–36. Available online: https://www.jstor.org/stable/43440410 (accessed on 10 June 2021).
- Weidman, R.H. Forest succession as a basis of the silviculture of western yellow pine. J. For. 1921, 19, 877–885. [Google Scholar] [CrossRef]
- Henry, A.; Flood, M.G. The Douglas firs: A botanical and silvicultural description of the various species of Pseudotsuga. In Proceedings of the Royal Irish Academy. Section B: Biological, Geological, and Chemical Science; Royal Irish Academy: Dublin, Ireland, 1919; Volume 35, pp. 67–92. [Google Scholar]
- Peet, R.K. Forest vegetation of the Colorado Front Range: Patterns of species diversity. Vegetatio 1978, 37, 65–78. [Google Scholar] [CrossRef]
- Pedlar, J.H.; McKenney, D.W.; Aubin, I.; Beardmore, T.; Beaulieu, J.; Iverson, L.R.; O’Neill, G.A.; Winder, R.S.; Ste-Marie, C. Placing Forestry in the Assisted Migration Debate. Bioscience 2012, 62, 835–842. [Google Scholar] [CrossRef]
- Taylor, S.W.; Carroll, A.L.; Alfaro, R.I.; Safranyik, L. Forest, climate and mountain pine beetle outbreak dynamics in western Canada. In The Mountain Pine Beetle: A Synthesis of Biology, Management, and Impacts on Lodgepole Pine; Safranyik, L., Wilson, W.R., Eds.; Canadian Forest Service: Victoria, BC, Canada, 2006; pp. 67–94. [Google Scholar]
- Mikkelson, K.M.; Bearup, L.A.; Maxwell, R.M.; Stednick, J.D.; McCray, J.E.; Sharp, J.O. Bark beetle infestation impacts on nutrient cycling, water quality and interdependent hydrological effects. Biogeochemistry 2013, 115, 1–21. [Google Scholar] [CrossRef]
- Chen, H.Y.H.; Luo, Y.; Reich, P.B.; Searle, E.B.; Biswas, S.R. Climate change-associated trends in net biomass change are age dependent in western boreal forests of Canada. Ecol. Lett. 2016, 19, 1150–1158. [Google Scholar] [CrossRef]
- Kane, J.M.; Varner, J.M.; Metz, M.; van Mantgem, P.J. Characterizing interactions between fire and other disturbances and their impacts on tree mortality in western U.S. Forests. For. Ecol. Manag. 2017, 405, 188–199. [Google Scholar] [CrossRef]
- Bryant, T.; Waring, K.; Meador, A.S.; Bradford, J.B. A Framework for Quantifying Resilience to Forest Disturbance. Front. For. Glob. Chang. 2019, 2, 56. [Google Scholar] [CrossRef]
- Wermelinger, B. Ecology and management of the spruce bark beetle Ips typographus—A review of recent research. For. Ecol. Manag. 2004, 202, 67–82. [Google Scholar] [CrossRef]
- Seidl, R.; Müller, J.; Hothorn, T.; Bässler, C.; Heurich, M.; Kautz, M. Small beetle, large-scale drivers: How regional and landscape factors affect outbreaks of the European spruce bark beetle. J. Appl. Ecol. 2016, 53, 530–540. [Google Scholar] [CrossRef]
- Nyland, R.D. Silviculture: Concepts and Applications; Waveland Press: Long Grove, IL, USA, 2016. [Google Scholar]
- Morris, J.L.; Cottrell, S.; Fettig, C.J.; Derose, R.J.; Mattor, K.M.; Carter, V.A.; Clear, J.; Clement, J.; Hansen, W.; Hicke, J.A.; et al. Bark beetles as agents of change in social-ecological systems. Front. Ecol. Environ. 2018, 16, S34–S43. [Google Scholar] [CrossRef]
- Johnson, E.A.; Miyanishi, K.; Weir, J.M.H. Old-growth, disturbance, and ecosystem management. Can. J. Bot. 1995, 73, 918–926. [Google Scholar] [CrossRef]
- Lundquist, J.E.; Reich, R.M. Landscape Dynamics of Mountain Pine Beetles. For. Sci. 2013, 60, 464–475. [Google Scholar] [CrossRef]
- Windmuller-Campione, M.A.; Long, J.N. If Long-Term Resistance to a Spruce Beetle Epidemic is Futile, Can Silvicultural Treatments Increase Resilience in Spruce-Fir Forests in the Central Rocky Mountains? Forests 2015, 6, 1157–1178. [Google Scholar] [CrossRef]




| Species | Common Name | Hosts | Susceptibility, Risk, and Hazard Ratings |
|---|---|---|---|
| Dendroctonus brevicomis Le Conte | western pine beetle | Pinus ponderosae Douglas ex Lawson | [18,19,20,21] |
| Dendroctonus frontalis Zimmermann | southern pine beetle | P. echinata Mill, P. elliottii Engelm., P. engelmannii Carr., P. leiophylla Schiede ex Schlechtendal et Chamisso, P. palustris Miller, P. ponderosae, P. taeda Linnaeus, P. virginiana Miller, others | [22,23,24,25,26] |
| Dendroctonus ponderosae Hopkins | mountain pine beetle | Pinus albicaulis Engelm., P. aristata Engelm., P. contorta Douglas, P. flexilis E. James, P. lambertiana Douglas, P. monticola Douglas ex D. Don in Lambert, P. ponderosae, others | [27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42] |
| Dendroctonus pseudotsugae Hopkins | Douglas-fir beetle | Pseudotsuga menziesii (Mirbel) Franco | [43,44,45,46,47,48] |
| Dendroctonus rufipennis Kirby | spruce beetle | Picea engelmannii Parry ex Engelmann, Pi. glauca (Moench) Voss, Pi. sitchensis (Bongard) Carrière | [49,50,51] |
| Dendroctonus simplex LeConte | eastern larch beetle | Larix laricina (DuRoi) K. Koch | [52] |
| Susceptibility Rating System | Host Size | Relative Density | Relative Density of Host | Other Variables |
|---|---|---|---|---|
| D. brevicomis in Pinus ponderosae [31] | Average diameter of P. ponderosae | Stand basal area | Proportion of stand basal area in P. ponderosae | Stand structure |
| Western pine beetle in D. ponderosae [31] | Average diameter of Ponderosa pine | Stand basal area | Proportion of stand basal area in Ponderosa pine | Stand structure |
| D. frontalis in southern pine species [34] | Average diameter of the stand | Total basal area of the stand | Not used—generally in single-species plantations | Number of attacked trees and number of other attack spots |
| D. ponderosae combined host model [72] | Average diameter of Pinus spp. Species | Stand basal area | Proportion of the stand basal area in Pinus spp. | |
| D. ponderosae in P. contorta [48] | Not used | Stand density index | Proportion of the stand basal area in P. contorta | Average stand age and elevation |
| Southern pine beetle [73] | Average diameter of the stand | Total basal area of the stand | Not used+ | Number of attacked trees and number of other attack spots |
| D. pseudotsugae in Ps. menziesii [52] | Average DBH of Ps. menziesii | Stand basal area | Proportion of stand basal area in Ps. menziesii | Average stand age |
| D. rufipennis in Pi. engelmannii [58] | Average diameter of live Pi. engelmannii | Stand basal area | Proportion of the stand basal area in Pi. engelmannii | Physiographic location |
| D. simplex for L. laricina [61] | Average diameter of L. laricina | Density of non-host gymnosperms | Not used | Presence of water and site index |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Windmuller-Campione, M.A.; DeRose, J.; Long, J.N. Landscape-Scale Drivers of Resistance and Resilience to Bark Beetles: A Conceptual Susceptibility Model. Forests 2021, 12, 798. https://doi.org/10.3390/f12060798
Windmuller-Campione MA, DeRose J, Long JN. Landscape-Scale Drivers of Resistance and Resilience to Bark Beetles: A Conceptual Susceptibility Model. Forests. 2021; 12(6):798. https://doi.org/10.3390/f12060798
Chicago/Turabian StyleWindmuller-Campione, Marcella A., Justin DeRose, and James N. Long. 2021. "Landscape-Scale Drivers of Resistance and Resilience to Bark Beetles: A Conceptual Susceptibility Model" Forests 12, no. 6: 798. https://doi.org/10.3390/f12060798
APA StyleWindmuller-Campione, M. A., DeRose, J., & Long, J. N. (2021). Landscape-Scale Drivers of Resistance and Resilience to Bark Beetles: A Conceptual Susceptibility Model. Forests, 12(6), 798. https://doi.org/10.3390/f12060798